
水稻卷叶基因研究进展
2021-04-20 05:57李蓓莫凯琴马银花
安徽农学通报 2021年6期
李蓓 莫凯琴 马银花
摘 要:水稻是禾本科单子叶C3植物,是我国主要的粮食作物之一。叶片是水稻进行光合作用、呼吸、蒸腾等活动的重要场所,是决定稻米产量的重要的工艺性质,而卷叶是水稻的一种特殊性质。研究显示,水稻叶片的适度卷曲对叶片挺直有利,改善了披捶状态,增加了光合作用,提高了水稻的产量。该文综述了水稻叶片卷曲的原因,阐述了水稻叶片卷曲的细胞学形成机制及相关基因的分子机制,以期为水稻叶片性状的研究与应用提供参考。
关键词:水稻;卷叶;泡状细胞;育种
中图分类号 Q89文献标识码 A文章编号 1007-7731(2021)06-0014-05
Advances in Gene Research of Rice Roll Leaf
LI Bei et al.
(School of Agriculture and Biotechnology/Hunan Provincial Collaborative Innovation Center for Field Weeds Control, Hunan University of Humanities, Science and Technology, Loudi 417000, China)
Abstract: Rice is a kind of gramineae endogen C3 plants, is one of the main food crops in our country. The rice leaf photosynthesis, respiration and transpiration is important places, is to determine an important technological properties of rice production, and the leaf is a kind of special properties of the rice. Studies have shown that the moderate curl of rice leaves is beneficial to the straightness of rice leaves, which improves the state of the pendulum and increases the photosynthesis and single yield of rice. In this paper, the causes of rice leaf curl were summarized, and the cytological