
不同土地利用方式对土壤磷有效性和溶磷细菌群落结构的影响
2021-01-13 05:05王敦刚张俏燕曹文超周广帆王进闯
热带作物学报 2021年12期
关键词:土地利用
王敦刚 张俏燕 曹文超 周广帆 王进闯


摘 要:為探究不同土地利用方式对土壤磷有效性和溶磷细菌群落的影响,以海南省白沙县龙眼园(LO)和由龙眼园转化的有机茶园(OTP)、传统无机茶园(CTP)土壤为研究对象,利用Illumina高通量测序技术分析土壤phoD功能微生物的群落结构,研究土壤速效磷与phoD功能微生物和酸性磷酸酶的相关性。结果表明:龙眼园转换成无机茶园后,在0~10 cm土层,pH、总氮(TN)、总磷(TP)、速效钾(AK)和有机碳(SOC)含量显著降低。同时,在10~20 cm土层,pH、总氮(TN)、总磷(TP)和有机碳(SOC)含量也显著降低。在0~10 cm土层,土壤速效磷(AP)含量在龙眼转化成有机茶园和无机茶园后分别降低了16%和71%。在10~20 cm土层,有机茶园土壤速效磷(AP)提高了56%,而无机茶园的速效磷(AP)含量降低了65%。相似性分析表明,土地利用方式明显改变了土壤phoD微生物群落结构。但是,土层深度并没有改变phoD微生物群落结构。RDA分析表明,土壤pH、总钾(TK)、速效钾(AK)和硝态氮(NO3–-N)是驱动phoD功能微生物群落结构分异的主要环境因子。土壤速效磷(AP)含量与phoD功能微生物密切相关,其中Betaproteobacteria在调控土壤磷的有效性中发挥重要作用。研究结果表明,龙眼园转化成茶园改变了phoD微生物群落结构,同时降低了有机茶园土壤总磷(TP)和无机茶园土壤有效磷(AP)含量。与无机茶园相比,有机管理可以提高土壤速效磷(AP)含量。
关键词:土地利用;有机管理;phoD功能微生物;土壤速效磷;酸性磷酸酶
中图分类号:S151.9 文献标识码:A
Abstract: To investigate the effects of different land use types on phosphorous availability and phosphate-solubilizing bacterial community, the soil of conventional inorganic managed Longan orchard (LO), the organic managed tea plantation (OTP) transformed from Longan orchard and the conventional inorganic managed tea plantation (CTP) transformed from Longan orchard were studied in Baisha County, Hainan Island. In the study, the community structure of soil phoD functional microorganisms were determined by Illumina sequencing, and analyzed the correlation between soil AP and phoD functional microorganisms and acid phosphatase.After converting LO into CTP, the contents of TN, TP, AK, SOC and pH were significantly reduced in the 0-10 cm soil layer, and the contents of TN, TP, SOC and pH were significantly reduced in the 10-20 cm soil layer. After LO converting into OTP and CTP, the contents of soil AP decreased by 16% and 71% in the 0-10 cm soil layer, respectively. The content of soil AP was increased by 56% in the OTP and decreased by 65% in the CTP in the 10–20 cm soil layer. Moreover, the phoD functional microbial community structure was mainly affected by land use types, rather than soil depth. The RDA analysis revealed that soil pH, TK, AK and NO3–-N were the main influencing factors driving for phoD functional microbial community structure change. Meanwhile, the content of soil AP was closely related to phoD functional microorganisms and Betaproteobacteria. These results indicated that the transformation of LO into OTP and CTP changed the phoD microbial community structure and reduced the the content of TP in the soil of OTP and AP in the soil of CTP. Meanwhile, OTP could increase soil available phosphorus content compared with CTP.
