
生长季放牧对高寒草甸传粉网络的影响
2021-04-08 02:14高二亮李昕蔚杨丽莉赵志刚路宁娜
生态学报 2021年4期
高二亮,毕 柽,李昕蔚,杨丽莉,刘 乐,姚 明,赵志刚, *,路宁娜
1 兰州大学生命科学学院,草地农业生态系统国家重点实验室, 兰州 730000 2 西北师范大学生命科学学院, 兰州 730000
植物-传粉者相互作用对生物多样性的产生和维持来说十分重要[1]。大约87%的开花植物依赖于动物传粉者进行有性生殖[2],而传粉者也能从开花植物中取得食物报酬(如花蜜,花粉等),因此保护植物与传粉者之间这种互惠的相互作用能够起到一举两得的效果。群落中多种植物和多种传粉者之间复杂的相互作用构成传粉网络[3]。传粉网络的拓扑结构能够影响生态系统的稳定性和功能[4]。例如,传粉网络一般具有嵌套性结构[5],即网络中的物种具有优先与泛化物种建立连接的倾向, 使得网络中特化物种的连接伙伴是泛化物种连接伙伴的一个子集。有研究认为,传粉网络的嵌套性结构能够维持生态系统的稳定性并且有利于物种共存[6-7]。因此,对于植物-传粉者的相互作用的研究将有助我们更好地理解环境变化会如何影响生物多样性。由于气候变化和人类活动的影响,传粉者面临着全球性的衰减[8],传粉网络也受到如生境丧失[9]、砍伐森林[10]以及生物入侵等[11]类活动导致的环境变化的影响,更好地理解这些人类活动干扰对传粉网络影响的过程和机理对保护生物多样性以及维持生态系统功能具有重要意义。
放牧是是一项在全球范围内普遍存在的人类活动,是造成草地植物群落结构和组成改变的重要干扰因素[12-13]。食草牲畜能够通过取食、踩踏和排泄等方式直接影响植物群落[14-15],而植物群落的改变会进而影响传粉者的多度和多样性[16],最终改变传粉网络的结构[17]。Yu等在蒙古研究了放牧对草原生态系统传粉功能的影响,他们发现放牧降低了网络的多样性[18]。而在苏格兰地区的林地生态系统中,Vanbergen等却得出不同的结果,他们发现放牧增加了传粉网络中相互作用的多样性和泛化程度[19]。生态群落是动态变化的,不同种类的传粉者和开花植物具有不同的物候期,因此导致传粉网络在时间尺度上也是一个动态变化的系统,在不同的时期具有不同的物种组成和相互作用模式[20-21]。不同物种对放牧有不同的响应[22],因此放牧在生长季的不同时期对传粉网络可能有不同的影响。
