
低磷胁迫对赤皮青冈幼苗叶片生理指标的影响
2021-03-04 01:33欧阳泽怡陈雯彬欧阳硕龙李志辉何友军颜立红蒋俐媛田晓明向光锋
中南林业科技大学学报 2021年1期
欧阳泽怡,陈雯彬,欧阳硕龙,李志辉,何友军,颜立红,蒋俐媛,田晓明,向光锋
(1.湖南省森林植物园,湖南 长沙 410004;2.中南林业科技大学 林学院,湖南 长沙 410004;3.湖南省林业调查规划设计院,湖南 长沙 410000;4.湖南省林业科学院,湖南 长沙 410004)
对植物来说,磷是贯穿体内核酸与细胞膜脂质的合成、各种酶的活化、信息传递等重要过程的参与者[1-3]。在缺磷的逆境中,植物在其形态、生理生化等方面都会发生极显著变化,进而影响到植物的生长和繁育,但植物也会主动发挥自身能力去适应胁迫逆境[4-5]。
在低磷逆境中,可溶性蛋白对植物的渗透调节能力存在巨大的影响,其含量可以在一定程度上反映植物抗逆性。细胞膜脂质受到活性氧自由基的氧化伤害产生丙二醛(MDA),所以其含量可以反映出细胞膜脂质过氧化的程度和自我修复能力,可体现植物抗逆性能力[6-7]。超氧化物歧化酶(SOD)、过氧化物酶(POD)是保护酶,它们有能力消除过多的活性氧,降低低磷逆境对植物的蛋白质、核酸、膜系统等的破坏程度,所以保护酶的活性变化可以反映出植物的抗逆性能力[8-10]。酸性磷酸酶(APA)活性的提高被认为可加快植物体内磷利用速度[14-16]。所以很多研究者将前述5个指标作为植物对逆境反应机理的重要指标[11-13]。
赤皮青冈Cyclobalanopsis gilva,又名红椆[17],生长在我国南方多个省份,是珍贵的硬木乡土树种;该树种成片天然林稀少[18],需进行科学繁育扩大人工林规模以满足市场需求。我国南方土壤类型大多数为典型缺磷红土壤[19-22],赤皮青冈这一树种在推广栽培过程中必定面临缺磷,因此研究赤皮青冈幼苗对低磷环境的耐受性很有必要。本研究选取赤皮青冈实生幼苗,进行室内控制条件下的低磷胁迫试验,分析低磷胁迫对赤皮青冈幼苗叶片生理指标的影响,为实现高效低耗、低污染的人工林可持续经营发展目标提供参考。
1 材料与方法
1.1 试验材料
以湖南省靖州侗族自治县的赤皮青冈1年生、2年生幼苗为试材。1年生苗高为(15±0.5) cm,地径为(1.9±0.1) cm;2年生苗高为(37±0.2) cm,地径为(3.5±0.2) cm。种源地位于109°36′E,26°16′N,该地降水较充裕,夏日少炎热,冬季少酷寒,早晚温度差别明显[23]。
1.2 试验地概况
试验地点设在天际岭国家森林公园内,该地位于长沙市,一年里超过75%的天数是无霜期。春秋季很短暂,春季气温起伏大,夏季特别炎热而漫长,冬季结冰少但体感较冷。年平均温度为17.2℃[13]。
1.3 试验处理
水培试验于2017年5—9月在湖南省天际岭国家森林公园的玻璃温室进行,营养液按照霍格兰配方[24]配置。5月初在实验苗圃内对赤皮青冈进行移植,培养1 周后于2017年5月11日进行磷胁迫处理,低磷胁迫试验设置8 个磷素水平:0,0.001,0.005,0.01,0.05,0.1,0.5,1 mmol·L-1(CK)。用KH2PO4配置成不同水平浓度,同时在改变浓度时用KCl 来调节营养液中K+浓度平衡,其他营养元素浓度保持相同。每个培养穴内分别放置1 株赤皮青冈幼苗,每个苗龄幼苗每个磷素水平设置3盆,重复3 次,共144 株。在胁迫试验过程中保证24 h 不间断通气,及时调整营养液pH 值至5.8,具体做法是使用0.1 mol·L-1氯化氢和0.1 mol·L-1氢氧化钠。为确保营养足够,每3 d 要倒掉使用过的营养液,注入新配置的营养液,每次测定相关指标间隔30 d。早上8:00 前用枝剪采取1年生、2年生赤皮青冈幼苗新鲜叶片,用冰袋保存,迅速带回实验室,测定前拭净叶片。