formation mechanism of rice leaf curl and the molecular mechanism of related gene molecules were expounded in order to further elucidate the cytological and molecular mechanism of rice leaf curl and promote the application of rice leaf shape.
Key words: Rice; Curly leaf; Vesicular cell; Breeding
水稻屬于禾本科植物,是全球一种重要的粮食作物[1],是理想单子叶模式植物[2],可自花授粉,易转化农杆菌,易栽培和繁殖。与其他禾本科植物相比,水稻具有不同的种类和突变性,基因组是同线性的[3]。
水稻的理想株型在培育过程中受到多方面的影响,如叶形、株型等。叶片形态改进,直接或间接地影响作物的光合作用,适当的叶片卷曲使其直立,可提高光合效率,加速干燥物质的积累,增加粮食产量[4]。袁隆平院士提出了水稻超高产的理想株型,而理想株型则需要上3叶“长、直、窄、凹和厚”[5],其中凹指水稻的叶片内卷,这种性状有效地减少了种植群体内的遮挡率,提高了群体的光合效果,促进了群体空气的流通,减少了蒸腾,延缓了叶片衰老[6]。
在水稻生产中,卷叶是一个重要的农艺性状,适当的卷叶能够有效地提高光合作用和作物产量。根据以往对稻卷叶的研究,发现影响水稻卷叶的因素有很多,如非生物威胁、生物威慑力和基因遗传、泡状细胞等。因此,在水稻的理想株型培育过程中,不仅可以控制泡状细胞的大小、尺寸,还可以控制叶片的卷曲度,也可以通过对非生物条件进行光照、温度、湿度和重金属进行调整,同时也可以通过真菌、细菌和病毒入侵的方式影响叶片的卷曲。除此之外,还有一些受基因控制着。
1 水稻卷叶类型及细胞学形态变化
1.1 叶片的发育 植物的叶片形态有多种,其内部的发育能力和植物在响应内或外部信号的过程中具有动态和灵活性。因此,即使是2个基因类型相同的植物,也不一定会产生相同的叶片[7]。叶片在植物生长过程中起着至关重要的作用,其正常发育过程是一个复杂的过程,包括从顶部分生组织周围区的开始和决定,叶极性建立,细胞分裂伸长和叶脉形成,探明叶片的发育机理有助于株形的改造,从而使生物设计中脱离叶形。
