
蝴蝶兰(Phalaenopsis spp.)与钻喙兰(Rhynchostylis retusa)远缘杂交种子无菌播种及后代表型分析
2021-03-19 05:56陈和明吕复兵肖文芳
中国农业大学学报 2021年3期
陈和明 吕复兵 李 佐 肖文芳
(广东省农业科学院 环境园艺研究所/广东省园林花卉种质创新综合利用重点实验室,广州 510640)
蝴蝶兰(Phalaenopsisspp.)因其花形奇特、花色艳丽、色泽丰富、花序整齐、花期长,素有“洋兰皇后”的美称。蝴蝶兰是目前世界上销售量最大的兰花品种,全球年消费量已超过2.8 亿株[1-2]。近年来,通过杂交育种技术已培育出许多蝴蝶兰的优良新品种,支撑了蝴蝶兰产业的快速发展,但多数蝴蝶兰没有香味、缺乏蓝色花且抗性较差[2-3]。因此,将蝴蝶兰近缘属野生种质资源的优良基因通过远缘杂交的方式导入蝴蝶兰中,丰富蝴蝶兰的基因库和提高蝴蝶兰的品质具有重要的理论和实践意义。
钻喙兰属(Rhynchostylis)植物有较高观赏价值,其花色淡雅且多数具芳香,花序饱满,花朵数多达上百朵,形似一条毛茸茸的狐狸尾巴,俗称“狐尾兰”,植株抗性较强[4]。已发现的钻喙兰属约6 种,分布于热带亚洲,我国有2 种,产于南方热带地区,分别为钻喙兰(R.retusa)和海南钻喙兰(R.gigantea)[5]。蝴蝶兰与钻喙兰远缘杂交组合最早于1965年在英国皇家园艺学会(RHS)上登录,此杂种以钻喙兰为母本,朵丽兰为父本,新杂交属名为Rhynchonopsis,杂种名为Winona Jordan,目前共获得41 个杂交组合在RHS上正式登录[6],其中:以蝴蝶兰为母本、钻喙兰为父本的杂交组合40 个,以钻喙兰为母本、蝴蝶兰为父本的杂交组合1 个。
蝴蝶兰远缘杂交研究主要有:张志胜等[7]研究了中国兰花与蝴蝶兰等气生兰的远缘杂交,但与蝴蝶兰杂交成功率极低且未获得杂种后代;李枝林等[8]报道了蝴蝶兰与兰属的碧玉兰进行属间杂交不成功的情况;Kim等[9]研究了风兰(Neofinetiafalcata)与蝴蝶兰之间的远缘杂交并获得杂种后代;陈和明等[10]对蝴蝶兰与火焰兰(Renanthera)进行正反远缘杂交,共计完成杂交组合48 个,获得441 株杂交后代。然而,蝴蝶兰与钻喙兰(R.retusa)远缘杂交研究尚未见报道。因此,本研究拟以18 份蝴蝶兰与钻喙兰(R.retusa)为研究对象,主要利用钻喙兰花数多、有气味且抗性较强的优良性状,进行蝴蝶兰与钻喙兰之间的远缘杂交试验,观察和分析种子的无菌播种和后代的表型性状,旨在选育出抗性较强、有香味或多花的远缘杂交新品种。