1.4 测定方法
1.4.1 赤皮青冈幼苗叶片可溶性蛋白含量的测定
参照考马斯亮蓝G-250 法(Bradford 法)[25]。
可溶性蛋白含量计算公式:可溶性蛋白含量(mg/g)=C×VT/(VS×WF×1 000)。公式中,C为标准曲线中查得的蛋白含量值(μg/mL);VT为提取液总体积(mL);WF为样品鲜质量(g);VS为测定时的加样量(mL)。
1.4.2 赤皮青冈幼苗叶片丙二醛(MDA)含量的测定
样品丙二醛的提取与测定参照陈隆升的方法[26]。
丙二醛含量计算公式:丙二醛浓度(μmol/mL)=[6.45(A532-A600)-0.56A450]×V/W。式中:A532、A600、A450分别代表样品在532、600 和450 nm 波长下的吸光值,V代表提取液体积(L),W代表样品鲜质量。
1.4.3 赤皮青冈幼苗叶片过氧化物酶(POD)活性的测定
参照考愈创木酚法[27]。
1.4.4 赤皮青冈幼苗叶片超氧化物歧化酶(SOD)活性的测定
参照考氮蓝四唑(NBT)法[28]。
1.4.5 赤皮青冈幼苗叶片酸性磷酸酶(APA)活性的测定
参照黄学林[29]的方法。
1.5 数据处理和方法
运用Microsoft Excel 2003 软件进行数据统计和作图,采用SPSS 20.0 软件处理数据,运用单因素方差分析方法(ANOVA)比较各组间试验数据,运用多因素方差分析方法比较1~2年生植株间、不同低磷胁迫处理和不同月份间指标的差异。
2 结果与分析
2.1 低磷胁迫对赤皮青冈幼苗叶片可溶性蛋白含量的影响
由表1可知,低磷胁迫下不同苗龄、不同月份、不同处理的赤皮青冈幼苗可溶性蛋白含量存在极显著差异(P<0.01)。从图1~2 可以看出,在低磷胁迫试验中,1年生、2年生赤皮青冈幼苗叶片可溶性蛋白含量均低于对照,且随胁迫程度加强和时间的延长,可溶性蛋白的含量逐渐减少。总体而言,2年生赤皮青冈幼苗叶片可溶性蛋白含量均高于1年生赤皮青冈幼苗。

表1 不同苗龄赤皮青冈幼苗叶片不同取样月份和不同处理间方差分析†Table 1 Analysis of variance between age.dispose and sampling time of Cyclobalanopsis gilva seedlings leaves
低磷胁迫下,赤皮青冈幼苗可溶性蛋白的含量随着磷元素浓度降低,极显著减少。重度低磷胁迫(0 mmol·L-1) 120 d 时下降幅度达到最大,与对照相比,1年生、2年生赤皮青冈幼苗的可溶性蛋白含量分别下降了76.88%、61.38%,并且存在极显著差异(P<0.01)。

续表1Continuation of table 1

图1 低磷胁迫下1年生赤皮青冈幼苗叶片可溶性蛋白含量Fig.1 The soluble protein content in leaves of Cyclobalanopsis gilva seedlings from annual under low phosphorus stress