近年来,因过度放牧和气候变化等因素,青藏高原的高寒草甸生态系统出现了严重的退化[23-24]。过度放牧造成草地生产力、植被盖度以及多样性的降低,最终导致草地的退化[25- 27]。先前在该区域的研究主要关注放牧如何影响草地植被和地下土壤等方面,而对于放牧如何影响生态系统的功能,尤其是传粉功能的研究很少。理解在青藏高原高寒草甸生态系统中放牧如何影响传粉网络对维持草地生态系统稳定和可持续的生产具有重要意义,并且能够为退化草地制定合理的放牧制度提供依据。本研究在青藏高原东部的甘肃省甘南州玛曲县境内选取了两个研究样点,通过连续三年对比禁牧和放牧条件下的样地,拟解决以下科学问题:(1)在高寒草甸生态系统中,放牧如何影响传粉者群落?放牧对不同类群传粉者的影响有何差异?(2)放牧对高寒草甸生态系统中传粉网络的结构有何影响?(3)传粉网络在不同月份之间是否有差异?放牧对传粉网络的影响是否在月份之间是否一致?已有研究表明在青藏高原地区放牧能够降低植物群落的多样性[25-26],也能够导致传粉者数量以及传粉者食物资源的减少[28],因此我们预测放牧会降低传粉网络的多样性和泛化程度。
1 材料与方法
1.1 研究地点
本研究在位于甘肃省甘南州玛曲县境内的兰州大学高寒草甸与湿地生态系统定位研究站的阿孜站(35°58′ N, 101°53′ E, 海拔3500 m)展开。研究区域内年均温为1.2℃,年均降水670mm, 植被类型为典型高寒草甸,主要优势物种包括线叶嵩草(Kobresiacapillifolia)、甘肃薹草(Carexkansuensis)、垂穗披碱草(Elymusnutans)、小花草玉梅(Anemonerivularis)、高原毛茛(Ranunculustanguticus)以及钝苞雪莲(Saussureanigrescens)等[29]。牦牛是本研究区域中唯一的放牧牲畜。在研究区域内选取了两个具有相似地形和植被的研究样点(标记为样点1与样点2,样点之间相距1km以上),每个样点包括一块放牧样地和一块禁牧对照样地,禁牧样地自2011年起,由大约100m × 100m的铁丝网围挡以阻止牲畜进入,而在冬季(非生长季,11月至次年3月)则允许牲畜进入取食干草。