
苜蓿、小冠花和百脉根花粉管引导和胚珠败育研究*
2021-01-22 09:23王晓娟黄利春张树振
云南农业大学学报(自然科学) 2021年1期
王晓娟,黄利春,张树振
(1.上海科技馆,上海自然博物馆自然史研究中心,上海 200127;2.科尔沁右翼前旗农业技术推广中心,内蒙古 科尔沁右翼前旗 137713;3.新疆农业大学 草业与环境科学学院,新疆 乌鲁木齐 830052)
作为蝶形花植物的典型代表,豆科牧草大多为雌雄同花但异花授粉,结实依赖传粉昆虫。围绕豆科牧草实际种子产量远低于潜在种子产量这一问题,前人提出昆虫传粉是制约其种子产量的重要因素[1]。传粉是一个复杂的过程,可概括为花粉释放—转运—降落3 个动态过程。花粉释放具有一定的时空效应,花药开裂是花粉释放的前提,昆虫作用是花粉释放的必要条件,花粉运输必须借助传粉昆虫,而花粉的沉降同样离不开传粉者的参与。因此,花粉来源及其活力可能影响种子形成,间接制约种子的产量[2]。由于雌配子体依次被包埋在胚珠、子房中,因此,被子植物在进化过程中形成了一种特殊的运送雄配子的方式—花粉管伸长和引导,即包含有2 个精细胞和1 个营养核的花粉管从雌蕊柱头定向延伸,将携带的2 个精子送达胚珠从而实现双受精[3]。通常,花粉到达柱头后经过分泌物水合,继而萌发生长,在引导组织的引导下,最终到达胚囊与胚珠结合,完成双受精。研究发现:一些常见豆科牧草胚珠败育严重,如紫花苜蓿(Medicago sativa)实际种子产量仅为潜在种子产量的4%,自然授粉条件下每小花平均胚珠数达13 个,结籽数却仅为2~3 个[4-5]。BOS 等[6]认为实际种子产量低于潜在种子产量是豆科牧草为了适应生存而逐渐形成的繁殖策略。
前人研究发现:大部分植物花的胚珠数都要多于其种子数[7]。针对这种现象存在2 种不同假说:一是广泛选择也称选择性败育假说,通过开花样式增加对传粉者的吸引,通过选择高质量的后代提高雌性适合度;二是雄性功能假说,通过高质量的花粉传播提高雄性适合度,这样可以补充因植食作用、天气或其他外界因素对花的破坏,保证资源和花粉的可利用性[8]。也有学者提出了花粉限制机制和资源限制机制2 种假说,前者强调因没有足够数量的花粉传递到柱头上而引起结实率低下,后者则指出没有足够多的营养资源供果实和种子发育成熟导致种子败育[9]。依据上述假说,植物形成多的花是一种“赌注策略”,其目的是在没有花粉限制和资源限制的条件下产生更多的后代[6]。
尽管以上观点侧重点不同,但是依然能发现二者之间的共同点:都强调了花粉因素,在传粉过程中花粉量不足可能制约种子产量,胚珠败育可能间接反映了花粉量不足。然而,对于大部分植物而言,导致果实与花(花器官)比率低下的原因还知之甚少,对豆科牧草多胚珠和低结实率现象还少有关注,有关授粉后的花粉管引导和受精后胚胎发育的基础数据缺乏。目前,关于这一问题的讨论仍然是植物繁殖生物学研究的热点。DUAN 等[10]研究揭示了拟南芥(Arabidopsis thaliana)中受体激酶(FERONIA) 具有确保精子递送和阻止多重受精的作用,FERONIA 通过一系列功能上相互联系的控制过程,阻止多个花粉管进入雌配子体。