2.2 低磷胁迫对赤皮青冈幼苗叶片丙二醛含量的影响

图2 低磷胁迫下2年生赤皮青冈幼苗叶片可溶性蛋白含量Fig.2 The soluble protein content in leaves of Cyclobalanopsis gilva seedlings from biennial under low phosphorus stress
由表1可见,低磷胁迫下不同苗龄赤皮青冈幼苗MDA 含量存在极显著差异;1年生、2年生赤皮青冈幼苗不同月份、不同处理之间MDA 含量均存在极显著差异;从图3~4 可以看出,在低磷胁迫试验中,1年生、2年生赤皮青冈幼苗叶片中的MDA 含量均高于对照,且随试验的进行而持续增加。总体而言,2年生赤皮青冈幼苗叶片MDA 含量均低于1年生赤皮青冈幼苗。关于丙二醛(MDA)含量,它是植物在经受逆境考验情况下产生的,这一逆境也包括衰老,MDA 含量变化是低浓度磷元素胁迫下的直接表现[30]。在实验所得数据中,可以从图3~4 直观看出,随着胁迫时间的推移,受试植株的MDA 含量持续升高,说明低磷胁迫对植株的损害是持续的,特别是对1年生赤皮青冈幼苗的影响远大于2年生植株。
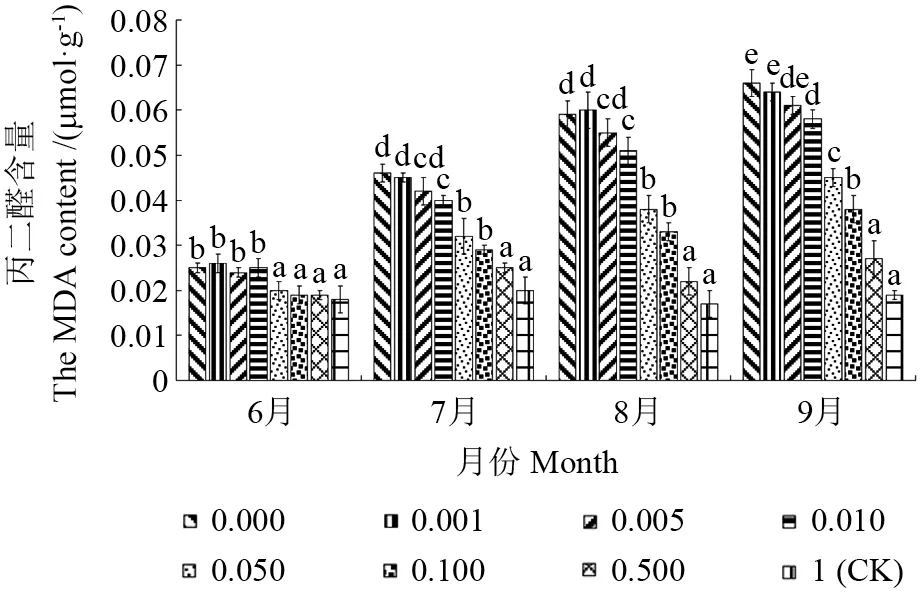
图3 低磷胁迫下1年生赤皮青冈幼苗叶片丙二醛含量Fig.3 The MDA content in leaves of Cyclobalanopsis gilva seedlings from annual under low phosphorus stress
2.3 低磷胁迫对赤皮青冈幼苗叶片过氧化物酶活性的影响

图4 低磷胁迫下2年生赤皮青冈幼苗叶片丙二醛含量Fig.4 The MDA content in leaves of Cyclobalanopsis gilva seedlings from biennial under low phosphorus stress
由表1可见,低磷胁迫下不同苗龄赤皮青冈幼苗POD 活性存在极显著差异;1年生、2年生赤皮青冈幼苗不同月份POD 活性存在极显著差异,不同处理之间POD 活性也存在极显著差异;从图5~6 可以看出,在低磷胁迫试验中,1年生、2年生赤皮青冈幼苗叶片POD 酶活性均高于对照,且随胁迫试验的持续进行,POD 活性会升高。2年生赤皮青冈幼苗不同月份POD 活性存在极显著差异,不同处理之间POD 活性也存在极显著差异;2年生赤皮青冈幼苗叶片POD 活性均高于1年生赤皮青冈幼苗。

图5 低磷胁迫下1年生赤皮青冈幼苗叶片POD 活性含量Fig.5 The POD activity in leaves of Cyclobalanopsis gilva seedlings from annual under low phosphorus stress
植物体内的过氧化物酶活性与其抗逆性呈正相关[31]。特别需要指出的是在赤皮青冈1年生、2年生幼苗对低磷胁迫的耐受浓度达到0.01 mmol·L-1时,过氧化物酶POD 的活性含量出现大幅度上升;培养液磷浓度在1~0.01 mmol·L-1变化范围内,赤皮青冈组织内部的POD 活性变化幅度最为巨大。由此可见,在外界环境磷浓度逐渐减少到0.01 mmol·L-1,赤皮青冈幼苗组织内部的POD 活性快速增强,植株的抗逆性会迅速增强,并趋于稳定。在磷浓度低于0.01 mmol·L-1,过氧化物酶的活性只是随着磷浓度的下降,活性稍有增强,但是增强幅度较小,这也从局部显示了赤皮青冈幼苗对低磷环境的耐受程度。