1 材料与方法
1.1 试验材料
以聚宝红玫瑰(P. Jiuhbao Red Rose)、光芒四射(P. Formosa Sunrise)、翔凤(P. Jiuhbao Red Rose‘Xiangfeng’)、斯卡利特(P. Scarlet in Snow)、沙拉金(P. Brother Sara Gold)、富乐夕阳(P. Fuller’s Sunset)、小樱桃(P. Jiaho Cherry)、珍珠红(P. Sunrise Firebird)、红冠(P. Frigdaas Spring Rose ‘Hongguan’)、宫粉(P. Jiuhbao Red Rose‘Gongfen’)、童真(P. Brother Sara Gold ‘Tongzhen’)、魅力(P. KV Charmer)、小飞象(P. Sogo Vivien)、日本满天红(P. Queen Beer ‘Japan’)、满山红(P. Chin-Lih Rose ‘Mansanhon’)、荧光蝴蝶兰(P.violacea)、版纳蝴蝶兰(P.mannii)和朵丽兰(P.pulcherrima)等18份蝴蝶兰,以及钻喙兰(R.retusa)(图1,图2)作为远缘杂交亲本(表1)。其中,蝴蝶兰自然花期3—6月,钻喙兰自然花期5—6月,均种植于广东省名优花卉种质资源库兰花圃内。

图1 钻喙兰开花植株

图2 钻喙兰花
1.2 杂交授粉
2013—2015年的5—6月,对蝴蝶兰和钻喙兰互为亲本进行正反杂交授粉各5朵,杂交方法参照Kim等[9]和陈和明等[10]的方法。在开花期选择晴天10:00—12:00进行杂交,挑选开放3~5 d的花朵,用消毒过的镊子取下蝴蝶兰新鲜的花粉块,然后用镊子轻轻地把花粉块置入钻喙兰的柱头里,同样取下钻喙兰新鲜的花粉块并置入蝴蝶兰的柱头里进行正反杂交,并对杂交的花朵挂上标签并注明杂交组合及授粉时间,同时套袋以防昆虫授粉,7 d后拆除套袋,待果荚成熟后进行无菌播种。
1.3 种子无菌播种