Keywords: land use; organic management; phoD functional microorganisms; soil available phosphorus; acid phosphatase
DOI: 10.3969/j.issn.1000-2561.2021.12.035
磷是植物生长发育必需的营养元素[1]。热带地区高温、降雨等气候条件导致土壤高度风化,常表现土壤磷含量不足[2]。同时,土壤Fe3+、Al3+矿物对磷的固定以及通过径流、生物吸收等途径,也会加剧热带酸性土壤的磷限制[3]。因此,磷被认为是热带农业系统中影响作物生长和产量的重要限制因子[4-6]。土壤有機磷占总磷库的30%~ 80%[7],有机磷通过磷酸酶水解成速效磷才能被植物和微生物利用[8-9]。因此,矿化土壤有机磷是提高农田土壤有效磷,维持热带农业可持续发展必要和有效的途径。
土壤微生物是土壤有机磷转化过程的主要驱动者。携带编码磷酸酶基因的phoD溶磷细菌是参与磷循环的重要功能类群[10]。当土壤缺磷时,溶磷细菌通过分泌有机酸,产生磷的水解酶以活化难溶性磷,进而影响土壤有机磷的转化过程。研究发现,phoD功能微生物的多样性和组成对土地利用变化[11],肥料的添加[12-14]以及有机物料的投入[15-16]都非常敏感。然而,土地利用变化主要聚焦于自然林或草地转变为农田的研究。近年来,由于天然林的保护和利益驱动,越来越多的低经济作物种植园转换成高经济作物种植园。这样的土地利用变化方式已经成为了农业生产的主要土地利用变化方式,而这种土地利用方式变化如何影响phoD功能微生物群落的研究还鲜见报道。另外,与传统农业相比,有机农业在生产中不使用人工合成的肥料、农药、生长调节剂等,而是通过施用有机肥满足作物营养需求。有机管理也是影响土壤磷可利用性的重要农业管理措施。目前,关于有机管理对土壤磷素形态特征、含量变化和土壤微生物群落研究较多[17],然而,有机管理对phoD功能微生物群落和多样性的影响尚待讨论。
茶是我国热带和亚热带重要的经济树种,经济效益较高。近些年,随着龙眼老龄化和价格下降等诸多原因,大面积的龙眼园被茶园所取代,这一过程造成土壤肥力和土壤微生物数量下降[15]。基于此,本研究通过对由传统无机管理龙眼园转化为传统无机管理和有机管理茶园的田间定位实验,探究不同土地利用方式对土壤速效磷、酸性磷酸酶及phoD功能微生物的影响。主要回答以下3个问题:(1)phoD功能微生物群落与土壤有效磷关系如何?(2)龙眼转化成无机茶园对土壤速效磷是有利还是不利?(3)如果不利,有机管理是否能通过改变phoD功能微生物群落,提高土壤速效磷含量。
1 材料与方法
1.1 材料
研究区位于海南省白沙县(109°38′ E,19°18′ N),属热带季风气候,年均降水量约为1900 mm,年均气温22.7 ℃,地势平坦,土壤类型为砖红壤。在土地转换前,该地均以相同的传统无机管理方式种植龙眼。部分龙眼园于2014年转换成了有机茶园和传统无机茶园,因此,该试验地土壤性质差异主要是由该区种植作物和土地转换后灌溉,施肥和管理等措施的变化导致。有关实验设计详见课题组之前发表的Wu等[17]的文章。
本研究选取相邻的龙眼园和不同管理方式的茶园,并基于植物生长营养需求其施肥量为:传统无机龙眼园(LO, Longan orchard),平均每年尿素、P2O5、K2O施用量分别为360、300、240 kg/hm2;有机茶园(OTP, organic tea planta-tions),平均每年干羊粪施用量为6000 kg/hm2,其中干羊粪中总有机碳(TOC)、全氮(TN)、全磷(TP)、有效钾(AK)及有效磷(AP)含量分别为270、8、6、5、0.46 g/kg,即平均每年总氮、总磷和速效钾的使用量为48、36、30 kg/hm2;传统无机茶园(CTP, conventional tea plantations),平均每年N、P2O5、K2O的施用量为450、225、225 kg/hm2。此外,与有机茶园禁用化肥与农药相比,传统无机管理方式还施用杀虫剂吡虫啉和溴氰菊酯,以及除草剂草甘膦和百草枯。总的来看,施肥量依次为传统无机茶园>龙眼种植茶园>有机茶园。