另有研究关注枣(Ziziphus jujuba)和长瓣短柱茶(Camellia grijsii)的花粉管引导,探讨其结实障碍问题[11-12]。前期本课题组在开展苜蓿花粉和胚珠遗传变异[5]、传粉昆虫访花行为及其环境互作[13-14]、花部适应机制和传粉系统[15]、花粉管极性引导[16]以及雌、雄配子体发育和胚胎发生[17-18]等繁殖生物学研究过程中,发现常见豆科牧草种子生产依靠传粉昆虫,但胚珠败育严重影响了每荚种子数,实际种子产量远低于理论种子产量这一问题。
鉴于此,本研究围绕3 种常见豆科牧草紫花苜蓿(Medicago sativa)、多变小冠花(Coronilla varia)和百脉根(Lotus cornioulatous)的胚珠败育现象,通过异花授粉后不同时期3 种豆科牧草花粉管生长、引导和胚珠受精的显微观察,探究其胚珠败育发生的限制因素。
1 材料与方法
1.1 供试材料和研究地点
供试材料为多年生紫花苜蓿(M.sativa)、多变小冠花(C.varia)和百脉根(L.cornioulatous),栽种于甘肃武威河西实验站(E102°84′,N37°67′,海拔1 593 m),该地区年均日照总时间1 664~2 532 h,年平均降水量154.6 mm,年均气温8 ℃。其中,紫花苜蓿为总状花序,小冠花和百脉根为伞形花序,3 种豆科牧草都有无限开花习性,花期较长。紫花苜蓿开花期为5 月中下旬至7 月下旬,长达70 d 左右,6 月上旬至下旬为盛花期,单个小花完全开放可以持续7 d 左右。多变小冠花花期5 月中下旬至7 月下旬,花期长达50~60 d,6 月上旬至下旬为盛花期,单个小花的花期为10 d 左右。百脉根花期6 月初至7 月下旬,长达50 d 左右,6 月中旬至7 月上旬为盛花期,单个小花的花期为15 d 左右(图1)。
1.2 苜蓿、小冠花和百脉根的人工和自然授粉杂交处理
每种牧草在试验地随机标记5 株,于盛花期分别进行人工自交授粉、人工异交授粉和自然授粉,每株每个处理选择10 个花序,每个花序处理7~10 朵小花。人工授粉在晴天9:30—11:30和15:00—16:00 进行。其中,异交授粉保证父本的花粉离母株在2~5 m,且花粉量充足,一般选择3~5 朵小花授予母本的柱头上。授粉前对父本的花粉进行检验,保证花粉平均活力≥80%。
1.3 苜蓿、小冠花和百脉根柱头的可授性和角质层检测
根据3 种豆科牧草的开花特性,将其小花花期分为3 个阶段:花冠紧闭、花冠展开(花药未裂) 和花冠展开(花药开裂)。3 种豆科牧草每株随机选取5 朵小花,采用联苯胺—过氧化氢法对其柱头可授性进行检验,反应液配方为联苯胺(1%)∶过氧化氢(3%)∶水=4∶11∶22 (体积比)[19]。把小花的雌蕊解剖出来,放到载玻片上,滴加反应液后移到解剖镜下观察,如果立刻出现大量气泡说明柱头具有可授性,反之则没有可授性。3 种豆科牧草每个单株随机选取5 朵小花,将雌蕊解剖出来,放到载玻片上,用0.02%中性红染色,在解剖镜下观察,出现红色则说明存在角质层,反之则无角质层[20]。

图1 紫花苜蓿(a)、多变小冠花(b)和百脉根(c)的花Fig.1 Florets of M.sativa (a),C.varia (b) and L.cornioulatous (c)