种子无菌萌发:将果荚流水冲洗30 min并用70%乙醇表面灭菌1 min,在超净工作台上用0.1%的HgCl2处理10 min,再用无菌水冲洗3~5次,用无菌滤纸吸干残余水分后,用解剖刀剖开果荚并观察里面的种子情况,然后用无菌镊子将种子轻轻夹到以下3种诱导培养基上培养。诱导培养基分别为M1:花宝1号3 g/L+BA 2.0 mg/L+NAA 0.1 mg/L;M2:花宝1号 3 g/L+BA 2.0 mg/L+NAA 1.0 mg/L;M3:花宝1号3 g/L+BA 2.0 mg/L+NAA 2.0 mg/L,以上培养基分别添加蔗糖30.0 g/L+椰汁100.0 ml/L+琼脂7.0 g/L,每个处理接种3瓶。以胚突破种皮形成白色原球茎为标准统计萌发率。培养条件:培养温度25~28 ℃,前期暗培养或有散射光即可,后期光照强度1 000~1 500 lx,光照时间12 h/d。
增殖与分化培养:将萌发后呈嫩绿色的原球茎接种到增殖培养基上,增殖培养基分别为L1:花宝1号3 g/L+BA 1.0 mg/L+Ad 1.0 mg/L;L2:花宝1号3 g/L+BA 2.0 mg/L+Ad 2.0 mg/L;L3:花宝1号3 g/L+BA 3.0 mg/L+Ad 3.0 mg/L。每个处理接种3瓶,每瓶10个原球茎,培养100 d后在超净工作台统计增殖的芽数量,并转接至分化培养基花宝1号3 g/L+BA 0.1 mg/L+NAA 0.01 mg/L 上进行分化培养。以上培养基分别添加蔗糖30.0 g/L+椰汁100.0 ml/L+琼脂7.0 g/L。培养条件:光照强度1 500~2 000 lx,光照12 h/d,培养温度25~28 ℃。
壮苗和生根培养:将已分化的苗转接到壮苗生根培养基上,生根培养基分别为T1:花宝1号3 g/L+NAA 0.1 mg/L;T2:花宝1号3 g/L+NAA 0.5 mg/L;T3:花宝1号3 g/L+NAA 1.0 mg/L,以上培养基分别添加蔗糖30.0 g/L+椰汁100.0 ml/L+琼脂7.0 g/L。每个处理接种5 瓶,每瓶10 个芽苗,培养100 d后统计小苗的叶片数、叶片长度、叶片宽和根数及根长。培养条件:光照强度1 500~2 000 lx,光照12 h/d,培养温度25~28 ℃。
1.4 杂种后代调查
2017年6月杂种F1代实生苗出瓶移栽,经过小苗、中苗和大苗的栽培管理后,杂种F1实生苗在2020年3—5月陆续开花,开花期参照Vo等[11]以及陈和明等[10]的方法调查杂种F1代和亲本植株的开花阶段的相关性状,调查开花率,以及6个数量性状:叶长、叶宽、花梗长、花梗粗、单枝花朵数、花径;假质量性状12个:叶片形状、叶色、花梗颜色、花梗姿势、萼片形状、花瓣形状、萼片主色、花瓣主色、唇瓣主色、花香、移植至成苗始花、开花期。