放牧样地选取在禁牧样地围栏外相邻的区域,全年放牧。在每个样地内,随机选取一块20m × 25m 的区域进行调查。2016年的7月和8月在2个样点的4个样地进行了调查取样,而在2017以及2018年的7月和8月则只在样点1的2个样地进行了调查取样,共计调查了16个传粉网络。
1.2 传粉者观测
利用摄像机定时拍摄的方法来对传粉者的访花行为进行观测。每个样地的每种植物选取5个(2016、2018)或8个(2017)植株进行观测。观测在晴朗无风的天气状况下每天10:00—17:00期间进行,在每个植株上随机选取一个花单位(floral unit, 即单花、头状花序、伞状花序或穗状花序)进行45mins(2016、2017)或60mins(2018)的拍摄记录。在室内对野外拍摄的视频进行观察,如果视频中有访花昆虫接触到植物的花药或柱头,则记录为一次传粉行为。我们在每个样地随机选取3个0.5m × 0.5m的样方(2016、2017)或3条1m × 5m的样带(2018),并记录其中每个物种的花单位的数目。利用传粉者物种j对植物物种i的单花访问频率(植物i的每个花单位上每小时接受到传粉者j的访问次数)乘以植物i的花单位在群落中的相对密度(植物i的花单位数目/群落中总的花单位数目)来计算物种水平上植物i与传粉者j的相互作用频率(interaction frequency),并利用每种植物与每种传粉者之间的相互作用频率构建了每个样地的植物-传粉者相互作用矩阵(传粉网络)。
1.3 传粉网络参数
利用R语言[30]中的‘Bipartite’[31]包中的networklevel函数计算了以下网络结构参数:(1)植物种类数。(2)传粉者种类数。(3)网络连接数,即网络中植物与传粉者之间建立的连接的种类数。(4)相互作用多样性(interaction diversity),即网络中相互作用的Shannon-winner指数。(5)连接度,表示网络的填充度,即连接数/(植物种类数 × 传粉者种类数)。(6)嵌套性,所选取的衡量嵌套性的指标为NODF值[32]。网络的嵌套性受网络大小的影响较大,为了在比较不同处理间的网络嵌套性时排除网络大小因素的干扰,利用‘Bipartite’包中的nullmodel函数对每个网络构建了1000个零模型,零模型的连接度和网络大小与实际观察的网络相同,但相互作用在网络中是随机分布的。通过将实际观察到的网络与它的零模型进行对比计算出网络嵌套性的z值,计算公式为:
z=[x-μ]∕σ
式中,x为实际观察到的网络的嵌套度,μ为1000个零模型的嵌套度的均值,σ为1000个零模型嵌套度的标准差。(7)网络特化程度(H2′)[33],取值为0—1,0表示网络最泛化,1表示网络最特化。(8)植物泛化程度(generalism of plants),表示网络中每种植物所具有的连接伙伴数量的加权平均数,值越大表示网络中的植物越泛化[34]。
1.4 统计分析
利用双因素方差分析检验了处理和月份以及其交互作用对于传粉网络结构参数以及各个类群传粉者物种数的影响。处理包含两个水平,即放牧处理和禁牧对照。月份也包含两个水平,即7月和8月。方差分析在R语言中利用anova函数进行。
2 结果
2.1 禁牧样地与放牧样地的传粉者群落
在为期三年的实验中共计观察了33种植物(表1),在拍摄的视频中共观测到传粉者66种,其中双翅目34种,膜翅目10种,鳞翅目10种,鞘翅目8种,其他目4种。其中禁牧样地共有传粉者59种,包括双翅目29种,膜翅目10种,鳞翅目10种,鞘翅目6种,其他目4种;放牧样地共有传粉者37种,包括双翅目18种,膜翅目10种,鳞翅目2种,鞘翅目4种,其他目3种(图1,图2),无论是禁牧样地还是放牧样地,传粉者群落都是双翅目占优势(图1,图3),主要包括蝇类(Fly)以及食蚜蝇类(Hoverfly)功能群。放牧后植物种类数(平均值±标准误:禁牧8.75±0.25;放牧7.625±0.46)以及传粉者种类数(禁牧17.875±1.156;放牧11±0.886)都显著降低(图4)。具体来看,放牧降低了双翅目(禁牧10.25±1.082;放牧6.125±0.811)、鞘翅目(禁牧1.875±0.227;放牧1±0.463,边际显著)以及鳞翅目(禁牧1.75±0.559;放牧0.25±0.164)的物种数,但对膜翅目(禁牧3.5±0.378;放牧3±0.535)的物种数没有显著影响(图4)。

表1 植物名录及编号
2.2 放牧对高寒草甸传粉网络的影响