1.4 苜蓿、小冠花和百脉根花粉管引导的荧光显微观察
在标记的植株上进行人工异花授粉,每个豆科牧草处理5 个花序,每个花序约8 朵小花。分别选取2、4、8、12、24 和48 h 后的小花,卡诺固定液固定24 h,观察前可转移保存在70%的酒精中。小心解剖出雌蕊,将雌蕊置于8 mol/L 的NaOH 溶液中透明5 h,蒸馏水冲洗2 次,再转移到含有0.1% 苯胺蓝的0.1 mol/L K3PO4溶液中,于5 ℃的黑暗环境中过夜染色,去除雌蕊,置于滴有50% 甘油和50% 0.5 mol/L 碳酸盐缓冲液(抗荧光淬灭),盖片,用手指均匀挤压,置于荧光显微镜(OLYMPUS BX51,Japan)下观察照相。每个小花观察4 个视野,统计胚珠的受精率,有花粉管进入胚珠则视为胚珠受精,没有花粉管到达的胚珠则视为未受精。
1.5 苜蓿、小冠花和百脉根胚珠败育位置及其败育率统计
在荚果成熟时分别对3 种牧草荚内种子的败育位置进行统计,每种植物统计100 个荚,败育位置分为上部、中部和下部3 个部位,连接荚果的部位是下部,荚果顶端是上部,中间部分统称中部。
1.6 统计分析
苜蓿、小冠花和百脉根胚珠受精率和胚珠败育率用平均数±标准误表示;3 种豆科牧草异花授粉后不同时间胚珠受精率和胚珠败育率采用SPSS 19.0 统计分析软件进行单因素方差分析(one-way ANOVA),利用最小显著差数法(LSD),当P<0.05 时,表明存在显著差异;当P<0.01时,存在极显著差异。
2 结果与分析
2.1 3 种豆科牧草的柱头可授性和柱头角质层
分别选取3 种豆科牧草3 个不同开放阶段的小花:花冠紧闭、花冠展开(花药未裂) 和花冠展开(花药开裂),柱头可授性试验结果发现:3 种豆科牧草在小花开放的各个阶段,柱头对联苯胺—过氧化氢都有积极的反应,即使是在第1 阶段小花还没有开放,花瓣紧闭时,柱头也会有零星气泡产生,不过量少而且速度极慢。到小花开放的第2 阶段,即花药裂开但花瓣内的雌雄合蕊还没有释放出来时,除小冠花外,另外2 种牧草的柱头反应强烈。到第3 阶段小花完全开放时,3 种牧草的柱头对联苯胺—过氧化氢反应都很强烈,柱头瞬间会产生大量的气泡,而且气泡很大。以上结果表明:花冠展开(花药开裂)阶段是3 种豆科牧草柱头授粉的最佳时间,同种豆科牧草的异花授粉花粉和柱头之间存在亲和性和柱头可授性(表1)。

表1 3 种豆科牧草的柱头可授性比较Tab.1 Comparisons of stigma receptivity in three leguminous forages
中性红染色主要是检测柱头角质层的破裂程度以表征柱头的可授性,柱头着色则说明其角质层破坏,只有角质层破坏后柱头分泌物才能溢出并黏附花粉[21]。对紫花苜蓿、多变小冠花和百脉根的柱头角质层进行中性红染色检验,发现不作任何处理的小花98%的柱头不能显色,用毛笔刷过的柱头有72%的柱头可以被染成红色,而自然授粉的则有88%的柱头可以被染色,说明3 种豆科牧草的柱头均存在机械屏障也就是柱头角质层(表1)。
2.2 授粉后3 种豆科牧草花粉管生长和引导显微观察