2 结果与分析
2.1 蝴蝶兰和钻喙兰正反远缘杂交的座果率差异
不同品种的蝴蝶兰与钻喙兰远缘杂交座果率差异明显(表1)。由表1可见:在36个正反远缘杂交中,以蝴蝶兰为母本、钻喙兰为父本的杂交组合18个,有12个杂交组合座果,其组合座果率在20%~80%均有;以钻喙兰为母本、蝴蝶兰为父本的杂交组合18个,但所有的组合均未座果,座果率为0%且授粉后5~10 d全部蔫萎凋谢;以蝴蝶兰为母本,钻喙兰为父本座果的12个杂交组合中,果荚发育时间超过100 d的仅有1个,为PR01(蝴蝶兰‘聚宝红玫瑰’×钻喙兰),果荚发育时间60~90 d,共有9个,分别为PR05、PR09、PR13、PR15、PR23、PR27、PR29、PR31和PR35,而30~60 d仅有2个,为PR19和PR21(表1)。
2.2 不同诱导培养基对远缘杂交种子萌发的影响
由表1可知,12个成功座果的杂交组合均是以蝴蝶兰为母本、钻喙兰为父本,种子经无菌播种、在不同诱导培养基上培养后,只有PR01(蝴蝶兰‘聚宝红玫瑰’×钻喙兰)的种子萌发(图3(a)和(b)),且在M1、M2、M3的3 种培养基上均能萌发,萌发率达到100%,但其原球茎形成时间和转绿时间有所不同,随着生长调节剂浓度的升高,所需时间变短;PR05、PR09、PR13、PR15、PR19、PR21、PR23、PR27、PR29、PR31及PR35杂交组合的种子在不同诱导培养基上培养2 年均未萌发(图3(c)和(d))。结合表1可知,PR01的果荚发育时间达到110 d,而PR05、PR09、PR13、PR15、PR19、PR21、PR23、PR27、PR29、PR31和PR35的果荚发育时间在 30~90 d,这些果荚与PR01相比,可能因为发育时间不足、种胚未能形成,导致种子不能萌发。
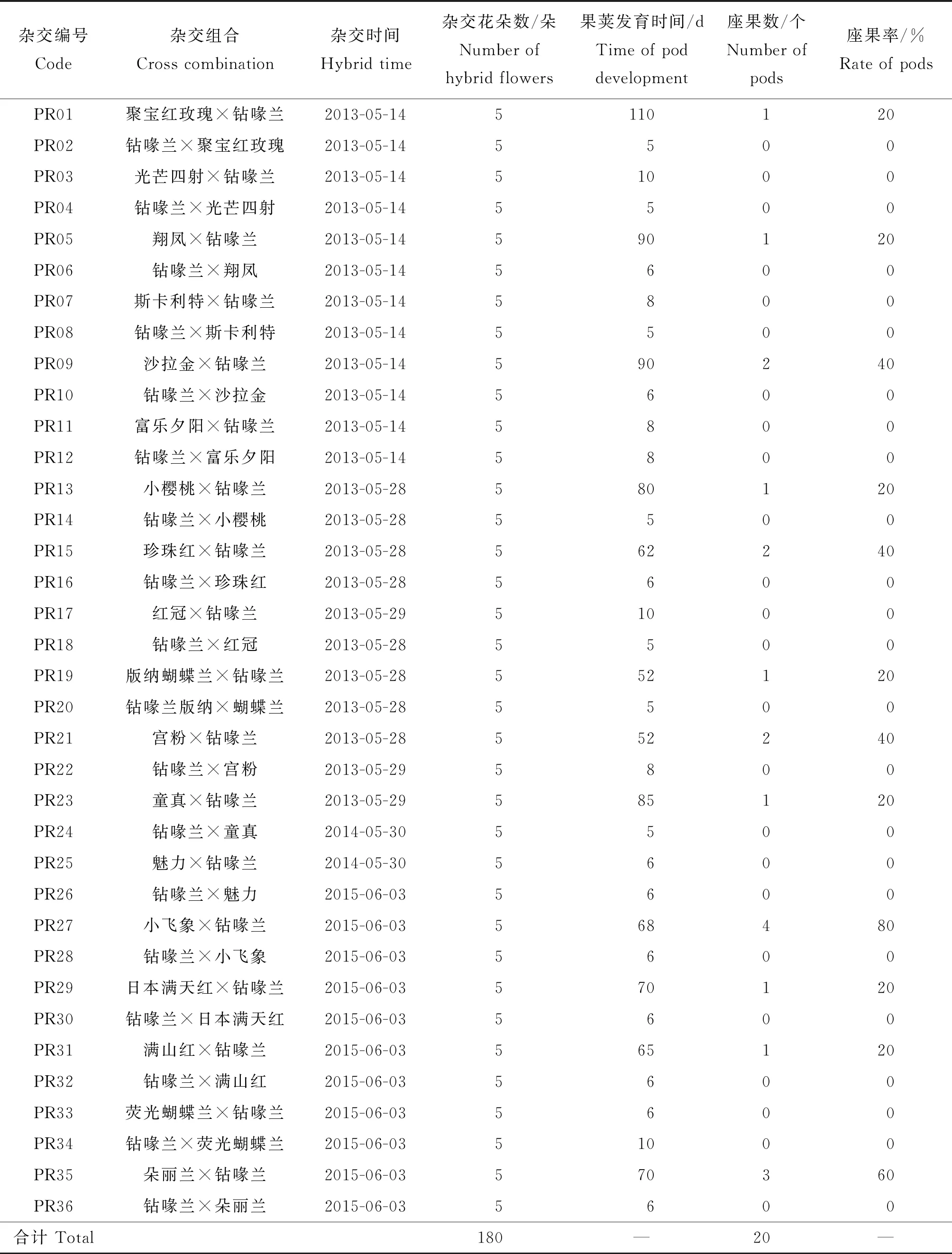
表1 不同品种蝴蝶兰与钻喙兰正反杂交及座果率
2.3 远缘杂交后代(PR01)的增殖