1.2 卷叶类型 叶形为重要的农艺特征,直接影响水稻光合作用、呼吸功能和蒸腾作能气体交换、极性运输和耐干旱、盐碱等。水稻叶不仅是光合作用的一种重要的器官,也是稻株形成的一种重要的组成因素。适度的卷曲提高了个人和群体在透光方面的能力,一方面,缩小了叶片和茎秆之间的夹角,使叶片直立,促使上下两面的光线受到改善,反射率下降,提高光合强度;另一方面,叶片的卷曲能保持不披垂,在群体的中后期保持下部光照,提高叶面指数,增强光合作用,提高能量利用率,从而改善水稻的质量。水稻中存在着丰富的卷叶资源,从卷叶的表型来看,大多数是正卷的(叶缘两侧沿叶中脉内卷,形态为半圆筒或圆筒状,另外还有一部分表现为反卷(叶缘沿中脉两侧,叶片外卷为半圆筒或圆筒状);从卷叶出现的部位看,可分为全株叶片均卷型和部分叶位叶(如剑叶)卷曲型;按卷叶的程度来分,有高度筒状卷曲型、中度卷曲型、轻微卷曲型。
1.3 影响水稻卷叶的因素 水稻表型是由外部环境和内部遗传2个因素决定的,相同的基因型水稻表现在不同的环境下,如水稻处于旱季的环境下,植物以平滑叶片卷起的运动方式应对极端威胁,叶片卷曲是一种运动形式,可以减少叶片的水分蒸发,防止脱水叶片,增加干旱环境下植物存活的概率[8-9]。造成植物卷叶表的外部环境因素主要有干旱、高温、辐射和除草剂等非生物威胁,以及昆虫食用、病毒,细菌和真菌感染等;引起植物卷叶表型的内部遗传因素主要有调节叶片极性和细胞结构的基因调节。
1.3.1 非生物胁迫 目前,研究较多的是干旱胁迫导致的叶片卷曲,缺水干旱、气温偏高、强日照、除草剂、臭氧、辐射、重金属等是限制植物生长的主要非生物胁迫因素[10-12]。植物应对干旱或者高温逆境刺激时,植物内部水势情况和渗透调节,使作为动力细胞的泡状细胞通过减少细胞膨胀压降低,导致叶片卷曲,改变叶片有效面积,通过减少蒸腾的作用[13-14]。研究表明,水稻在受到干旱的威胁时,比其他作物更容易卷曲、干燥和产量降低,从而使其受到干旱的威胁[15]。在同一种干旱的胁迫下,菏稻和糯米有不同的形态、生理变化,菏稻有叶卷,菏稻有叶卷,菏稻有可伸展的状态[16]。高盐、低温和渗透压等因素对突变体生长有一定的影响。水稻叶卷与叶片的渗透、温度等因素存在线性关系,高温或低温会导致不同程度的水稻叶卷曲[13]。
1.3.2 生物胁迫 真菌、细菌和病毒的侵染以及虫害等是影响叶片卷曲的主要生物胁迫因素。研究发现,病毒、细菌和真菌感染植物在不同的程度下可引起叶片卷曲,伴随病原体侵入和发育,叶片出现卷曲、萎缩,颜色从灰绿到黄,直至叶片死亡[17]。在细菌感染的研究中,如黄单胞杆菌属感染水稻,最终导致水稻叶片产生细菌性条斑病,病原体的侵染使得植物叶片慢慢变黄,引起叶片卷曲甚至萎蔫[18-22]。