1.2 方法
1.2.1 样品采集 2017年7月20日采集土壤样品。每个种植园建立3个20 m×20 m的样方。除去地表杂物和表层浮土,采用五点法,在每个样方用直径为5 cm的土钻取0~10 cm,10~20 cm土层土壤样品,混匀后作为一个样方的土壤样品。土壤样品过2 mm筛后分为2份,一份存储于–80 ℃用于测定phoD功能微生物群落,另一份自然风干用于测定土壤基础理化性质。
1.2.2 土壤理化性质分析 采用电位法(水土体积比2.5∶1)测定土壤pH;TOC分析仪法测定土壤有机碳(SOC)含量;半微量开氏法测定全氮(TN)含量;H2SO4-HClO4消煮钼锑抗比色法测定全磷(TP)含量;NaOH熔融–火焰光度法测定土壤全钾(TK)含量;紫外分光光度法测定硝态氮(NO3–-N)和铵态氮(NH4+-N)含量;0.5 mol/L NaHCO3浸提钼锑抗比色法测定土壤有效磷(AP)含量[18];1 mol/L NH4OAc浸提-火焰光度法测定土壤有效钾(AK)含量[19]。
1.2.3 phoD功能微生物分析 土壤酸性磷酸酶活性(ACP)采用Saiya-Cork等[20]的方法,即用125 mL 50 mmol/L乙酸盐缓冲液(pH=5)提取1.0 g新鲜土壤,磁力搅拌器均化5 min后将200 μL提取液转移到96孔微板中,并以磷酸4-甲基伞形酮磷酸酯(MUP)为底物。将微孔板置于20 ℃恒温,黑暗环境中培养4 h后,使用多功能微板读取器(SpectraMax M5, molecular devices, USA)测定ACP活性。
采用土壤DNA分离试剂盒(MoBio, California, USA),从0.5 g土壤中提取分离土壤DNA,对phoD微生物采用ALPS-F730和ALPS-R110引物进行PCR扩增,在Illumina MiSeq platform(illumina, san diego, USA)平台进行高通量测序,基于对GenBank数据库中phoD序列数据的相似度搜索,以97%的相似度将数据聚类为不同的操作分类单元(OTUs)。
1.3 数据处理
利用单因素方差分析(one-way ANOVA)和多重比较(duncan test)分析不同处理土壤理化性质和土壤酶活性的差异显著性,通过独立样本T(T-test)检验分析土层间土壤理化性质与土壤胞外酶的差异显著性。采用一般线性模型(GLM)明确土地利用方式和农业管理模式、土层以及二者的交互作用(SPSS 17.0 for Windows, SPSS, Inc.)。利用R进行相似性分析(ANOSIM)比较不同土地利用方式和农业管理模式下phoD微生物群落的结构差异性(R development core team 2016)。冗余分析(RDA)用于明确不同土地利用方式和农业管理模式下土壤理化因子与phoD功能微生物群落之间的关系(CANOCO 5.02)。
2 结果与分析
2.1 土地利用方式对土壤理化性质的影响
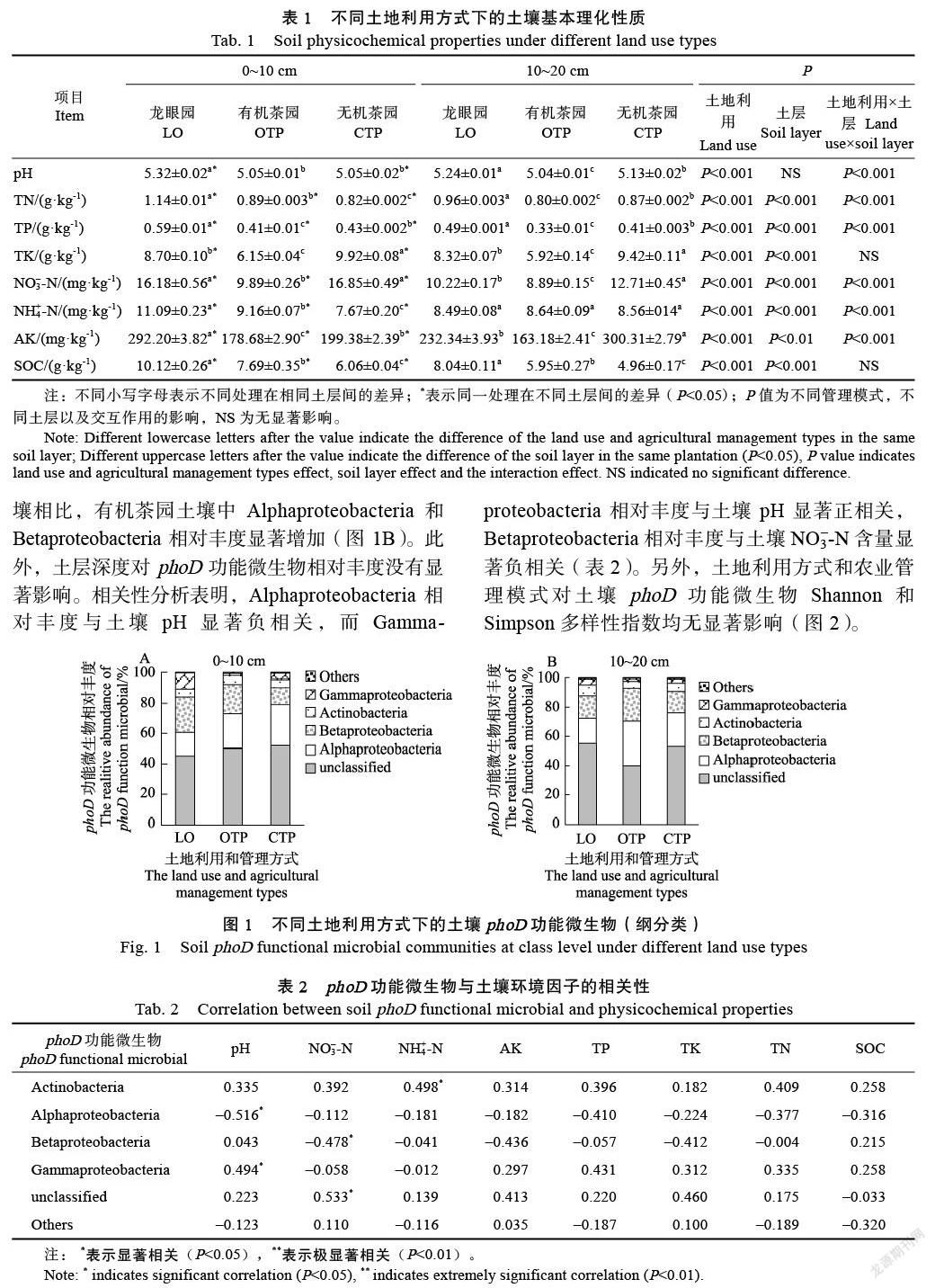
在0~10 cm土层中,龙眼园转化为有机和无机茶园后,除NO3–-N和TK外,其余土壤理化参数均显著降低。与无机茶园土壤相比,有机茶园土壤中TN、SOC和NH4+-N含量显著增加,而TP、TK和AK含量显著降低。在10~20 cm土层中,龙眼园转化为有机和无机茶园后,土壤中pH、TN、TP以及SOC含量均显著降低。与无机茶园土壤相比,有机茶园土壤中除SOC含量显著增加,NH4+-N含量无显著变化外,其他土壤理化参数均显著降低(表1)。