由于仅PR01(蝴蝶兰‘聚宝红玫瑰’×钻喙兰)杂交后代的种子萌发,因此,只对PR01的原球茎进行了增殖研究,不同激素水平对PR01原球茎增殖的影响结果见表2。由表2可知,在接种的各处理的30 个原球茎中,L1处理的增殖系数为2.0,而L2和L3处理的增殖系数分别达到3.1和3.3,表明处理L2和L3,即BA 2.0~3.0 mg/L、Ad 2.0~3.0 mg/L 适合于PR01原球茎的增殖。从图3(a)、(b)和(e)来看,图3(a)为杂交组合PR01的果荚,图3(b)为PR01种子萌发长成原球茎,图3(e)为PR01原球茎的增殖情况;而图3(c)和(d)为果荚发育时间为30~90 d的果荚,以及无菌播种后不萌发的情况。

表2 不同激素水平对PR01原球茎增殖的影响
2.4 远缘杂交苗(PR01)的壮苗生根
将PR01分化的幼苗(图3(f))转接到壮苗生根培养基上,培养100 d后,结果见表3。由表3结果可以看出,3个处理T1、T2、T3,在叶片数、叶片长、叶片宽上无显著差异,但根数和根长上具有明显差异,表现在根数量T1、T2与T3之时存在显著差异,而T1和T2之间差异不明显,根长T2、T3与T1差异明显,但T2、T3之间不存在差异。因此,T2有利于PR01无菌苗的壮苗生根(图3(g)和(h)。

(a)发育时间充足的果荚(杂交组合PR01);(b)PR01种子长成原球茎;(c)发育时间不充足的果荚;(d)种子无菌播种后不萌发;(e)PR01原球茎的增殖;(f)PR01分化成苗;(g)PR01壮苗生根;(h)PR01移栽种植。
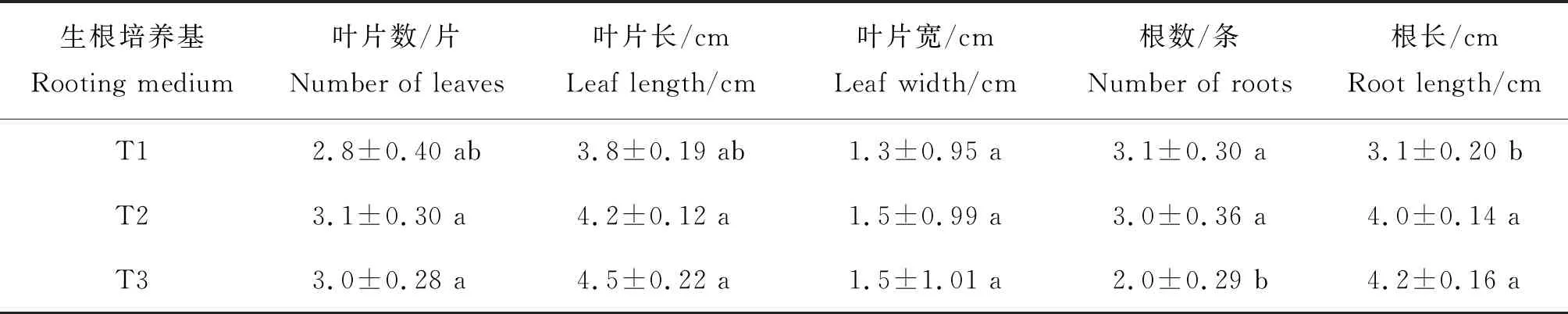
表3 不同培养基对PR01壮苗生根的影响
2.5 远缘杂交后代F1(PR01)的数量性状分析
2017年6月杂种F1代(PR01)实生苗出瓶移栽,经过3年的生长,于2020年3月始花,其后代开花率及其主要数量性状见表4。此杂种F1群体植株数141株,但开花株数仅有22株,开花率为15.6%;而母本蝴蝶兰‘聚宝红玫瑰’和父本钻喙兰的开花率均为100%,表明经过远缘杂交后,杂种F1的开花不一致,出现了性状分离(表4);从叶长和叶宽的性状来看:母本蝴蝶兰‘聚宝红玫瑰’和父本钻喙兰的叶长均值分别为17.3、36.0 cm,叶宽分别为6.8、2.5 cm,子代F1叶长、叶宽分别为20.1、5.4 cm,介于双亲之间(表4);母本蝴蝶兰‘聚宝红玫瑰’和父本钻喙兰的花梗长分别为71.7、43.8 cm,花梗粗分别为6.3、3.2 mm(表4),单枝花朵数分别为9、54朵,花径分别为9.7、1.7 cm,子代F1的花梗长、花梗粗、单枝花朵数和花径分别为37.4 cm、3.8 mm、9.2朵和5.3 cm,与双亲比较除了花梗长比双亲小,花梗粗、单枝花朵数和花径均介于双亲之间,但叶长、叶宽、单枝花朵数和花径趋向于母本蝴蝶兰‘聚宝红玫瑰’(表4)。