1.4 泡状细胞及其对水稻叶片卷曲的作用 泡状细胞,被认为泡状细胞与叶片的内卷和开展作用有关,又被称之为运动细胞(motor cell),在叶片上排列成若干纵列,是一类具有薄垂周壁且高度液泡化的大型细胞(即薄壁细胞),为单子叶植物所特有(除沼生目外)。其存在位置是2个维管束,位于靠近叶片的轴面。在泡状细胞的中央含有大型中间液泡,其长轴与叶脉平行,无叶绿体或少量。通过对叶片的横切面图进行观察,可以发现,泡状细胞呈簇形,并以折扇状排列。中间部分的细胞体积是最大的,而两侧细胞体积则是最小的[23]。一般情况下,当叶片在蒸腾作用下丢失太多的水分,泡状细胞就会松弛,从而减少蒸腾作用,使叶子向内折叠或卷曲。当蒸腾减少时,泡状细胞又因吸水而膨胀,叶片变得平展。
1.5 卷叶基因 在水稻中,泡状细胞还有一个重要的功能,就是通过控制泡状细胞的数量、大小等来调节叶片的伸展卷曲。截至目前,已经有16个基因被克隆来控制卷叶。在正卷叶片的突变体中,通常会发现较少的或面积更小的泡状细胞,如OW1/NAL7[24-25]、NRL1[25-27]和RL14[28]。在能够控制叶片的反卷基因突变体中,泡状细胞数量增加或同时伴有泡状细胞面积增加的情况,如LC2[29]、OsZHA1[30]、ACL1[31]、ADL1[32]和Roc5[33]突變。当然,也有一些例外,如SRL1[34],基因突变体的泡状细胞增加,但叶片表现为正卷。根据分析,原因是虽然泡状细胞数量增加,但表皮细胞却减少了,不仅仅是一个突变。水稻YABBY1基因利用反馈调节法获取的GA生物合成,参与反馈调节,YABBY1基因共同抑制植物的泡状细胞,导致水稻叶片反卷[35]。OSHBL是ClassIIIHD-ZIP家族中的一个基因,经过表达,水稻叶片呈正卷状,在近轴面上形成泡泡细胞,而厚壁细胞位于远轴上的数量则减少了[36]。
2 泡状细胞相关的分子机理
2.1 与泡状细胞发育相关的基因 在水稻中,泡状细胞一个重要的功能是通过控制泡状细胞的数目、大小等来调控叶片的卷曲。目前,已成功克隆出的16个可调控卷叶的基因中存在与泡状细胞发育有关的基因数目为13个。通常在能控制叶片内卷基因的突变体中,泡状细胞面积会减小,如COWI/NAL7,NRLI,RLI4[24-28]。NARROW AND ROLLED LEAF1(NRL1)以及ROLLED LEAF 14(RL14),编码1个纤维素合酶和2OG-Fe(I)氧化酶,分别在它们的突变体中正调控泡状细胞的发育,从而导致泡状细胞的变小,引起水稻叶片的内卷[36-37]。OsYABBY6是泡状细胞发育和叶片伸展所必需的,并且参与了叶片近轴面的发育调控[38]。
在能控制叶片外卷的基因突变体中,泡状细胞的数目增加或同时伴有泡状细胞面积增加的情况,如LC2,OsZHDI,ACLI,ADL1和Roc5[39-43]。如过表达Abaxially Curled Leaf 1(ACL1)能导致水稻叶片中泡状细胞的数目增加以及面积增大,从而引起水稻叶片向外卷曲。而过表达水稻中ACL1的唯一的同源基因ACL2,也会导致水稻叶片发生一致的表型[28]。OsZHD1编码1个锌指结构同源盒转录因子,过表达OsZHD1能够导致在水稻近轴的细胞层泡状细胞数目增加,进而引起水稻叶片外卷[44]。