双因素方差分析结果表明,放牧因素显著降低了传粉网络的连接数(禁牧31.375±1.880;放牧19.625±1.179)、相互作用多样性(图1)以及植物的泛化程度,而增加了网络的连接度(边际显著),对嵌套性z值以及网络特化程度则没有影响(表1,图4),表明放牧能够降低群落中植物和传粉者的物种数以及植物-传粉者之间的相互作用种类数以及多样性,但传粉网络的拓扑结构在放牧干扰下仍能保持相对稳定。
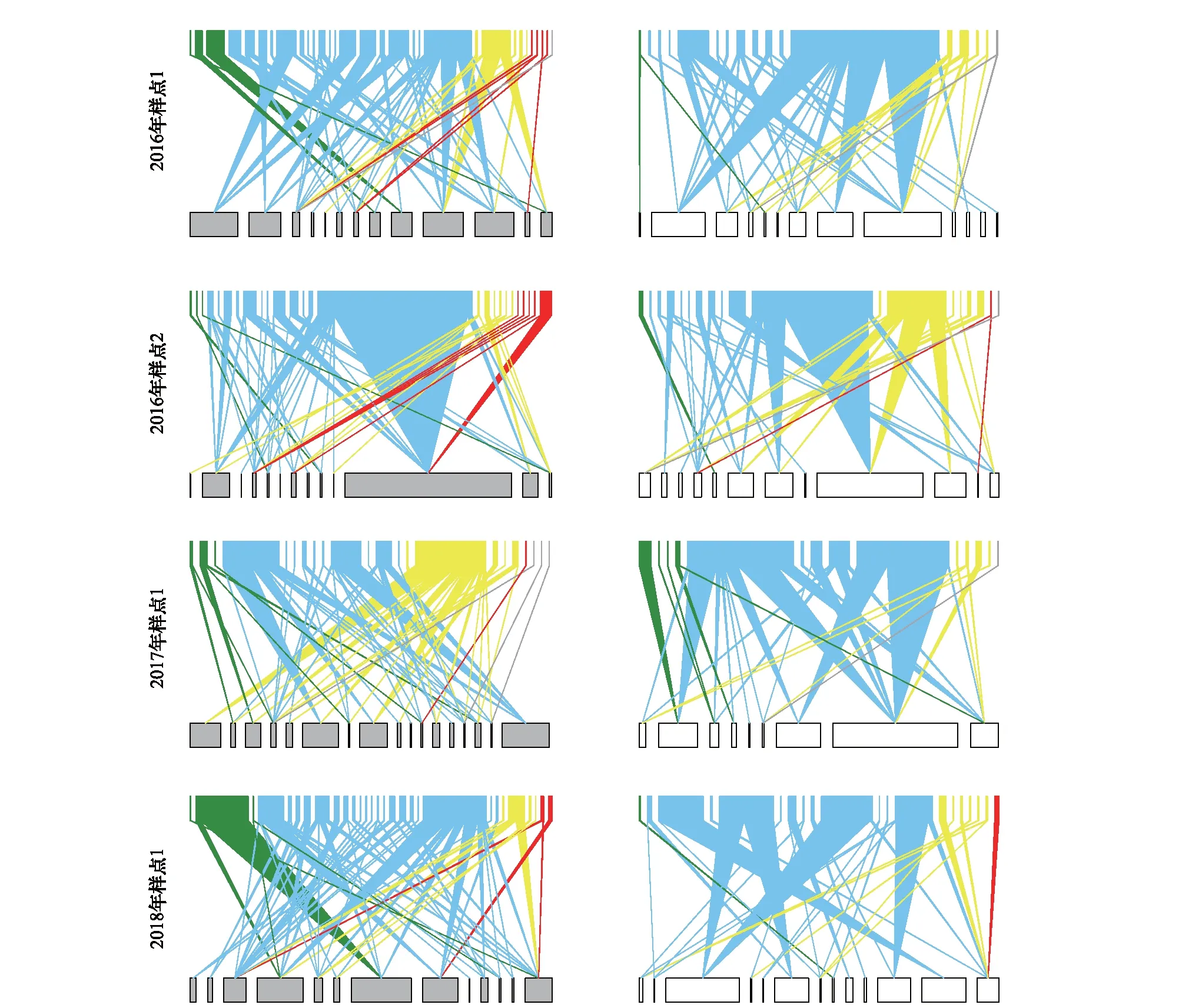
图1 禁牧(灰色条块)与放牧样地(白色条块)月份合并的传粉网络双向图Fig.1 Bipartite plant-pollination networks of ungrazed (grey) and grazed (white) communities summed across months由上至下分别为2016年样点1、2016年样点2、2017年样点1以及2018年样点1中禁牧样地(下部条块为灰色)和放牧样地(下部条块为白色)的传粉网络双向图。位于双向图上部和下部的条块分别表示网络中的传粉者和植物。传粉者中绿色表示鞘翅目、蓝色表示双翅目、黄色表示膜翅目、红色表示鞘翅目、灰色表示其他类群。上下条块之间的连线表示植物与传粉者之间的相互作用。条块及连线的粗细程度表示物种及它们之间的相互作用频率(interaction frequency)的大小
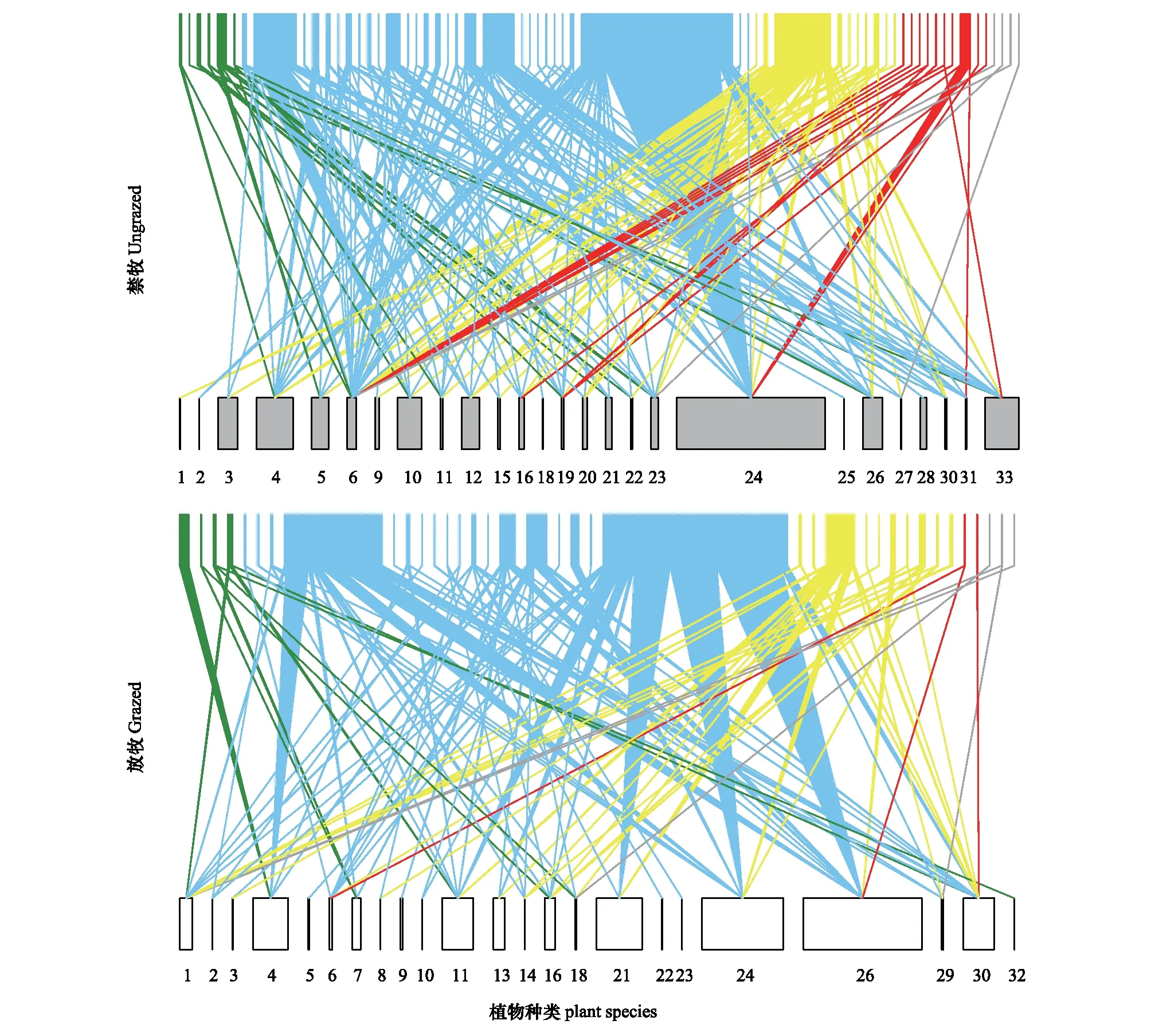
图2 禁牧与放牧样地样点、年份和月份合并的传粉网络双向图Fig.2 Bipartite plant-pollination networks of ungrazed and grazed communities summed across sites, years and months
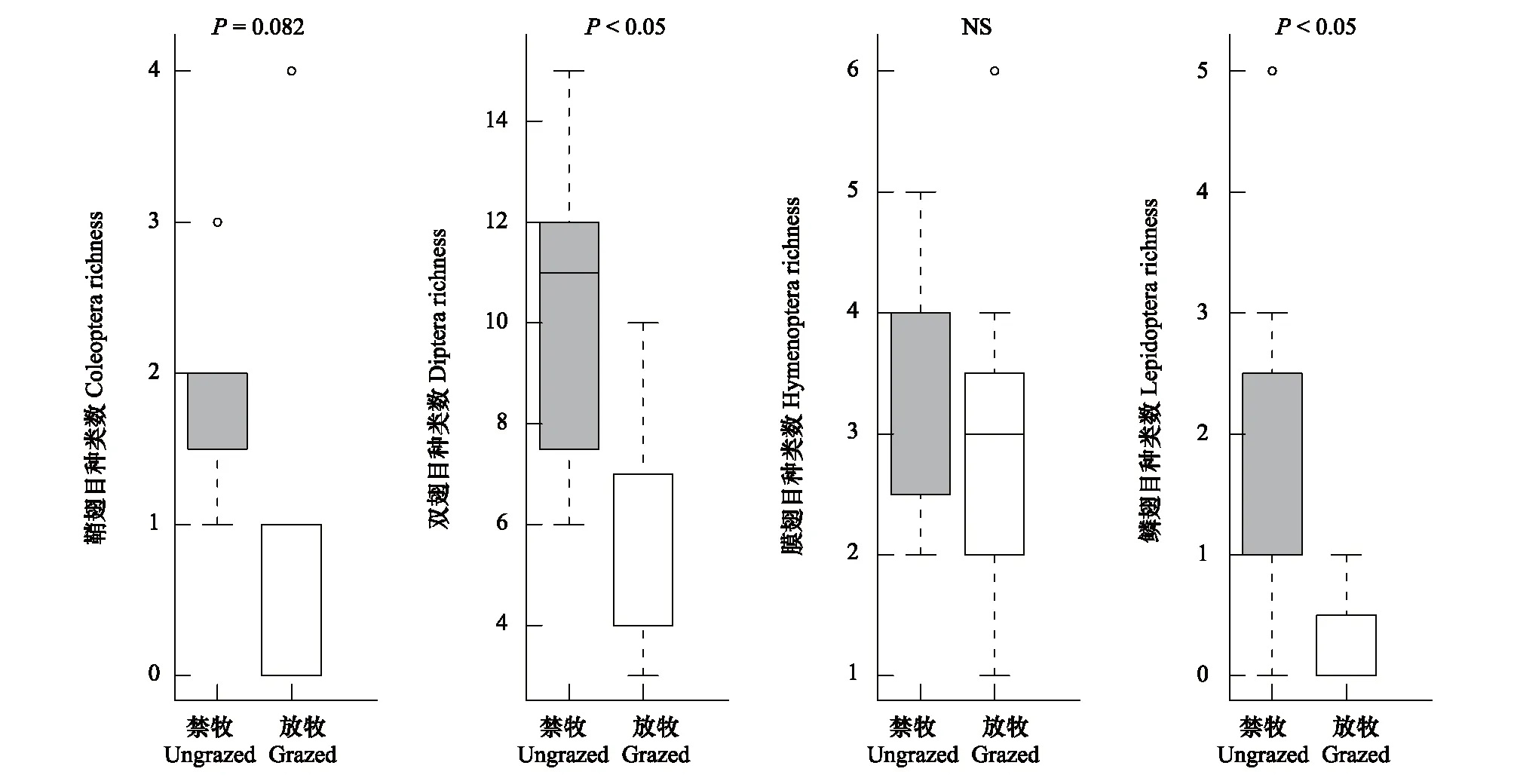
图3 禁牧与放牧样地鞘翅目、双翅目、膜翅目以及鳞翅目传粉者物种数的箱线图Fig.3 The boxplots of species richness of Coleoptera, Diptera, Hymenoptera and Lepidoptera in ungrazed and grazed communities灰色表示禁牧(U)网络,白色表示放牧(G)网络,图上标注了方差分析结果的P值,NS表示差异不显著