图6 低磷胁迫下2年生赤皮青冈幼苗叶片过氧化物酶活性含量Fig.6 The POD activity in leaves of Cyclobalanopsis gilva seedlings from biennial under low phosphorus stress
2.4 低磷胁迫对赤皮青冈幼苗叶片超氧化物歧化酶活性的影响
由表1可见,低磷胁迫下不同苗龄赤皮青冈幼苗SOD 活性存在极显著差异;1年生赤皮青冈幼苗不同月份SOD 活性存在极显著差异,不同处理之间SOD 活性也存在极显著差异;从图7~8可知,在低磷胁迫试验中,1年生、2年生赤皮青冈幼苗叶片SOD 活性均高于对照,且随胁迫时间的延长SOD 活性升高。2年生赤皮青冈幼苗不同月份SOD 活性存在极显著差异;2年生赤皮青冈幼苗叶片SOD 活性均高于1年生赤皮青冈幼苗。在低磷胁迫试验中,1年生、2年生赤皮青冈幼苗叶片SOD 活性均高于对照,且随胁迫时间的延长SOD 活性升高。

图7 低磷胁迫下1年生赤皮青冈幼苗叶片超氧化物歧化酶活性含量Fig.7 The SOD activity in leaves of Cyclobalanopsis gilva seedlings from annual under low phosphorus stress
超氧化物歧化酶(SOD)活性的变化趋势与过氧化物歧化酶(POD)活性的变化趋势基本一致,2年生的赤皮青冈幼苗高于1年生的赤皮青冈幼苗,同样在外界磷浓度达到0.01mmol·L-1,SOD 活性基本达到变化的最大值,且随着外界磷浓度降低,SOD 活性变化趋于稳定。结合SOD和POD 这两类植物抗逆性指标进行变化趋势类比,可发现,想要最大化的激活赤皮青冈植物内部的抗逆物质,低磷环境中的磷含量需要低于0.01 mmol·L-1。在通常的研究中,0.01 mmol·L-1的磷浓度培养液已经属于低浓度[3],而且在低磷环境中,赤皮青冈幼苗生长性状表现良好。由此也可以证明赤皮青冈幼苗对低磷环境的耐受性良好。

图8 低磷胁迫下2年生赤皮青冈幼苗叶片超氧化物歧化酶活性含量Fig.8 The SOD activity in leaves of Cyclobalanopsis gilva seedlings from biennial under low phosphorus stress
2.5 低磷胁迫对赤皮青冈幼苗叶片酸性磷酸酶活性的影响
由表1可见,低磷胁迫下不同苗龄赤皮青冈幼苗叶片APA 活性存在极显著差异;1年生赤皮青冈幼苗不同月份叶片APA 活性存在极显著差异,不同处理之间叶片APA 活性也存在极显著差异;从图9~10 可知,在低磷胁迫试验中,1年生、2年生赤皮青冈幼苗叶片APA 活性均高于对照,叶片APA 活性会随时间推移升高。1年生赤皮青冈幼苗6—9月份叶片APA 活性在各梯度P 浓度下与对照组存在极显著差异(P<0.01)。2年生赤皮青冈幼苗不同月份叶片APA 活性存在极显著差异,不同处理之间表现亦然(表1);2年生赤皮青冈幼苗叶片APA 活性均高于1年生赤皮青冈幼苗。

图9 低磷胁迫下1年生赤皮青冈幼苗叶片酸性磷酸酶活性含量Fig.9 The APA activity in leaves of Cyclobalanopsis gilva seedlings from annual under low phosphorus stress