由图2 可见:苜蓿在授粉2 h 后少量花粉在柱头上萌发(图2a),但是还不能穿过花柱基部(图2b);4 h 后花粉萌发的数量明显增多(图2c),并且可见少量花粉管穿过花柱基部(图2d),但是尚未到达花柱基部的胚珠位置(图2e);授粉8 h后柱头上可见大量花粉萌发(图2f),穿过花柱的花粉管数量也在不断增加(图2g),而且有少数花粉管到达胚珠附近(图2h);授粉12 h 后,到达胚珠位置的花粉管增多(图2i);授粉4 h 后,可以观察到花粉管向胚珠珠孔位置的生长和引导(图2j~k);授粉48 h 后可见花粉管成功引导进入胚囊,双受精发生,胚胎发育启动(图2l)。

图2 苜蓿异花授粉后2~48 h 花粉管的生长和引导Fig.2 Growth and guidance of pollen tubes in M.sativa 2-48 h after cross pollination
图3 显示:小冠花在授粉2 h 后,柱头上的花粉尚未萌发(图3a),花柱道未见花粉管生长(图3b);在授粉4 h 后,花柱基部可见花粉管生长(图3c),并有花粉管已经到达胚珠位置(图3d),此时胚囊的受精率可达88.97%;在授粉8 h 后,可以观察到柱头上大量花粉萌发(图3e),随后花粉管生长穿过花柱基部到达胚珠(图3f),以及花粉管向胚珠珠孔端的生长和引导(图3g),此时,胚囊的受精率达到96.43%,此后,胚囊受精率基本不变(表2);授粉12~24 h,可以观察到每个胚珠都有花粉管的引导生长和进入珠孔端(图3h~i);授粉48 h 后可以明显看到胚胎的发育(图3j~l)。
图4 显示:百脉根在授粉2 h 后柱头上的花粉还没有萌发(图4a),花柱道和胚珠未见花粉管的分布(图4b);在授粉4 h 后,可以观察到百脉根柱头上花粉的萌发,并且可见少量花粉管向花柱道和胚珠方向的生长(图4c~d),此时胚囊的受精率可达89.81%;在授粉8 h 后,可见花粉管向花柱基部的生长(图4e)和花粉管在胚珠位置的分布(图4f),该阶段胚囊的受精率达95.38%,此后,胚囊受精率保持不变(表2)。与小冠花类似,授粉12~24 h,观察到大量花粉管向胚珠方向生长(图4g),花粉管向珠孔端的运动(图4h),以及花粉管向胚珠珠孔处的引导(图4i~k);授粉48 h 后,花粉管进入胚囊,并且每个胚珠都可以观察到花粉管的进入(图4l)。
3 种豆科牧草不同的授粉时间后花粉管陆续进入胚囊,苜蓿在授粉24 h 后花粉管的数量尽管和授粉48 和96 h 后没有显著差异,但是数量还在不断增加,在授粉96 h 后胚囊的受精率达76.40%,而小冠花和百脉根在授粉4 h 后胚囊的受精率分别达到88.97%和89.81%,授粉24 h 后几乎接近100%受精。总体来看3 种牧草的受精率都比较高,苜蓿相对小冠花和百脉根而言,受精率较低,但也可以满足>70%的胚囊受精,然而本结果并不与苜蓿种子成熟时较低的结实率相符。
2.3 3 种豆科牧草胚珠败育位置和败育率的比较
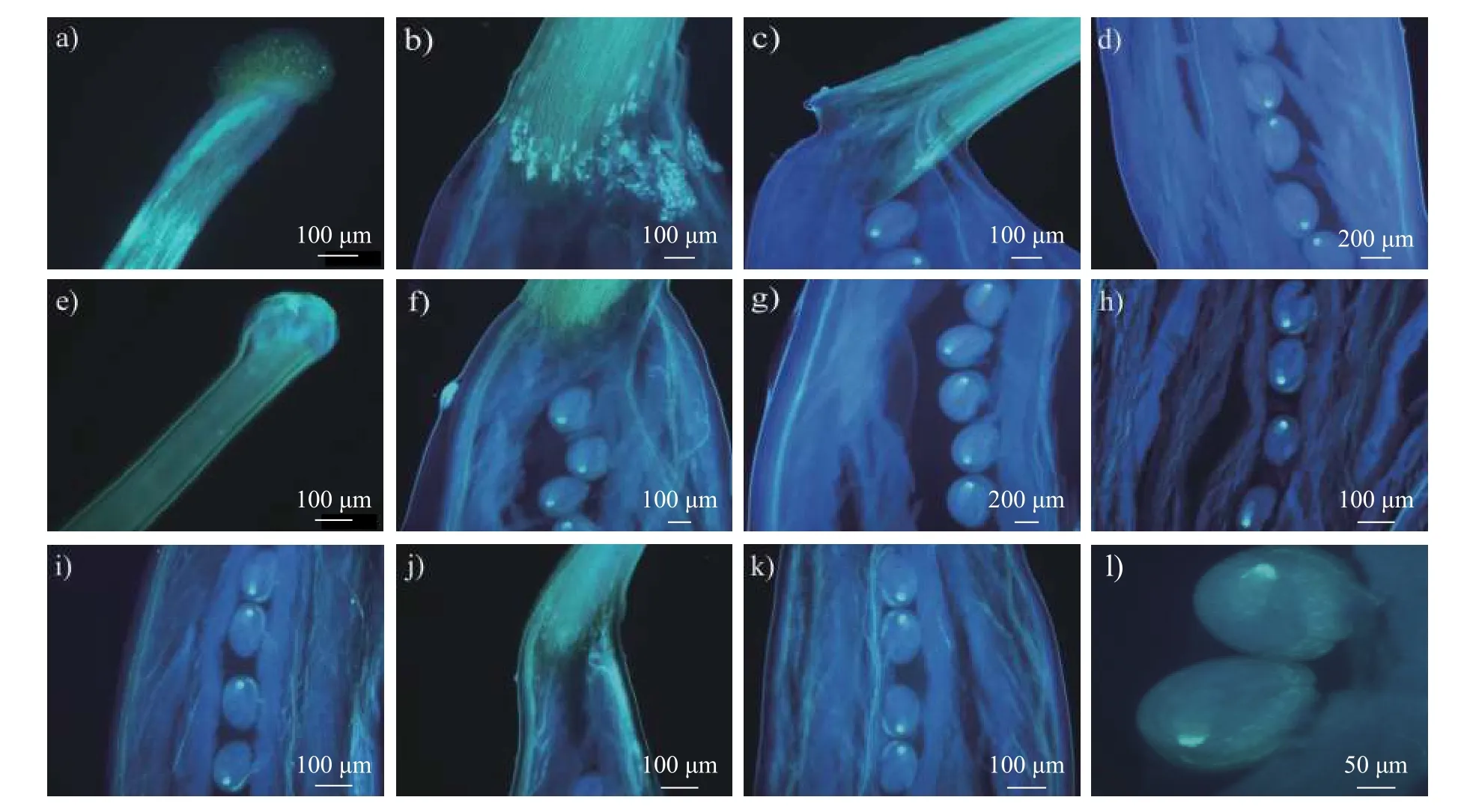
图3 小冠花异花授粉后2~48 h 花粉管的生长和引导Fig.3 Growth and guidance of pollen tubes in C.varia 2-48 h after cross pollination