2.3 月份对高寒草甸传粉网络的影响
月份对所有网络指标都没有显著的影响(表2),表明传粉网络中的物种数和多样性以及拓扑结构在不同月份之间能保持相对稳定;对所有网络指标,处理和月份之间都没有显著的交互作用(表2),表明在不同月份下放牧对于传粉网络的作用都是相同的。
3 讨论
3.1 放牧对高寒草甸传粉者群落的影响
研究区域中,无论是禁牧样地还是放牧样地,传粉者群落都是双翅目占优势,主要包括蝇类(Fly)以及食蚜蝇类(Hoverfly)功能群。双翅目昆虫对强风、低温、高强度的光照等高海拔地区的气候条件具有较强的适应能力,这使其成为高海拔地区主要的传粉者类群[35],许多高寒地区的相关研究也表明双翅目昆虫是当地主要的传粉者类群[36-37]。放牧显著减少了双翅目、鳞翅目以及鞘翅目传粉者的种类数(图3),而对膜翅目传粉者没有显著影响。膜翅目昆虫通常具有较强的飞行能力[38],使得其在禁牧与放牧样地之间流动性较大,因而放牧干扰对其造成的影响较小。相反地,双翅目、鳞翅目以及鞘翅目昆虫飞行能力相对较弱,使得其物种数在放牧后显著减少,这降低了传粉者的多样性,可能对植物的繁殖产生不利影响。而且,放牧后鳞翅目和鞘翅目传粉者的种类数降到了很低的水平(图3),相关的研究也表明放牧会显著降低草地中鞘翅目昆虫的数量和多样性[39]。尽管鳞翅目和鞘翅目昆虫不是高寒地区的主要传粉者类群[35],但其在维持草地生态系统食物网的稳定方面仍具有重要的作用[6],因此不合理放牧导致的昆虫多样性丧失,会对生物多样性的保育以及生态系统的稳定性造成不利影响。
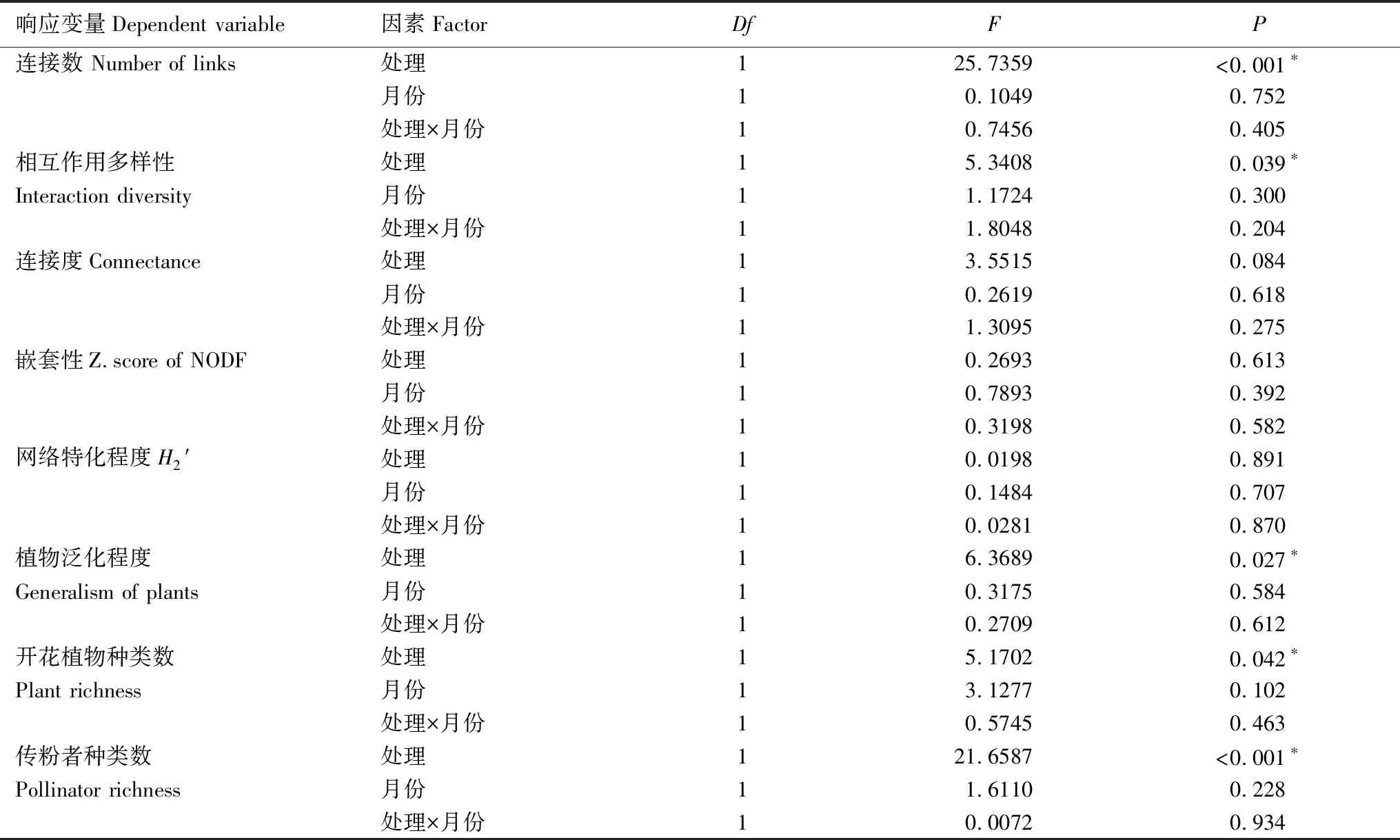
表2 处理与月份及其交互作用对传粉网络的影响双因素方差分析结果
3.2 放牧对传粉网络的多样性和稳定性的影响