图10 低磷胁迫下2年生赤皮青冈幼苗叶片酸性磷酸酶活性含量Fig.10 The APA activity in leaves of Cyclobalanopsis gilva seedlings from biennial under low phosphorus stress
植株体内的磷元素含量和土壤中的磷元素含量直接影响着植物体内酸性磷酸酶的活性。大量实验证明,植物应答低磷胁迫,体内酸性磷酸酶活性会极显著增加[32]。赤皮青冈幼苗内部的APA 含量在1~0.01 mmol·L-1区间变化最大,以磷浓度0.01 mmol·L-1为界限,随着浓度的降低,APA 含量的变化趋于稳定。从酸性磷酸酶指标数据中,可以发现其变化形式与SOD、POD 活性变化情况基本一致。基于上述情况可以发现,在培养液磷浓度为0.01 mmol·L-1时,在不影响幼苗正常发育的情况下,赤皮青冈幼苗对低磷的耐受性已经达到最大值。
2.6 低磷胁迫下赤皮青冈幼苗叶片主要生理变化的相关性
由表2可以看出,1年生、2年生赤皮青冈幼苗叶片可溶性蛋白含量与MDA 含量呈极显著负相关(P<0.01),1年生、2年生赤皮青冈幼苗叶片MDA 含量与POD、SOD 活性呈极显著正相关(P<0.01)。即磷元素浓度降低后,赤皮青冈幼苗可溶性蛋白含量减少,细胞渗透调节能力下降,细胞膜受到破坏,细胞膜脂质过氧化程度加强,过氧化产物MDA 含量增加;低磷胁迫使POD、SOD活性增强,清除活性氧化自由基能力增强,有利于缓解细胞膜遭受的损伤,降低MDA 含量,但是不能完全清除胁迫后增加的活性氧自由基[33-34]。

表2 低磷胁迫下1年生、2年生赤皮青冈幼苗叶片主要生理变化的相关性分析†Table 2 Partial correlation analysis of the physiological indexes of Cyclobalanopsis gilva seedlings from annual,biennial under low phosphorus stress
3 讨 论
3.1 赤皮青冈幼苗叶片可溶性蛋白含量对低磷胁迫的响应
在低磷逆境下,植物细胞为了增强自身细胞渗透调节能力,通常是通过累积渗透物质来达到目标,而可溶性蛋白就是符合这一需求的物质[35]。
在本次低磷胁迫试验中,1年生、2年生赤皮青冈可溶性蛋白含量随低磷胁迫的程度增大而呈降低的趋势,说明低磷胁迫导致赤皮青冈幼苗可溶性蛋白合成受到抑制,赤皮青冈幼苗细胞渗透调节能力下降。马尾松[32]、小麦[24]、油茶[26]等叶片也会有相似表现,陈隆升等[26]认为这可能是由缺磷蛋白质合成受阻导致。此外,2年生赤皮青冈幼苗可溶性蛋白含量降低幅度明显小于1年生赤皮青冈幼苗,说明在同等的低磷胁迫下,2年生赤皮青冈幼苗细胞渗透调节能力比1年生赤皮青冈幼苗强。
3.2 赤皮青冈幼苗叶片丙二醛含量对低磷胁迫的影响
细胞膜脂质受到活性氧自由基的氧化伤害产生丙二醛,所以丙二醛的含量可以反映出细胞自我修复能力和膜脂质过氧化的程度,也是体现植物抗逆性能力的有效指标[36]。