3 种豆科牧草荚果成熟后对荚内种子的败育位置统计结果(表3)发现:苜蓿荚果上部败育率最高为94%,荚果中部和下部分别为88%和81%,3 个位置的败育率没有显著差异。小冠花荚果上部的败育率最高为82%,荚果中部和下部的败育率分别为74%和78%,3 个位置的败育率也没有显著差异;百脉根荚果中部的败育率最高为86%,荚果上部和荚果下部的败育率分别为76%和82%,3 个位置的败育率同样没有显著差异。以上结果表明:3 种牧草种子在荚内没有特定的败育位置,种子在荚内的败育位置具有随机性。
3 讨论
苜蓿、小冠花和百脉根作为重要的豆科牧草,其种子产量普遍低下,实际种子产量显著低于潜在种子产量,已有学者从传粉昆虫[1,22]、种子产量性状[23]等方面进行了探析,旨在解析异花授粉豆科牧草种子生产要素,促进产量提升。豆科牧草普遍存在的胚珠败育现象,与其授粉后花粉萌发、花粉管引导和胚珠受精过程紧密相关。豆科牧草属典型的湿(润)型柱头,成熟柱头表面呈现“蜂窝状”结构,并且柱头表面具有一层角质层,在柱头成熟过程中角质层降解,使位于角质层下方的分泌物泌出到柱头上,以利于黏附和识别花粉[18,21,24]。柱头角质层主要有2 个功能,一是可以阻挡自身的花粉,从而起到限制自花授粉的作用;二是角质层是一层机械组织,可以保护柱头,使柱头免受或减少外来的不良侵害。当柱头的角质层被破坏后花粉才会萌发生长,而传粉昆虫往往就是扮演这种“破坏者”的主要角色。本研究对苜蓿、小冠花和百脉根柱头检测结果显示:柱头在花瓣还没有完全开放时就具有一定的可授性,但可授性较低,当花药完全开裂时,柱头的可授性最强;其中,苜蓿和百脉根的柱头在花药还没有开裂时即具有可授性,当花药完全开裂时,3 种牧草的柱头可授性最佳(表1)。DAFNI等[25]认为花药完全开裂时正是传粉昆虫活动的高峰期,柱头上的角质层被破坏的概率最高,这样可以使传粉昆虫携带的花粉较容易突破角质层萌发。另外,植物花粉的萌发及花粉管生长与授粉当天的气候条件也存在密切的关系。柱头表皮由角质层覆盖,水和大分子物质可通过,但对花粉管穿透形成障碍。3 种豆科牧草花粉萌发和生长显微观察结果表明:来自异花授粉的花粉与柱头的水合反应、花粉管萌发可以很快发生,时间在1~2 h (图2~4)。这一方面与花粉在花药开裂时干化的程度有关;另一方面,恰当的花粉发育时期是接受花粉能力的关键时期,不成熟的柱头或衰老、退化的柱头都不能提供正常的授粉过程。

图4 百脉根的异花授粉后2~48 h 花粉管生长和引导Fig.4 Growth and guidance of pollen tubes in L.cornioulatous 2-48 h after cross pollination
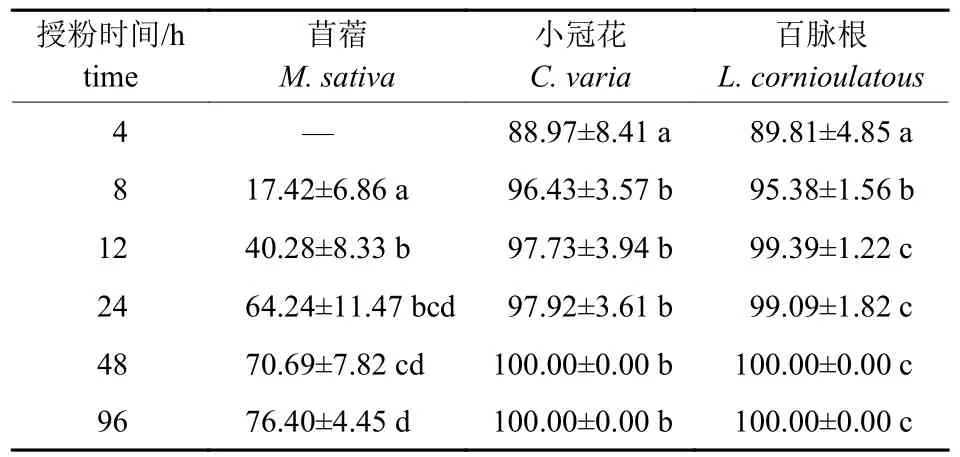
表2 3 种豆科牧草不同授粉时间胚珠受精率Tab.2 Ovule fertilization rate after cross pollination in three leguminous forages %