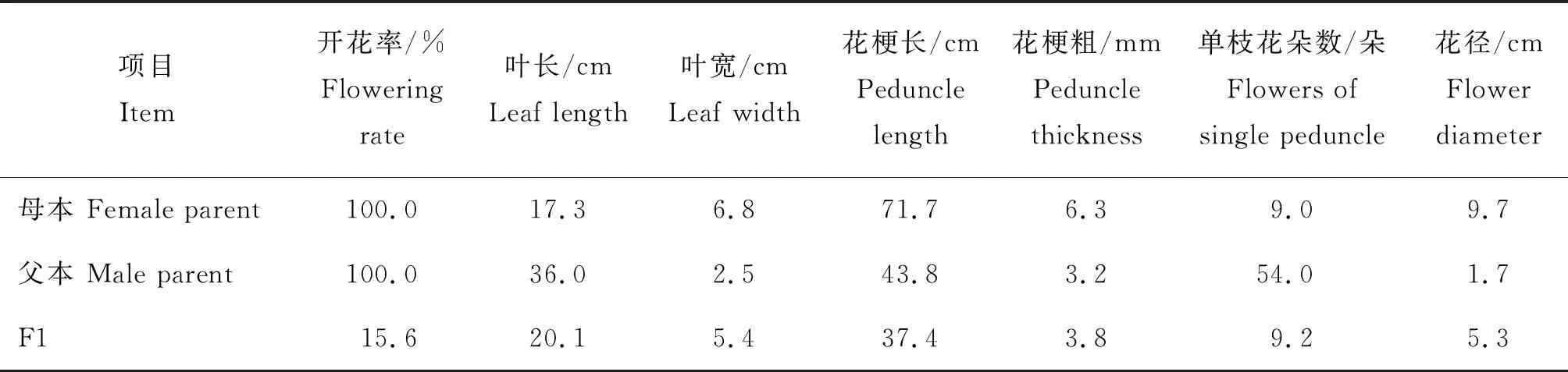
表4 PR01亲本及F1的开花率及其主要数量性状
2.6 远缘杂交后代F1(PR01)的假质量性状分析
从表5杂种F1和亲本主要假质量性状来看,子代F1叶片形状均为窄倒卵圆形(图4(c)),介于母本倒卵圆形(图4(a))和父本披针形(图4(b))之间;子代F1叶色、花梗颜色均为绿色,以及花梗姿势表现为直立,均与母本一致;子代F1萼片形状、花瓣形状均为卵圆形,介于双亲之间,但无论是叶片、花梗还是萼片、花瓣均趋向于母本;从花的主色分析,母本萼片、花瓣的主色均为深紫(N57-A)(图4(e)),父本萼片、花瓣的主色均为白色(NN155-D)(图4(e)),而子代F1的萼片、花瓣主色表现为从深紫(N57-A)、紫红(N57-B)、粉红(N57-C)、浅粉(N57-D)和淡粉(62-B)均有(图4(d)和(e)),介于母本萼片、花瓣主色深紫(N57-A),父本钻喙兰萼片、花瓣主色白色(NN155-D)之间,而母本唇瓣主色为深紫(58-A)(图4(e)),父本唇瓣主色为淡紫(58-D)(图4(e)),子代F1唇瓣主色分别表现为深紫(58-A)、紫红(58-B)和浅紫(58-C)(图4(d)和(e)),因此,无论从萼片、花瓣主色,还是唇瓣主色,子代F1的主色均介于双亲之间,表明子代遗传了亲本的花色。同时,子代F1均无香味,而母本无香味,但父本钻喙兰有气味,但不香(表5)。植株从移植至成熟大苗始花,母本蝴蝶兰‘聚宝红玫瑰’需2年,父本钻喙兰需4年,而子代F1需3年,表明子代从移植至成熟大苗的始花时间介于双亲之间,但开花期子代与母本相近,表现为3—5月(表5)。
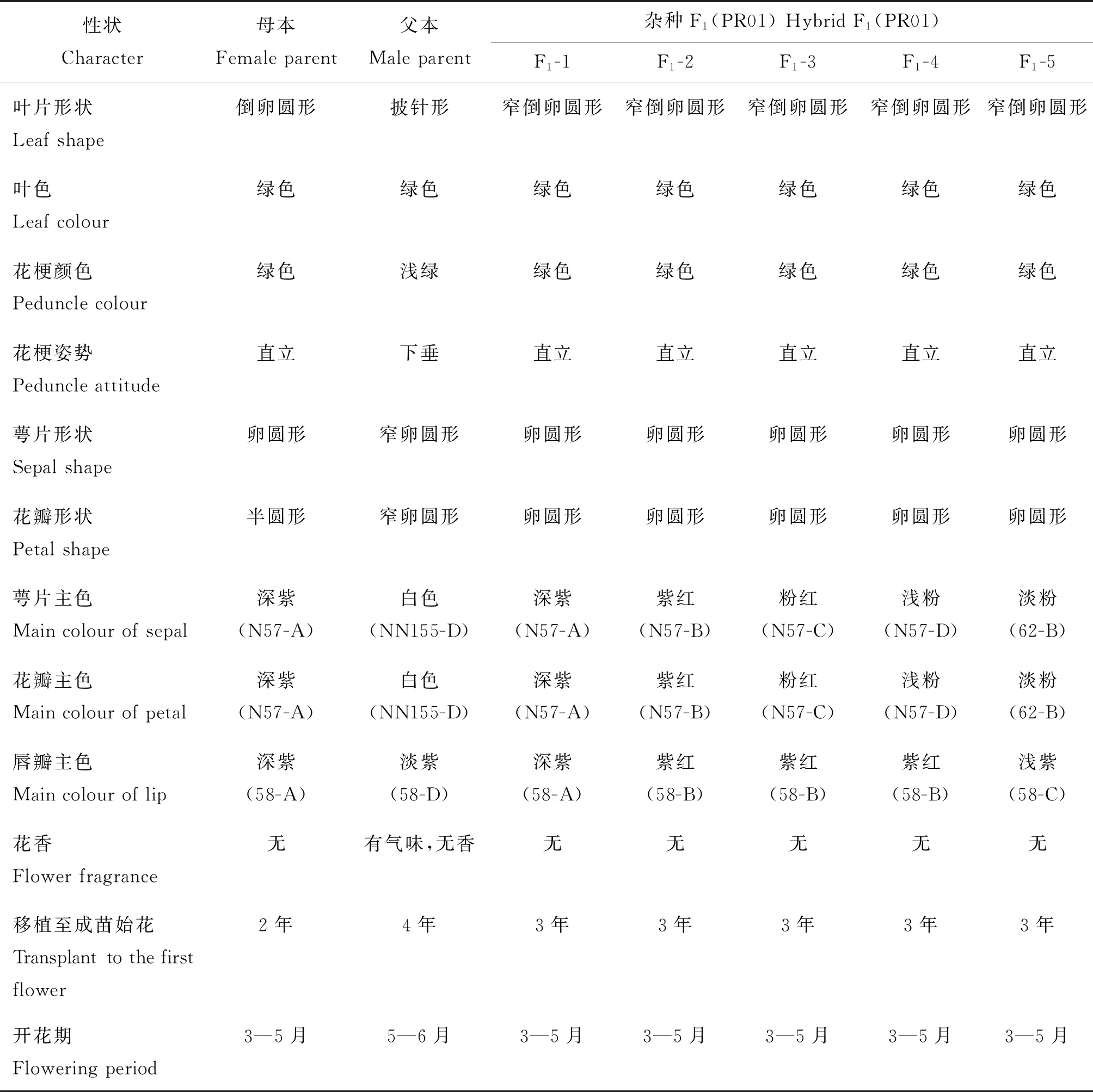
表5 杂交后代株系F1(PR01)与亲本主要假质量性状的比较

(a)母本蝴蝶兰‘聚宝红玫瑰’植株;(b)父本钻喙兰植株;(c)子代F1(PR01)的植株;(d)F1植株(PR01)开花情况;(e)子代F1(PR01)花色分离。