在本次低磷胁迫试验中,1年生、2年生赤皮青冈幼苗体内丙二醛的含量随低磷胁迫程度增高而呈升高的趋势,表明赤皮青冈细胞膜脂质受到活性氧自由基的氧化伤害随低磷胁迫程度加强而加重。说明低磷胁迫下,赤皮青冈受到磷胁迫影响从而叶片产生更多的自由基,加快细胞膜脂质过氧化的程度,导致叶片内丙二醛含量的增加。大豆[8]、豇豆[9]、甘蔗[10]、水稻[11]等植物叶片在低磷胁迫下亦有类似表现,其中张玉霞等[8]发现,磷低效基因大豆丙二醛含量的变化程度较磷高效基因大豆更大。此外,不同苗龄赤皮青冈幼苗的丙二醛含量变化存在一定的差异,相比而言,2年生赤皮青冈幼苗叶片在各低磷处理下MDA 含量始终明显低于1年生赤皮青冈。说明在同等的低磷胁迫下,2年生赤皮青冈幼苗产生较少的自由基,细胞膜脂质过氧化程度比1年生赤皮青冈幼苗弱。
3.3 赤皮青冈幼苗叶片保护酶活性对低磷胁迫的响应
活性氧含量的提升会诱导植物其他信号,从而控制抗氧化系统,有很多研究者将其作为植物对逆境反应机理的重要指标[8-10]。保护酶活性的提高使赤皮青冈幼苗在一定程度上更加适应低磷胁迫,甘蔗[14]、番茄[30]、马尾松[32]等植物的酶活性在低磷胁迫期间的表现亦是如此,王晶等[30]认为,保护酶的绝对活力与低磷胁迫无关,而变化幅度与低磷胁迫有关。
3.4 赤皮青冈幼苗的POD 和SOD 酶活性随低磷胁迫程度增加而上升,
重度低磷胁迫(0 mmol·L-1)120 d 时达到最大。在低磷胁迫试验中,1年生、2年生赤皮青冈幼苗叶片POD、SOD 的活性均随磷浓度的降低而呈升高的趋势,说明低磷胁迫刺激赤皮青冈幼苗增强保护酶活性,导致清除自由基的能力提高,使得赤皮青冈细胞膜脂质的过氧化程度缓解;且随胁迫试验的持续进行,1年生、2年生赤皮青冈幼苗叶片POD、SOD 的活性的增加幅度越来越小,最后趋于平缓,这可能是赤皮青冈幼苗开始形成一定的耐低磷机制,使抗氧化系统维持一定的平衡状态;而且在低磷胁迫试验中,2年生赤皮青冈幼苗在各低磷胁迫处理下始终保持较高的POD和SOD 酶活性,并在0.1 mmol·L-1的P 浓度处理下达到差异极显著水平(P<0.01)。2年生赤皮青冈幼苗叶片保护酶活性大于1年生赤皮青冈,表明2年生赤皮青冈的活性氧清除系统能力强于1年生赤皮青冈幼苗。
3.5 赤皮青冈幼苗叶片酸性磷酸酶(APA)活性对低磷胁迫的响应
酸性磷酸酶与植物磷营养有着密切的联系,它能够使环境中原有的有机磷发生化学反应后转化为无机磷,植物由此更容易获得磷,所以植物酸性磷酸酶活性的变化是体现植物内磷利用效率的有效指标[14-16,33]。目前已经有很多研究者发现低磷逆境可以提高植物酸性磷酸酶活性,Nanamori等[36]研究发现,在低磷胁迫下,水稻和牧草叶片酸性磷酸酶受到影响且与体内的磷利用效率相关性极显著。如Yun 等[37]研究表明,玉米在低磷胁迫处理14 d 后,正常供磷植株叶片酸性磷酸酶活性极显著低于磷胁迫处理的植株。
在本次试验中,1年生、2年生赤皮青冈幼苗叶片酸性磷酸酶活性随磷胁迫程度增加而增加,说明在磷胁迫下,随磷浓度降低,为满足自身对磷元素的需求,从而提高植物体内酸性磷酸酶活性来提高磷的利用效率,这种提升是赤皮青冈幼苗的APA 活性对低磷胁迫的响应。这种响应非常明显,当磷浓度低于0.5 mmol·L-1时,低磷胁迫极显著提高了赤皮青冈幼苗叶片APA、根系APA 和根系分泌APA 的活性(P<0.01)。比较不同苗龄赤皮青冈幼苗在各梯度低磷处理下磷酸酶活性的变化后可知,2年生赤皮青冈幼苗增加幅度普遍高于1年生,即2年生赤皮青冈幼苗的磷利用效率高于1年生赤皮青冈,表明2年生赤皮青冈幼苗磷适应低磷逆境能力比1年生赤皮青冈幼苗强。
3.6 低磷胁迫下赤皮青冈幼苗叶片内MDA 含量、POD 活性和SOD 活性的相关性