与预测一致,放牧降低了传粉网络的多样性,植物种类数、传粉者种类数、植物-传粉者相互作用种类数和多样性都在放牧后降低(图4)。由于植物和传粉者丰富度在放牧后的减小,尽管网络连接度在放牧后有所增加,网络中实际实现的连接数量仍然显著降低。植物-传粉者相互作用的多样性受到相互作用种类数以及相互作用均匀度的影响,本研究中相互作用多样性在放牧后的降低主要是由于相互作用种类数的降低,因为我们通过方差分析发现相互作用的均匀度在处理之间没有显著差异(F=0.1809,P=0.678)。放牧同时降低了传粉网络中植物的泛化程度,但是网络整体的特化程度(H2′)却没有受到放牧影响。植物泛化程度,表示网络中每种植物所具有的连接伙伴数量的加权平均数,可以理解为网络中植物的连接伙伴的多样性,植物的连接伙伴多样性越高,则植物越泛化[34]。而网络特化程度(H2′)则衡量的是实际观察到的网络中物种之间的相互作用模式与根据相互作用矩阵中每个物种的边际总和预测生成的网络相互作用模式之间的差异大小,即网络中相互作用的选择性的大小或者说物种之间相互作用的专一性,选择性、专一性越高,网络则越特化[33]。特化的网络意味着网络中物种之间的功能互补性增加,冗余程度减小,网络的恢复力减小[40]。此外,发现放牧没有影响传粉网络的嵌套度,而增加了网络的连接度(边际显著),这两个值都与网络的稳定性有关,嵌套度、连接度越高,网络越稳定[6, 41]。因此,尽管网络中每种植物平均的传粉者多样性在放牧后降低了,但是网络的稳定性和恢复力并没有受到放牧的影响。
国外已有一些学者研究了放牧对于传粉网络的影响,与本研究比较相似的一个研究来自Marrero等在阿根廷潘帕斯地区的研究,他们对比了每年9月到次年4月禁牧的样地与全年放牧的样地,发现,放牧样地的植物和传粉者种类数与禁牧样地之间没有差异,但放牧样地的传粉网络具有更高的特化程度(H2′)[42],这与我们的研究结果相反。国内学者也有相关研究,Xie等人在四川省红原县的高寒草甸上的研究发现,夏季放牧降低了熊蜂属的物种多样性以及其食物资源的可利用性[28],这与我们的研究结果类似。虽然许多研究都关注了放牧对于传粉网络的影响,但这些研究却没有得出一个广泛适用的结论,这可能是因为这些研究中的气候及生境类型、食草动物的种类以及放牧强度都不一致。Amparo等人在希腊莱斯沃斯岛上11个具有不同放牧强度的常绿矮灌木丛群落中调查了传粉网络,发现放牧对传粉网络的影响符合中度干扰假说[43-44],网络在中等强度的放牧条件下多样性最高[45]。研究中放牧降低了传粉网络的多样性,说明在该研究区域,放牧样地的放牧强度已经超过了最适强度,随着放牧强度的继续增加,网络的多样性将进一步下降。