在龙眼园中,0~10 cm土层土壤理化参数均显著高于10~20 cm土层。有机茶园中,0~10 cm土层土壤的TN、TP、NO3–-N、NH4+-N、AK以及SOC含量显著高于10~20 cm土层土壤。无机茶园中,0~10 cm土层土壤的TP、TK、NO3–-N以及SOC含量显著高于10~20 cm土层土壤,而pH、TN、NH4+-N以及AK含量显著低于10~20 cm土层土壤。此外,除TK和SOC外,不同土地利用和管理措施与土层交互作用对其他土壤理化性均有显著影响(P<0.001,表1)。
2.2 土地利用方式对phoD功能微生物群落的影响
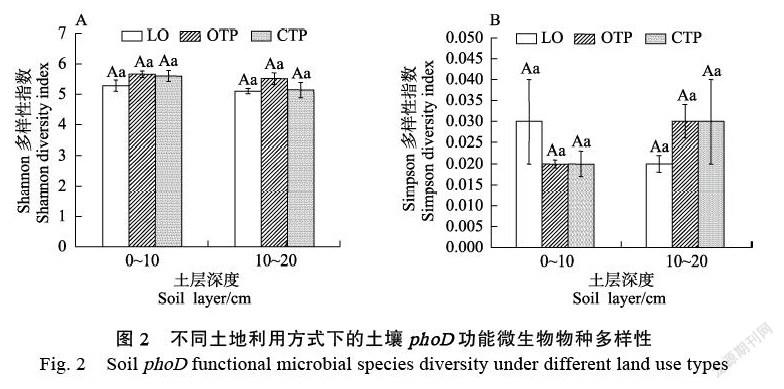
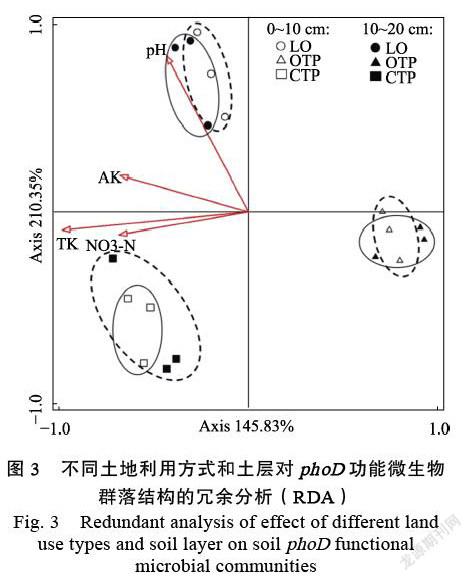
在0~10 cm土层中,由龙眼园转化为有机和无机茶园后,土壤中Gammaproteobacteria相对丰度均显著降低。Betaproteobacteria相对丰度变化依次为龙眼园>有机茶园>无机茶园(图1A)。在10~20 cm土层中,由龙眼园转化成有机茶园后,土壤中Alphaproteobacteria和Betaproteobacteria相对丰度显著增加,而转化成无机茶园后,土壤phoD功能微生物相对丰度并无显著差异。与无机茶园土壤相比,有机茶园土壤中Alphaproteobacteria和Betaproteobacteria相对丰度显著增加(图1B)。此外,土层深度对phoD功能微生物相对丰度没有显著影响。相关性分析表明,Alphaproteobacteria相对丰度与土壤pH显著负相关,而Gamma-proteobacteria相对丰度与土壤pH显著正相关,Betaproteobacteria相对丰度与土壤NO3–-N含量显著负相关(表2)。另外,土地利用方式和农业管理模式对土壤phoD功能微生物Shannon和Simpson多样性指数均无显著影响(图2)。
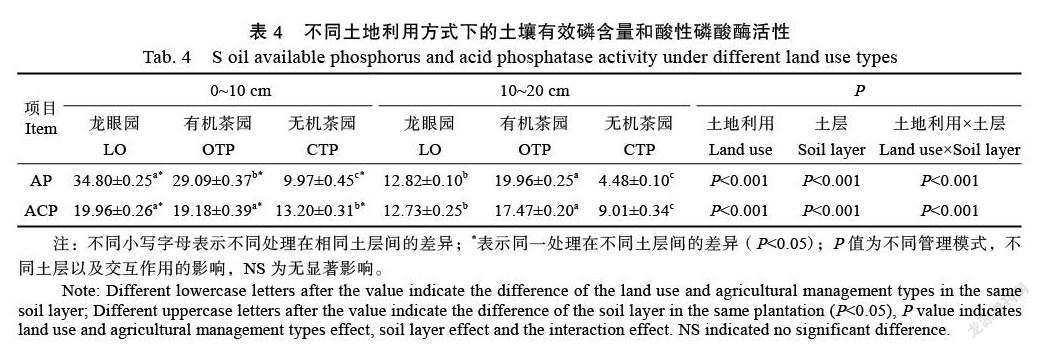
相似性分析表明(表3),土壤phoD功能微生物群落结构变化主要是由不同土地利用和农业管理模式导致(ANOSIM,R=0.625,P=0.001),而与土层深度无关(ANOSIM,R=–0.111,P=1.000)。同时,RDA结果显示,在不同土地利用和农业管理模式下,土壤phoD功能微生物群落具有显著差异,而在不同土层深度时没有显著差异(图3)。土壤phoD功能微生物群落主要受到土壤pH、TK、AK和NO3–-N含量的影响(P<0.05),共解释了微生物群落结构56.18%的变异,其中第一軸解释了变异的45.83%,第二轴解释了另外的10.35%。
2.3 土地利用方式对土壤有效磷和酸性磷酸酶的影响
0~10 cm土层中,龙眼园转化为有机和无机茶园后,土壤AP含量分别降低了16%和71%。此外,龙眼园在转化为有机茶园后,土壤ACP活性并无显著改变,而在转化为无机茶园后,土壤ACP活性降低了34%。与无机茶园土壤相比,有机茶园中土壤ACP活性增加了45%。在10~20 cm土层中,龙眼园转化为有机茶园后土壤AP含量提高了56%,ACP提高了37%。可是,与龙眼园相比,无机茶园土壤AP含量与ACP活性均显著降低,土壤AP和ACP活性含量分别降低了65%和29%(表4)。
2.4 土壤速效磷、酸性磷酸酶与土壤理化性质和phoD功能微生物群落的相关性
在0~10 cm土层中,土壤AP含量和ACP活性与NH4+-N、TN和SOC含量均呈显著正相关关系。土壤AP含量和ACP活性与phoD功能微生物群落中Betaproteobacteria相对丰度呈显著正相关。在10~20 cm土层中,土壤AP含量和ACP活性与Betaproteobacteria相对丰度呈显著正相关,而与NO3–-N、TK以及AK含量呈显著负相关。此外,2个土层土壤的AP含量与ACP活性呈显著正相关(表5,表6)。