但这样的规律并不是适用于所有的突变体,比如SRLI基因,虽然其突变体泡状细胞数目增加,但叶片却表达为内卷叶,其原因是泡状细胞数增多,而表皮细胞数目却减少,致使近轴面的细胞总量未发生变化[45]。不仅是突变体,还有利用反向遗传学方法获得的水稻YABBY1基因,参与反馈调节中的GA生物合成,当YABBY1基因的共同抑制植物泡状细胞数目增多时,致使水稻叶片发生外卷[46]。OsHB1是HD-ZIPII家族的基因之一,该基因过表达后叶片表现为内卷,其位于远轴面的厚壁细胞数量减少且在近轴一面异形为泡状细胞[47]。
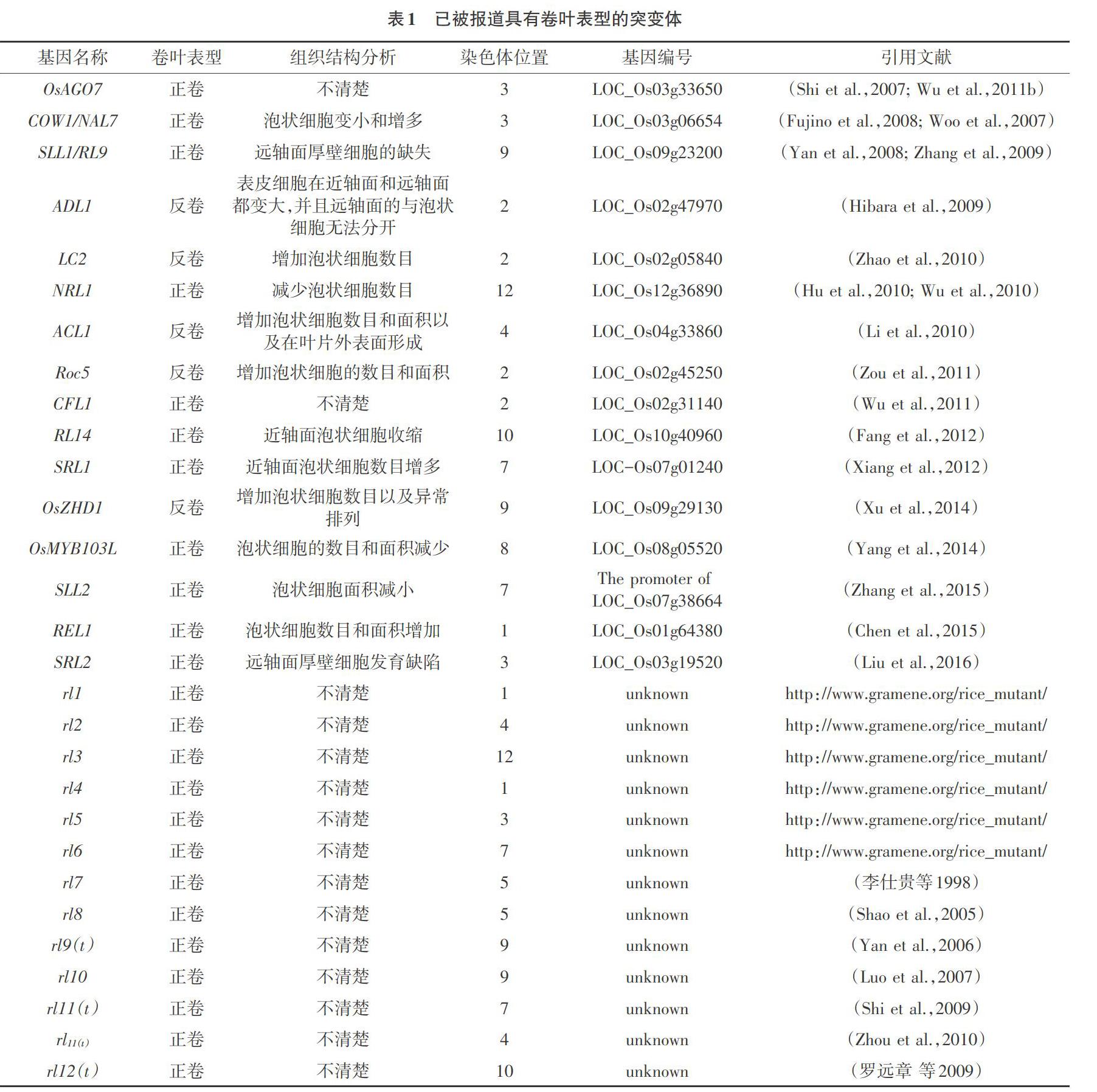
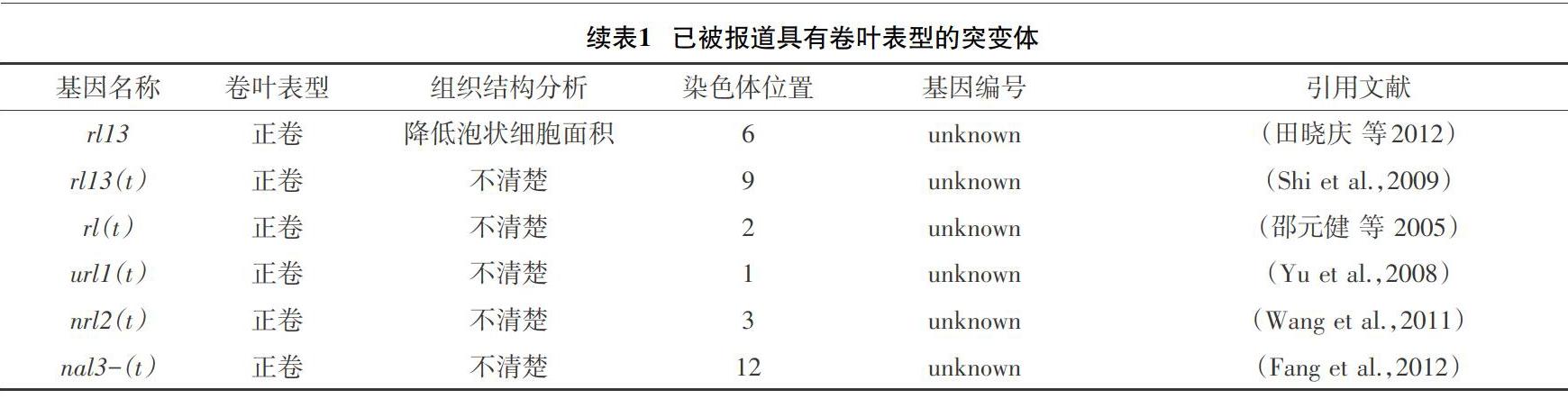
2.2 已经定位和克隆的卷叶基因 截至目前,对水稻叶片卷曲的基因已进行了许多的研究,仅在水稻中就有35种叶卷突变体已被报告,其中16种已经克隆,仍有19种未被克隆(见表1)。目前,大多数成功地定位并克隆的水稻卷叶基因都与水稻泡状细胞相关,只有少部分与近轴或远轴极性的水稻叶片发育有关。
3 水稻卷叶性状与发育的关系
叶片是植物光合作用和蒸腾作用发生的部位,叶片形态是理想株型设计的重要指标之一。在中国的超级稻育种计划中,上部3片叶的微滚动表型是首选,因为稍微弯曲的叶子被认为会增加光合作用区域,从而增加产量。从已有的研究成果来看,过度卷曲的水稻叶片可能会导致减产,而适度的卷曲对于水稻的生长发育来说是有益的,所以育种学家对水稻卷叶的研究从未停止。目前,通过水稻突变体来研究调控叶片卷曲基因的报道越来越多,众多育种成果也开始推广种植。比如,近几年以卷叶不育系培矮64S配制的卷叶杂交稻两优培九、65396已经在南方稻区得到了推广种植[48]。65396已在云南永胜县涛源乡种植,取得了17.07t/hm2的高产纪录[49]。因此,叶片的卷曲性状是育种中重要的选择指标之一。
4 问题与展望
截至目前,经过长时间的研究积淀,已经取得了较丰富的稻卷资源和水稻叶突变体筛选。遗传分析、卷叶基因克隆、基因定位及功能研究已取得了较大的成果,已发现的基因有一部分能够克隆,而由于伴随着其他特征的变异,一些卷叶突变体很难在实际的育种中应用。对于已有水稻卷叶基因,更多的是研究方法和方式,需要继续进行。水稻产量由多个因素共同控制,也就是水稻的品种、栽培环境、叶卷程度和通风状况等都有一定的影响。
水稻叶的卷叶平衡具有多种复杂性,需要有复杂的卷叶控制处理系统加起来才能控制水稻叶片内部细胞膜的结构,并进一步可以控制水稻卷叶平衡是否正常发生。那么要想卷叶达到最好的平衡,该如何控制?