本实验结果表明,随着低磷胁迫程度的进一步加深和试验的持续进行,赤皮青冈幼苗叶片内MDA 含量、POD 活性和SOD 活性均持续上升,并在1~0.01 mmol·L-1这段浓度变化的区间,上述3 个指标变化情况最为明显。说明低磷胁迫一方面刺激赤皮青冈叶片产生更多的MDA,进而引起赤皮青冈叶片细胞活性氧自由基代谢失衡;另一方面由于低磷胁迫后赤皮青冈活性氧自由基积聚刺激了保护酶系统的提升,能清除自由基,削弱了膜脂过氧化,进而降低赤皮青冈体内MDA 含量。由此可以看出这两方面形成一种动态平衡[24-31,38-39],是赤皮青冈耐低磷胁迫的主要抗性机制。但在磷浓度低于0.01 mmol·L-1之后,上述3 个指标变化情况基本趋于稳定,则说明赤皮青冈幼苗在不影响正常生长的情况下,对于低磷的耐受限度为0.01 mmol·L-1,外界环境的有效磷低于该含量就可能会造成幼苗发育不正常或者死亡,在本实验中低于0.01 mmol·L-1浓度培养液中的幼苗在试验完成后逐渐枯萎死亡,死亡率达到45%。
本研究的局限在于只选取了1年生、2年生赤皮青冈幼苗叶片为实验材料研究赤皮青冈幼苗生理指标在低磷胁迫下的变化,并不能全面反映不同苗龄的赤皮青冈幼苗对低磷环境的适应情况。在以后的研究中,应选取更多不同苗龄的赤皮青冈幼苗,获得不同低磷胁迫处理对不同苗龄赤皮青冈幼苗影响的实验数据,为阐明不同苗龄赤皮青冈幼苗适应低磷胁迫的机制提供更详细的数据证明。
4 结 论
本研究发现2年生赤皮青冈幼苗MDA 含量上升幅度小于1年生赤皮青冈幼苗,保护酶SOD、POD 活性升高幅度大于1年生赤皮青冈幼苗。相同低磷胁迫程度下,1年生赤皮青冈幼苗MDA 含量均高于2年生赤皮青冈幼苗,2年生赤皮青冈幼苗保护酶SOD、POD活性均高于1年生赤皮青冈幼苗。以上表明相同低磷胁迫程度下,1年生赤皮青冈幼苗受到低磷胁迫的危害更大;2年生赤皮青冈幼苗的活性氧清除能力优于1年生赤皮青冈幼苗。
为适应及抵抗外界的低磷逆境,赤皮青冈幼苗叶片蛋白质含量、MDA 含量、保护酶活性和APA 活性都经历了极显著改变,这证明赤皮青冈幼苗可以通过自身激素及酶等物质的综合调节来减少低磷环境对植物的伤害,并通过自身酶活性的增强来止损和加强外界环境磷元素的吸收以适应低磷逆境,最终达到低磷耐受极限0.01 mmol·L-1。长期生存在0.01 mmol·L-1的低磷环境下,植株无法正常生长发育。
结合本研究和实际生产,因2年生赤皮青冈幼苗耐低磷表现优于1年生赤皮青冈幼苗,所以在较为缺磷的林地进行造林作业时要尽量选取苗龄较大的赤皮青冈幼苗。赤皮青冈这一树种对低磷环境的耐受性较高,耐受极限完全低于湖南地区酸性红壤中的磷含量。在湖南广布的酸性红壤地区都能栽培赤皮青冈的幼苗,但是在培育赤皮青冈高效用材林和使赤皮青冈达到最优生长情况的前提下,并不建议在酸性红壤低磷环境下栽培赤皮青冈。
猜你喜欢
昆明医科大学学报(2021年8期)2021-08-13
报刊荟萃(上)(2017年7期)2017-07-16
魅力中国(2016年29期)2017-05-27
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年6期)2016-07-25
农家顾问(2016年7期)2016-05-14
医学研究杂志(2015年9期)2015-07-01
食品工业科技(2014年23期)2014-03-11
食品与生活(2014年2期)2014-03-04