3 讨论
3.1 不同土地利用方式下土壤phoD功能微生物相对丰度、群落结构及多样性变化
土壤phoD功能微生物在磷素转化过程中起着重要作用。本研究发现,龙眼园转换成茶园后,在0~10 cm土层中,无机和有机茶园土壤中Gammaproteobacteria相对丰度均显著降低,Betaproteobacteria相对丰度变化依次为龙眼园>有机茶园>无机茶园(图1A)。相关性分析表明土壤pH与Alphaproteobacteria和Gammaproteobacteria相对丰度显著正相关。龙眼园转化成茶园后,茶树根系分泌物中含有大量的有机酸,如草酸、柠檬酸和苹果酸能够使茶树种植园中土壤pH下降[21-22],这可能导致了土壤有机质分解过程发生差异,例如,不同C/N值的有机物发生改变[23],资源底物的改变会引起微生物群落结构变化,进而导致Alphaproteobacteria和Gammaproteobacteria相对丰度的降低。而Betaproteobacteria群落与土壤NO3–-N含量呈显著负相关(表2)。这主要是由于Betaproteobacteria中大多微生物属于厌氧反硝化微生物,在NO3–-N含量较高的环境中有较低的丰度[24]。与施用化肥相比,本研究所施用的有机肥为腐熟的干羊粪,其含有的纤维素和半纤维素,
不易被分解,并且释放的养分很容易被吸收,导致有机茶园土壤中残留的NO3–-N含量减少,因此使得有机茶园土壤中Betaproteobacteria相对丰度显著增加。另外,Gammaproteobacteria和Betaproteobacteria中许多菌属具有增强磷酸盐溶解以促进植物生长的作用,如Pseudomonas、Hydrogenophaga等[25-26],这类细菌在土壤碳磷循环中发挥重要作用,因此在0~10 cm土层中,Gammaproteobacteria和Betaproteobacteria相对丰度显著降低表明龙眼园转化成有机和无机茶园后不利于土壤有机磷的转化。而与无机茶园相比,其在有机茶园高的相对丰度表明有机管理更有利于有机磷的转化。在10~20 cm土层中,龙眼园转化成茶园有机茶园后,土壤中Alphaproteobacteria和Betaproteobacteria的相对丰度均显著增加,而无机茶园土壤中phoD功能微生物并无显著变化。另外,土壤中Alphaproteobacteria和Betaproteobacteria的相对丰度表现为有机茶园>无机茶园(图1B)。Alphaproteobacteria中具有许多能够产生酸性磷酸酶的菌属,如Roseivivax能够释放酸性磷酸酶加速土壤有机磷的转化[27],有机茶园土壤高的Alphaproteobacteria和Betaproteobacteria丰度表明由龙眼园转化成有机茶园有利于亚表层土壤磷的矿化。与无机管理相比,有机管理更有利于酸性磷酸酶的释放进而促进土壤有机磷矿化。
土壤微生物群落结构和组成受到土壤理化性质的显著影响[28]。本研究中土壤pH是驱动phoD功能微生物群落变化的主要环境因子(图3),这与前人研究相似[29-30]。在无机茶园中,由于无机管理长期施用化肥会导致土壤板结、酸化,抑制微生物生长,而有机肥、有机物料的的施用能够改善土壤结构和养分状况,为微生物生长提供充足的碳源、氮源以及较佳的环境,这可能是导致有机和无机管理措施下土壤phoD功能微生物群落结构出现差异的原因[31-32]。这与Sakurai等[33]在农田耕地中的研究结果一致。另外,农药与除草剂的施用对微生物组成也有负面的影响。土地利用方式虽然显著改变了phoD功能微生物的群落结构,但是并没有改变其多样性指数,这与以往大多数研究不同[34-35],可能的原因在于以往研究多以森林生态系统转化为农田生态系统为主。森林生态系统由于人为干扰少,拥有较高的物种多样性,在转化为农田生态系统后,会降低其微生物多样性指数,这表明土壤微生物多样性与植物多样性息息相关[36]。而本研究均是以单一种植作物的农田生态系统转化为另外单一种植作物的农田生态系统,这可能导致其phoD功能微生物多样性指数无显著变化。