水稻卷叶的性状是由基因本身或有关受体激素的调节而成?又是如何传递信号,调节下游的基因?这些需要使用突变材料深入地研究相关的基因,加深基因和受体激酶之间的关系,基因和受体激酶之间的直接关系,从分子机理学的角度更清楚地揭示卷叶基因的作用。卷叶基因可能通过这种复杂机制对泡状细胞的生长和发育进行调节,从而控制稻叶的卷叶。
基于水稻卷叶的性状,对于提高稻米产量和改善稻米品质具有重要的研究意义,水稻卷叶的性状仍是未来研究的热点。特别是在育种应用上,更要研究和开展卷叶品种的育种。
参考文献
[1]Clark L G,Zhang W,Wendel J F.A phylogeny of the grass family(Poaceae) based on ndhf sequence date [J].Systematic Botany,1995,20:436-460.
[2]Itoh J i,Nonomura K l,Ikeda K,et al.Rice plant development:from zygote to spikelelet[J].Plant Cell Physiology,2005,46(1):23-47.
[3]Bennetzen,Ma J.The genetic colinearity of rice and other cereal on the basis of genomic sequence analysis [J].Current Opinion in Plant Biology,2003,6(2):128-133.
[4]Lang Y,Zhang Z,Gu X,et al. Physiological and ecological effects of crimpy leaf character in rice (Oryza sativa L.) II. Photosynthetic character,dry mass production and yield forming[J].Acta Agronomica Sinica,2004,30:806-810.
[5]Osborn B A,Raven J A.Light absorption by plants and its implications for photosynthesis[J].Biological Reviews,1986,61 (1):1-6.
[6]張俊杰.水稻卷叶突变体sll2的遗传分析及泡状细胞发育调控研究[D].南京:南京农业大学,2015.
[7]Bar,M.,and Ori,N.Leaf development and morphogenesis[J].Development,2014,141:4219-4230
[8]Kadioglu,A.,Terzi,R.A dehydration avoidance mechanism:leaf rolling[J]. Bot Rev 2007,73:290-302.
[9]Kadioglu,A.,Terzi,R.,Sauhan,N.,et al. VB Current advances in the investigation of leaf rolling caused by biotic and biotic stress factors[J].Plant Science,2012,182:42-48.
[10]Kidner C A,Timmermans M C. Mixing and matching pathways in leaf polarity[J].Curr Opin Plant Biol.,2007,10:13-20.
[11]Hu J,Zhu L,Zeng D L,et al. Identification and characterization of NARROW AND ROLLED LEAF 1,a novel gene regulating leaf morphology and plant architecture in rice[J].Plant Mol. Biol.,2010,73(3):283-292.
[12]Kadioglu,A.,Terzi,R,,Saruhan,N.,et al. Current advances in the investigation of leaf rolling caused by biotic and abiotic stress factors[J].Plant Science,2012,182:42-48.
[13]Kadioglu A,Terzi R. A dehydration avoidance mechanism:leaf rolling[J].Botanical Review,2007,73(4):290-302.
[14]Lafitte H R,Price A H,Courtois B. Yield response to water deficit in an upland rice mapping population:associations among traits and genetic markers[J].Theor. Appl. Genet.,2004,109(6):1237-1246.
[25]Moulia,B. The biomechanics of leaf rolling[J].Biomimetics, 1994,2:267-281.
[16]Lian,H.L,Yu,X,Ye,Q.,et al. The role of aquaporin RWC3 in drought avoidance in rice[J].Plant Cell Physiol., 2004,45:481-489.