3 讨论与结论
兰花远缘杂交存在属间亲和性低的问题,因此,杂交组合成功的座果率较低[12]。属间杂交亲和性低主要表现在远缘杂交过程中雌性障碍[13]或花粉不育[14],导致提早落果或果荚发育不充分。Kim等[9]在风兰(Neofinetiafalcata)与蝴蝶兰远缘杂交研究中得出,26%的杂交组合中,种子持续生长3~4个月,才能形成正常的种胚。本试验中,以钻喙兰为母本、蝴蝶兰为父本的杂交组合,在授粉杂交后5~10 d全部发育不成功(表1);以蝴蝶兰为母本、钻喙兰为父本的杂交组合,果荚发育时间达到 90~120 d 的仅有1个,为PR01(蝴蝶兰‘聚宝红玫瑰’×钻喙兰),发育时间为110 d(表1),且通过无菌播种并在不同诱导培养基上培养后种子均能萌发(图3(a)、(b)),但发育时间为30~90 d的11个果荚无菌播种后均未萌发(表1,图3(c)和(d)),研究结果与Kim等[9]、陈和明等[10]和Kishor等[22]的结果一致。
兰花种子一般采用无菌播种的方法进行繁殖,由于种子无胚乳,所含营养物质极少,在自然环境中极难萌发[15]。在试验中,12个座果的杂交组合,其果荚分别在M1、M2、M3的诱导培养基上无菌播种后,只有PR01的种子在3种诱导培养基上萌发,且在M2、M3的培养基上,PR01的原球茎形成时间和变绿时间较快,与欧阳英等[16]、丘亮伟等[17]的研究结果一致。在PR01种子的增殖过程中进行了不同浓度的6-BA和Ad配比研究,结果表明处理L2和L3,即BA 2.0~3.0 mg/L、Ad 2.0~3.0 mg/L有利于原球茎的增殖(表2),这与王玲等[18]、崔广英等[19-20]的研究结果相似,即6-BA和NAA搭配使用,增殖效果更佳。
壮苗生根培养能够提高出苗成活率,此阶段一般需要降低培养基的激素或生长调节剂的浓度,以利于形成健壮幼苗。李金雨等[21]认为,生根培养基MS+NAA 0.5 mg/L+10%香蕉泥与MS+IBA 0.5 mg/L+10%香蕉泥对生根的影响,两者之间无差异,其余的处理间均在0.01水平上存在极显著差异,其中以NAA 0.5 mg/L+10%椰子汁的培养基对生根的效果最好,平均生根数达3.6 条;同时,试验结果进一步表明,椰子汁或香蕉泥等对生根具有良好的促进作用。本试验中,壮苗生根培养基T2,即花宝1号3 g/L+NAA 0.5 mg/L,同时添加蔗糖30.0 g/L+椰汁100.0 ml/L+琼脂7.0 g/L最有利于PR01壮苗生根,平均根数量达到3.0 条,根长4.0 cm(表3),说明较低浓度的NAA并添加适量的椰子汁促进了PR01后代株系的生根壮苗。