3.2 不同土地利用方式下土壤磷素特征
不同土地利用方式可以影响土壤磷素的状况。土壤速效磷是用于反应土壤微生物和植物对磷素需求的有效指标[37]。本研究中,在0~10 cm土层时,龙眼园转化为茶园后,土壤总磷和速效磷含量显著下降。土壤磷含量取决于植物凋落物分解和植物吸收的平衡关系。龙眼树属于高大常绿乔木,茶树则是以收获茶叶为主的小乔木经济作物,茶园的凋落物数量远远低于龙眼园,在龙眼园转化为茶園后,其土壤有机物的输入显著减少。另外,幼龄茶树生长旺盛,对土壤磷的需求较大,从而导致了茶园土壤中总磷和速效磷含量降低[38-39],这表明龙眼园转化为茶园后减少了土壤磷含量。虽然传统无机茶园比有机茶园土壤含有更高的总磷,但有机茶园土壤速效磷含量却显著高于传统无机茶园,这是因为有机茶园具有较
高的磷酸酶活性能矿化有机质释放磷。本研究中,尽管无机管理模式施用了更多的磷肥,但是磷主要以Ca-P和闭蓄态磷形式存在,导致无机茶园中速效磷含量较低[17]。这可能说明与有机茶园相比,无机茶园需要投入大量的磷肥才能保证茶树正常生长,从而可能增加土壤磷养分流失风险。在10~20 cm土层中,与龙眼园和无机茶园相比,有机茶园表现出了较高的酸性磷酸酶活性。由于施用有机肥能够改善亚表层土壤结构,提高微生物对磷的吸收,促进酸性磷酸酶的释放[40],进而使得有机茶园中10~20 cm土层有较高的速效磷含量。同时,土壤速效磷含量与Betaproteobacteria相对丰度呈显著正相关(表6)。这与Luo等[41-42]的研究一致,Betaproteobacteria属于生长速度相对较快的菌,因此能够产生更多的磷酸酶来有效的促进土有机磷的矿化,这表明有机管理可通过调节phoD功能微生物群落结构,增加Betaproteobacteria的相对丰度来分泌更多的酸性磷酸酶来促进土壤有机磷矿化成无机磷,缓解茶树生长的磷限制,进而提高土壤磷利用率。在不
同土层中,龙眼园、有机茶园和无机茶园土壤总磷和速效磷含量均表现为在0~10 cm土层显著高于10~20 cm土层。这可能的原因是0~10 cm土层有机质含量更高,这些有机质占据了磷的吸附点,降低了磷的吸附,使得土壤释放了更多的磷。另外,在0~10 cm土层时,土壤微生物直接参与了土壤磷的转化,而在10~20 cm土层时,土壤微生物可能通过影响土壤理化性质而间接影响磷的转化,这导致10~20 cm土层土壤磷转化变缓,使得磷的储存降低[17]。
4 结论
在龙眼园转换成有机和无机茶园后,显著降低了土壤总磷和速效磷,表明龙眼转化成茶园对土壤磷的生物可利用性产生负面效应。同时,土地利用方式的转变改变了土壤phoD功能微生物群落结构,其中Betaproteobacteria相对丰度与土壤速效磷含量显著正相关,表明Betaproteobacteria在调控土壤磷的有效性中发挥了重要作用。另外,与无机茶园相比,有机茶园通过调节phoD功能微生物群落显著提高了土壤速效磷含量,这表明有机管理能在一定程度上提高土壤磷的供应能力。
参考文献
[1] Vitousek P, Porder S, Houlton B, et al. Terrestrial phospho-rus limitation: mechanisms, implications, and nitrogen- phosphorus interactions[J]. Ecological Applications, 2010, 20(1): 5-15.