[17]王丹.水稻類受体胞质激酶LRRK1调控卷叶的功能分析[D].长沙:湖南大学,2017.
[18]Sariyeva G,Kenjebayeva S,Lichtenthaler H K . Adaptation Potential of Photosynthesis in Wheat Cultivars with a Capability of Leaf Rolling under High Temperature Conditions[J]. Russian Journal of Plant Physiology,2019,57(1):28-36..
[19]Lang Y Z,Zhang Z J,Gu X Y,et al. A physiological and ecological effect of crimpy leaf character in rice (Oryza sativa L). II. Photosynthetic character,dry mass production and yield forming[J].Acta Agron Sin., 2004,30:883-887.
[20]Zhang G H,Xu Q,Zhu X D,et al. SHALLOT-LIKE1 is a KANADI transcription factor that modulates rice leaf rolling by regulating leaf abaxial cell development[J].Plant Cell.2009,21:719-735.
[21]Chen Q,Xie Q,Gao J,et al. Characterization of Rolled and Erect. Leaf 1 in regulating leave morphology in rice[J]. J Exp Bot., 2005,66:6047.
[22]Richards R A,Rebetzke G J,Condon A G,et al. Breeding opportunities for increasing the eficiency of water use and crop yield in temperate cereals[J].Crop Sci., 2002,42:111-121.
[23]Itoh J,Hibara K,Sato Y,Nagato Y. Developmental role and auxin responsiveness of class III homeodomain leucine zipper gene family members in rice[J].Plant Physiology,2008,147:1960-1975.
[24]Fujino K,Matsuda Y,Ozawa K,et al. NARROW LEAF 7 controls leaf shape mediated by auxin in rice[J].Molecular Genetics and Genomics,2008,279:499-507.
[25]Woo Y,Park H,Su′Udi M,et al. Constitutively wilted 1,a member of the rice YUCCA gene family,is required for maintaining water homeostasis and an appropriate root to shoot ratio[J].Plant Molecular Biology,2007,65:125-136.
[26]Hu J,Zhu L,Zeng D,et al. Identification and characterization of NARROW AND ROLLED LEAF 1,a novel gene regulating leaf morphology and plant architecture in rice[J].Plant Molecular Biology,2010,73:283-292.
[27]Wu C,Fu Y,Hu G,Si H,Cheng S,Liu W. Isolation and characterization of a rice mutant with narrow and rolled leaves[J].Planta,2010,232,313-324.
[28]. Fang L,Zhao F,Cong Y,et al. Rolling-leaf14 is a 2OG-Fe (II) oxygenase family protein that modulates rice leaf rolling by affecting secondary cell wall formation in leaves[J].Plant Biotechnology Journal,2012,10:524-532.
[29]Zhao S,Hu J,Guo L,Qian Q,et al. Rice leaf inclination2,a VIN3-like protein,regulates leaf angle through modulating cell division of the collar[J]. Cell Research,2010,20:935-947.
[30]Xu Y,Wang Y,Long Q,et al. Overexpression of OsZHD1,a zinc finger homeodomain class homeobox transcription factor,induces abaxially curled and drooping leaf in rice[J]. Planta,2014,239:803-816.
[31]Li L,Shi Z,Li L,Shen G,Wang X,An L,Zhang J. . Overexpression of ACL1 (abaxially curled leaf 1) increased bulliform cells and induced abaxial curling of leaf blades in rice[J].Molecular Plant,2010,3:807-817.
[32]Hibara K,Obara M,Hayashida E,et al. The ADAXIALIZED LEAF1 gene functions in leaf and embryonic pattern formation in rice[J].Developmental Biology,2009,334:345-354.
[33]Zou L,Sun X,Zhang Z,Liu P,et al. Leaf Rolling Controlled by the Homeodomain Leucine Zipper Class IV Gene Roc5 in Rice[J]. Plant Physiology,2011,156:1589-1602.