表3 3 种豆科牧草胚珠败育位置及其败育率Tab.3 Position of ovule abortion and ovule abortion rate in three leguminous forages %
一旦成熟的花粉落在柱头上,经过花粉与柱头的水合反应,花粉在柱头萌发,并穿过柱头乳突细胞间隙,花粉管伸向位于花柱和子房的引导组织。已有研究证明花粉管早期生长不受胚珠的影响,说明该阶段生长和引导是由孢子体控制的[26]。本研究观察发现:3 种豆科牧草花粉管生长的速度明显不同,尽管小冠花和百脉根花柱比苜蓿长,但是花粉管生长速度要比苜蓿快。苜蓿花粉管突破引导组织需要8 h,而小冠花和百脉根只需要4 h (图2~4)。在已有研究的其他植物中,矮慈姑(Sagittaria pygmaea)人工授粉0.5 h 后花粉萌发,花粉管在授粉2 h 后部分到达子房,4 h内进入胚珠,花粉管生长速率为1.0 mm/h[27];陆地棉(Gossypium hirsutum)花粉萌发在授粉后1~4 h、12 h 后大量的花粉管伸长到花柱基部,花粉管的平均生长速度为0.29 cm/h[28];辣椒(Capsicum annuum)从授粉到开始受精大约需要12 h,花粉管生长速度为0.8 mm/h,子房内胚珠全部完成受精需要26 h 以上[29];软条红梅花柱内异花授粉花粉管生长速度明显比自花授粉的快,前者花粉管能够到达花柱基部,平均速度为0.83 mm/h,而自花授粉的花粉管在花柱中部被抑制而停止生长[30]。
花柱是连接柱头和子房的柱状结构,是花粉管进入子房的通道。花粉萌发后花粉管的早期生长包括2 个主要过程,一是克服柱头表面的影响,二是找到引导组织。已有研究发现:花粉进入柱头后,在花柱中的生长除了传输通道中营养物质的供应外,还受到从花柱到胚珠梯度分布化学信号及其他信号物质的指引[31]。玉米中一种被称为花粉过敏原组I (β-expansins)的蛋白质能够促进花粉管侵入柱头组织[32]。另有研究发现:百合柱头中有诱导花粉管向药性的小碱性蛋白质(chemocyanin),能有效地引导携带精细胞的花粉管进入胚珠,该蛋白只在百合中有功能,而百合柱头中另一种蛋白SCA (由百合引导组织表皮分泌的脂质转运蛋白,lipid transfer protein)可明显促进小碱性蛋白质对花粉管的引导[33]。从图2~4可以看出:苜蓿、小冠花和百脉根花柱的中央均有数条纵行的花柱道,自柱头通向子房,属空心型花柱类型。3 种豆科牧草的花柱沟道表面为高度腺性的花柱道细胞,所产生的黏性分泌物释放到花柱道的表面,传粉后花粉粒萌发所形成的花粉管沿着花柱道并吸收、利用花柱道表面的分泌物营养自身而进一步生长、穿行。由此可见,不同植物的花粉管生长速度不同,甚至同一牧草的不同品种(基因型)之间也有一定的差别[34]。
花粉管在花柱中的生长会受到来自胚珠的信号指引,包括长距离指引和短距离指引。双子叶植物花粉管穿过引导组织出现在胎座表层,然后沿着珠柄的表皮向珠孔端生长进入胚珠,最终抵达被深埋的雌配子体实现受精[3,35]。玉米和拟南芥等植物作为研究花粉管极性生长的模式材料,通过细胞遗传分析已鉴定发现了许多控制花粉管极性生长的相关基因,雌配子体特异性的蛋白质ZmEA1 和LUREs 被确定为花粉管生长和引导诱物[36-37]。DUAN 等[10]最新研究发现:拟南芥受体激酶(FERONIA) 具有确保精子递送和阻止多重受精的作用,引导花粉管成功“绕开”已经被“占位”的胚珠,找到尚未“占位”的胚珠。苜蓿、小冠花和百脉根授粉4 h 后的胚囊受精率分别为76.40%、88.97% 和89.81%,表明异花授粉状态下,进入雌蕊中的花粉管数量满足胚珠的受精需要,其结实率低下并非由于花粉限制。