[2] Wang H, Liu S, Zhang X, et al. Nitrogen addition reduces soil bacterial richness, while phosphorus addition alters community composition in an old-growth N-rich tropical forest in Southern China[J]. Soil Biology and Biochemistry, 2018, 127: 22-30.
[3] Marklein A R, Houlton B Z. Nitrogen inputs accelerate phosphorus cycling rates across a wide variety of terrestrial ecosystems[J]. New Phytologist, 2012, 193(3): 696-704.
[4] Wang J, Ren C, Cheng H, et al. Conversion of rainforest into agroforestry and monoculture plantation in China: Consequences for soil phosphorus forms and microbial community[J]. Science of the Total Environment, 2017, 595: 769-778.
[5] Vitousek P M, Sanford Jr R L. Nutrient cycling in moist tropical forest[J]. Annual review of Ecology and Systematics, 1986, 17(1): 137-167.
[6] Cross A F, Schlesinger W H. A literature review and evalua-tion of the. Hedley fractionation: Applications to the bio-geochemical cycle of soil phosphorus in natural ecosys-tems[J]. Geoderma, 1995, 64(3-4): 197-214.
[7] 趙少华, 宇万太, 张 璐, 等. 土壤有机磷研究进展[J]. 应用生态学报, 2004, 15(11): 2189-2194.
[8] Richardson A. Prospects for using soil microorganisms to improve the acquisition of phosphorus by plants[J]. Australian Journal of Plant Physiology, 2001, 28(9): 897-906.
[9] Nannipieri P, Giagnoni L, Renella G, et al. Soil enzymology: classical and molecular approaches[J]. Biology and Fertility of Soils, 2012, 48(7): 743-762.
[10] Ragot S, Kertesz M, Bunemann E. phoD Alkaline phospha-tase gene diversity in soil[J]. Applied and Environmental Microbiology, 2015, 81(20): 7281-7289.
[11] Neal A L, Rossmann M, Brearley C, et al. Land‐use influ-ences phosphatase gene microdiversity in soils[J]. Environmental Microbiology, 2017, 19(7): 2740-2753.
[12] Luo G, Ling N, Nannipieri P, et al. Long-term fertilisation regimes affect the composition of the alkaline phosphomo-noesterase encoding microbial community of a vertisol and its derivative soil fractions[J]. Biology and Fertility of Soils, 2017, 53(4): 375-388.
[13] Chen X D, Jiang N, Condron L M, et al. Impact of long-term phosphorus fertilizer inputs on bacterial phoD gene community in a maize field, Northeast China[J]. Science of The Total Environment, 2019, 669(4): 1011-1018.
[14] Chen X D, Jiang N, Condron L M, et al. Soil alkaline phos-phatase activity and bacterial phoD gene abundance and di-versity under long-term nitrogen and manure inputs[J]. Geoderma, 2019, 349(4): 36-44.
[15] Sun Q, Qiu H, Hu Y, et al. Cellulose and lignin regulate partitioning of soil phosphorus fractions and alkaline phos-phomonoesterase encoding bacterial community in phos-phorus-deficient soils[J]. Biology and Fertility of Soils, 2019, 55(1): 31-42.
[16] Xie Y Y, Wang F H, Wang K, et al. Responses of bacterial phoD gene abundance and diversity to crop rotation and feedbacks to phosphorus uptake in wheat[J]. Applied Soil Ecology, 2020, 154: 103604.
[17] Wu T T, Liu W J, Wang D G, et al. Organic management improves soil phosphorus availability and microbial properties in a tea plantation after land conversion from longan (Dimocarpus longan)[J]. Applied Soil Ecology, 2020, 154.
[18] Olsen S R. Estimation of available phosphorus in soils by extraction with sodium bicarbonate[M]. US Department of Agriculture, 1954.
[19] 中國科学院南京土壤研究所. 土壤理化分析[M]. 上海: 上海科学技术出版社, 1978.
[20] Saiya-Cork K R, Sinsabaugh R L, Zak D R. The effects of long term nitrogen deposition on extracellular enzyme activity in an Acer saccharum forest soil[J]. Soil Biology and Biochemistry, 2002, 34(9): 1309-1315.
[21] Wang X P. The introductory study of tea roots exuding organic acids[J]. Journal of Tea Science, 1994, 14: 17-22.
[22] 杨亚军. 中国茶树栽培学[M]. 上海: 上海科学技术出版社, 1986.