PR01远缘杂种F1株系在叶片大小、花梗粗、单枝花朵数和花径等数量性状,以及花色(包括花萼、花瓣和唇瓣)、叶色、花梗颜色、叶片形状、萼片形状、花瓣形状等假质量性状均介于双亲之间,这与已有研究结果相似[22]。兰花属间杂交后代的花色、花梗直立和香味被认为是定性和定量遗传[23],本试验中,母本蝴蝶兰‘聚宝红玫瑰’的花色为深紫红,父本钻喙兰花色主要表现为白色,在PR01远缘杂种F1中,大部分株系均表现接近蝴蝶兰的花色性状,而以白色为主的钻喙兰花色在F1中也有体现(图4(d)和(e)),花梗直立在F1中与母本一致;相比之下,母本无香味,父本钻喙兰有气味但不香,但子代F1中无香味也无气味(表5),表明气味或香味的遗传相对复杂,必须进一步研究。在远缘杂交后代开花方面,Kishor等[22]在火焰兰与万代兰远缘杂交的研究中发现,杂交后代幼苗移栽后种植4年10个月,仅有10%的植株形成花蕾并成功开花,陈和明等[10]在火焰兰与蝴蝶兰远缘杂交中,杂交后代移植3年后,其开花率为9.8%~37.3%,而本试验中远缘杂种PR01移植3年后,其开花率为15.6%,表明蝴蝶兰与钻喙兰远缘杂种F1株系间开花时间不同,出现了性状分离。
本研究共设计远缘杂交36个组合,仅有1个杂交组合的果荚由于发育时间充分获得了种子,经无菌播种后种子萌发并获得实生苗,杂种后代经过3年的种植后开花,其花色及植株性状产生了分离。同时,蝴蝶兰与钻喙兰的远缘杂交成功率低,可能与种胚有没有形成、胚发育是否正常、胚发育是否充分或胚龄等有关。
猜你喜欢
林业科学研究(2022年4期)2022-08-03
中国农业科技导报(2022年4期)2022-06-14
麦类作物学报(2022年5期)2022-05-20
蔬菜(2021年3期)2021-11-26
种子(2021年6期)2021-07-16
文苑·经典美文(2019年9期)2019-09-09
天然产物研究与开发(2018年11期)2018-11-30
疯狂英语·新读写(2018年2期)2018-09-07
上海农业学报(2017年3期)2017-04-10
语文世界(小学版)(2015年2期)2015-04-07