豆科牧草具有自交不亲和性,自交衰退明显,提高种子产量主要是通过杂交育种手段来改善。紫花苜蓿、小冠花和百脉根胚珠败育率极高,分别为87.67%、78.00%和81.33%,且荚内胚珠败育位置也具有随机性(表3)。由此可见,胚珠败育减少了每荚种子数,大大降低了其种子产量。通常导致植物选择性败育的原因主要有资源限制和遗传因子2 个方面,这种选择性败育常常可见于豆科植物中,豆科植物产生过多的花和胚珠是其选择高质量后代的一种进化机制[38]。从资源限制角度分析,雌雄同株植物通过雌蕊选择性败育,把资源转移到雄性器官的发育上,形成形态较大、色泽艳丽的花冠,增强对传粉者的吸引,并产生较丰富的花粉粒等,这些报酬可以增加昆虫的拜访频率,因而在雌雄同株上不存在花粉限制[39]。有研究发现:苜蓿的胚珠不育性与珠心胼胝质的沉积密切相关,但不育性具有较大的遗传变异,胼胝质沉积的随机性决定了种子在荚内败育的随机性[40-41]。从遗传因子方面来看,受精后胚珠败育是3 种豆科牧草结实率低下的主要原因,而胼胝质的沉积确实在一定程度上影响了苜蓿胚珠的育性。
4 结论
3 种豆科牧草苜蓿、小冠花和百脉根的柱头在小花还没有完全开放时就具有一定的可授性,当花药完全开裂时,柱头的可授性最强,为昆虫授粉最佳时期。3 种豆科牧草来自异花授粉的花粉与柱头的水合反应、花粉管萌发时间在1~2 h,其中,小冠花和百脉根花柱比苜蓿长,其花粉管生长速度要比苜蓿快,小冠花和百脉根花粉管突破引导组织仅需要4 h 而苜蓿需要8 h。3 种豆科牧草属空心型花柱类型,花柱的中央均有数条纵行的花柱道,花柱沟道表面高度腺性的细胞产生的粘性分泌物可为花粉管的生长和穿行提供营养。苜蓿、小冠花和百脉根授粉4 h 后的胚囊受精率分别为76.40%、88.97%和89.81%,表明异花授粉状态下,进入雌蕊中的花粉管数量满足胚珠的受精需要,其结实率低下并非由于花粉限制。因此,3 种豆科牧草胚珠败育并非由于受精前的花粉萌发和花粉管引导限制,而是和受精后的胚胎发育受阻有关。
猜你喜欢
生物技术通报(2023年2期)2023-03-07
蔬菜(2022年7期)2022-07-29
山东农业科学(2021年8期)2021-09-12
山东农业大学学报(自然科学版)(2020年2期)2020-05-21
草业科学(2019年11期)2019-12-12
数学小灵通(1-2年级)(2018年12期)2018-12-24
海南师范大学学报(自然科学版)(2017年4期)2018-01-22
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
新疆大学学报(自然科学版)(中英文)(2014年2期)2014-11-06
知识窗(2014年4期)2014-04-30