[23] Schnitzer M. Effect of low pH on the chemical structure and reactions of humic substances[M]//Effects of acid precipitation on terrestrial ecosystems. Boston, MA: Springer, 1980: 203-222.
[24] 高文萱, 闫建华, 杜会英, 等. 土壤nirS、nosZ型反硝化菌群落结构及多样性对牛场肥水灌溉水平的响应[J]. 农业环境科学学报, 2019, 38(5): 1089-1100.
[25] Alori E T, Glick B R, Babalola O O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture[J]. Frontiers in microbiology, 2017, 8: 971.
[26] Giles C D, Hsu P C L, Richardson A E, et al. Plant assimila-tion of phosphorus from an insoluble organic form is im-proved by addition of an organic anion producing Pseudo-monas sp[J]. Soil Biology and Biochemistry, 2014, 68: 263-269.
[27] Chen M H, Sheu S Y, Chen C A, et al. Roseivivax isoporae sp. nov., isolated from a reef-building coral, and emended description of the genus Roseivivax[J]. International journal of systematic and evolutionary microbiology, 2012, 62(6): 1259-1264.
[28] Dequiedt S, Saby N P A, Lelievre M, et al. Biogeographical patterns of soil molecular microbial biomass as influenced by soil characteristics and management[J]. Global Ecology and Biogeography, 2011, 20(4): 641-652.
[29] Wang Y, Zhang F, Marschner P. Soil pH is the main factor influencing growth and rhizosphere properties of wheat fol-lowing different pre-crops[J]. Plant and Soil, 2012, 360(1-2): 271-286.
[30] Kang E, Li Y, Zhang X, et al. Soil pH and nutrients shape the vertical distribution of microbial communities in an al-pine wetland[J]. Science of The Total Environment, 2021: 145780.
[31] Stark C, Condron L M, Stewart A, et al. Influence of organic and mineral amendments on microbial soil properties and processes[J]. Applied Soil Ecology, 2007, 35(1): 79-93.
[32] Manjunath M, Kumar U, Yadava R B, et al. Influence of organic and inorganic sources of nutrients on the functional diversity of microbial communities in the vegetable cropping system of the Indo-Gangetic plains[J]. Comptes Rendus Biologies, 2018, 341(6): 349-357.
[33] Sakurai M, Wasaki J, Tomizawa Y, et al. Analysis of bac-terial communities on alkaline phosphatase genes in soil supplied with organic matter[J]. Soil Science and Plant Nu-trition, 2008, 54(1): 62-71.
[34] 王 蕊, 吴 宪, 李 刚,等. 东北丘陵区林地转型耕地对土壤编码碱性磷酸酶基因的细菌群落的影响[J]. 微生物学报, 2020, 60(10): 2211-2225.
[35] Vershinina O A, Znamenskaya L V. The Pho regulons of bacteria[J]. Microbiology, 2002, 71(5): 497-511.
[36] Bahram M, Netherway T, Hildebrand F, et al. Plant nu-trient-acquisition strategies drive topsoil microbiome struc-ture and function[J]. New Phytologist, 2020, 227(4): 1189-1199.
[37] Fan Y, Zhong X, Lin F, et al. Responses of soil phos-phorus fractions after nitrogen addition in a subtropical forest ecosystem: Insights from decreased Fe and Al oxides and increased plant roots[J]. Geoderma, 2019, 337: 246-255.
[38] De Schrijver A, Vesterdal L, Hansen K, et al. Four decades of post-agricultural forest development have caused major redistributions of soil phosphorus fractions[J]. Oecologia, 2012, 169(1): 221-234.
[39] Barbier S, Gosselin F, Balandier P. Influence of tree species on understory vegetation diversity and mechanisms in-volved-A critical review for temperate and boreal forests[J]. Forest Ecology and Management, 2008, 254(1): 1-15.
[40] Katsalirou E, Deng S, Gerakis A, et al. Long-term manage-ment effects on soil P, microbial biomass P, and phosphatase activities in prairie soils[J]. European Journal of Soil Biology, 2016, 76: 61-69.
[41] Luo G, Sun B, Li L, et al. Understanding how long-term organic amendments increase soil phosphatase activities: insight into phoD-and phoC-harboring functional microbial populations[J]. Soil Biology and Biochemistry, 2019, 139: 107632.
[42] Philippot L, Raaijmakers J M, Lemanceau P, et al. Going back to the roots: the microbial ecology of the rhizosphere[J]. Nature Reviews Microbiology, 2013, 11(11): 789-799.
責任编辑:白 净
猜你喜欢
大气科学学报(2022年3期)2022-07-22
环球人文地理·评论版(2016年5期)2017-01-03
南水北调与水利科技(2016年5期)2016-12-27
山东农业科学(2016年11期)2016-12-17
现代经济信息(2016年27期)2016-12-16
商(2016年33期)2016-11-24
中国集体经济(2016年27期)2016-11-19
商(2016年30期)2016-11-09
中国市场(2016年34期)2016-10-15