[34]Xiang J J,Zhang G H,Qian Q,et al. SEMI-ROLLED LEAF1 Encodes a Putative Glycosylphosphatidylinositol-Anchored Protein and Modulates Rice Leaf Rolling by Regulating the Formation of Bulliform Cells[J]. Plant Physiology,2012,159(4):1488-1500.
[35]Dai M,Zhao Y,Ma Q,Hu Y,et al. The rice YABBY1 gene is involved in the feedback regulation of gibberellin metabolism[J].Plant Physiology,2007,144:121-133.
[36]Itoh J,Hibara K,Sato Y,et al. Developmental role and auxin responsiveness of class III homeodomain leucine zipper gene family members in rice[J].Plant Physiology,2008,147:1960-1975.
[37]Wu C,Fu Y P,Hu G C,et al. Isolation and characterization of a rice mutant with narrow and rolled leaves[J].Planta,2010,232:313-324.
[38]Fang L K,Zhao F M,Cong Y F,et al. Rolling-leaf14 is a 2OG-Fe(Ⅱ) oxygenase family protein that modulates rice leaf rolling by affecting secondary cell wall formation in leaves[J].Plant Biotechnol J,2012,10:524-532.
[39]夏妙林,唐冬英,楊远柱,等.水稻OsYABBY6基因参与叶片发育的初步研究[J].生命科学研究,2017,21(01):23-30.DOI:10.16605/j.cnki.1007-7847.2017.01.005
[40]Hibara K,Obara M,Hayashida E,et al. The ADAXIALIZED LEAF1 gene functions in leaf and embryonic pattern formation in rice [J]. Development Biology,2009,334(2):345-354.
[41]Li L,Shi Z Y,Li L,et al. Overexpression of ACL1 (abaxially curled leaf 1) increased nulliform cells and induced abaxial curling of leaf blades in rice[J].MolPlant,2010,3:807-817.
[42]Zhao S,Hu J,Guo L,Qian Q,Xue H. Rice leaf inclination2,a VIN3-like protein,regulates leaf angle through modulating cell division of the collar[J].Cell Research,2010,20:935-947..
[43]Zou L,Sun X,Zhang Z,Liu P,Wu J,Tian C,Qiu J,Lu T. Leaf Rolling Controlled by the Homeodomain Leucine Zipper Class IV Gene Roc5 in Rice[J].Plant Physiology,2011,156:1589-1602.
[44]Xu Y,Wang Y,Long Q,et al. Overexpression of OsZHD1,a zinc finger homeodomain class homeobox transcription factor,induce abaxially curled and drooping leaf in rice[J]. Planta,2014,239:803-816.
[45]Xiang J,Zhang G,Qian Q,et al. SEMI-ROLLED LEAF1 encodes a putative glycosylphosphatidylinositol -anchored protein and modulates rice leaf rolling by regulating the formation of bulliform cells[J].Plant Physiology,2012,159:1488-1500.
[46]Dai M,Zhao Y,Ma Q,et al. The rice YABBY1 gene is involved in the feedback regulation of gibberellin metabolism[J].Plant Physiology,2007,144:121-133.
[47]Itoh J,Hibara K,Sato Y,et al. Developmental role and auxin responsiveness of class III homeodomain leucine zipper gene family members in rice[J].Plant Physiology 2008,147:1960-1975.
[48]陳宗祥,潘学彪,胡俊.水稻卷叶性状与理想株形的关系[J].江苏农业研究,2001(04):88-91.
[49]彭桂峰,李义珍,扬高群.两系杂交稻培矮64S/E-32的超高产特性与栽培研究:超高产的决定因素[J].杂交水稻,2000,15(1):27-29.
(责编:张宏民)
猜你喜欢
军事文摘(2021年22期)2021-11-26
今日农业(2021年9期)2021-07-28
小聪仔(幼儿版)(2020年11期)2020-02-01
现代农业科技(2016年20期)2016-12-20
农家科技下旬刊(2016年9期)2016-12-15
西藏大学学报(自然科学版)(2015年1期)2015-12-25
西藏科技(2015年9期)2015-09-26