3.3 高寒草甸传粉网络在时间尺度上的动态
实验结果表明,高寒草甸生态系统的传粉网络在时间尺度上具有一定的稳定性,网络的多样性和泛化程度以及嵌套性等结构在两次不同时间点之间都没有显著差异,并且放牧在不同月份之间对于传粉网络的影响是相似的(表2)。尽管高寒草甸的植物群落开花物候相对分离,具有较高程度的物种更替[46],但传粉网络的结构在7月和8月两次调查之间能保持稳定,说明虽然物种组成有差异,但网络中原有物种的角色和功能能够被其他更替的物种所代替。研究结果与Basilio等人的研究类似,他们一年内对传粉网络的季节动态进行了观测,发现网络的结构参数保持相对的稳定[47]。需要注意的是研究只在当地生长季的盛花期进行了两次观测取样,生长季初期和末期的传粉网络有待进一步研究。
4 结论
研究在青藏高原高寒草甸生态系统探讨了放牧对传粉网络的影响。研究的结果表明,高寒草甸生态系统中,传粉者群落的物种组成以双翅目昆虫为主,而双翅目中主要包括蝇类(Fly)以及食蚜蝇类(Hoverfly)功能群。放牧减少了双翅目、鳞翅目以及鞘翅目访花昆虫的物种数,但对膜翅目访花昆虫没有显著影响。放牧对于传粉网络具有负面作用,体现在网络中植物、传粉者以及它们之间的相互作用的种类和多样性的减小,但网络的拓扑结构没有显著变化,说明网络的稳定性和恢复力在放牧后仍能保持相对的稳定。研究结果表明该区域放牧强度已超过最适强度,对生物多样性造成了不利影响。未来需要深入研究高寒草甸生态系统中放牧强度对传粉网络的影响模式,以期为合理的放牧制度模式提供理论参考。
猜你喜欢
中国科学探险(2022年8期)2022-12-31
湖北植保(2022年4期)2022-08-23
空军工程大学学报(2021年4期)2021-09-23
土壤(2021年1期)2021-03-23
新疆大学学报(自然科学版)(中英文)(2020年2期)2020-07-25
中国农村水利水电(2020年4期)2020-06-12
大科技·百科新说(2018年3期)2018-04-11
电影文学(2017年24期)2017-11-16
金融经济(2017年7期)2017-07-15
福建农业学报(2016年6期)2016-11-01
