
水稻粒长遗传及其功能基因研究进展
2021-04-23 04:34潘大建陈文丰孙炳蕊吕树伟江立群毛兴学范芝兰
广东农业科学 2021年3期
张 静,李 晨,潘大建,陈文丰,孙炳蕊,刘 清,吕树伟,江立群,毛兴学,范芝兰
(广东省农业科学院水稻研究所/广东省水稻育种新技术重点实验室,广东 广州 510640)
水稻(Oryza sativaL.)为人类最重要的粮食作物之一,其种植历史悠久、食用人口众多。目前,全世界约有30 多亿人口以稻米为主食。随着全球人口的不断增加、可耕作土地的逐步减少、生态环境及食品消费结构的变化,水稻生产呈刚性需求增长[1]。据预测,2030 年世界人口将达到80亿,届时全球对水稻产量的需求将比现在增加约40%[2]。随着人民生活水平的提高,稻米需求呈现多样化趋势,稻米品质也受到越来越高的重视。提高产量、改良稻米品质已是现代水稻育种的两个主要目标。
水稻粒型不仅是影响水稻产量的重要农艺性状,还与外观品质性状密切相关。粒长是水稻粒型的三大构成因素(粒长、粒宽、粒厚)之一。粒长、粒宽和粒厚对粒重的总影响力达94.4%,直接影响水稻单产,其中粒长对水稻粒重的贡献更大。研究表明,细长粒稻米通常表现较好的外观品质[3],且世界上大多地区(包括中国南方,美国,东南亚各国)的消费者更偏爱于长粒型的稻米。因此,改良水稻粒长成为一个非常重要的育种目标。著名的Basmati 系列香米、我国广东的丝苗品种都是长粒优质稻的典型代表,近年来大面积应用的优质不育系如野香A 和泰丰A 等也具有典型的长粒表型。试验表明粒长遗传受单基因、双基因、多基因或微效基因控制,但近年来国内外研究结果倾向粒长以多基因控制为主,属于数量遗传性状[4-5]。随着新一代测序技术的出现和功能基因组学的迅猛发展,有关水稻粒长的研究取得了较大进展,目前已经定位的与水稻粒长相关的QTLs 有120 多个,并克隆了一些控制粒长的基因。通过重测序定位及KASP 精细定位来定位水稻粒长基因,进而克隆该粒长基因,再通过基因编辑技术来改良籽粒性状,提高水稻产量,也已成为最新克隆基因的热点。本文就水稻粒长的影响因素、粒长与其他相关性状的关系、QTL 定位、重要粒长基因的克隆与功能分析以及粒长基因在分子育种上的应用等方面进行综述,以期为提高水稻产量和改善稻米品质提供参考。
1 水稻粒长的母体遗传
水稻籽粒由母代组织和子代部分两部分组成,前者包括颖壳和种皮,其中种皮由母体子房壁和珠被发育而来;后者包括胚和胚乳,经由双受精过程而发育。另外,籽粒生长可通过母体细胞质基因影响植株的光合呼吸作用,从而间接控制籽粒性状的表现。
在生长过程中,水稻谷粒离不开母体植株,有时可能还受母体生存状态影响[6],所以水稻粒形的遗传表达主要受母体加性和直接加性效应控制,且以母性加性效应为主。陈建国等[7]认为粒长的遗传基础较为复杂,胚乳核基因、母体植株核基因、细胞质基因对其均起作用,但以母体核基因的效应为主。通过粳型杂交稻的研究,林建荣等[8]发现稻米的糙米长性状主要受母体加性效应控制,且以母体遗传率为主。卢瑶等[9]对籼型杂交水稻的粒长研究表明,粒长性状主要受种子直接效应和母体效应的影响,以种子直接加性效应为主。
2 水稻粒长的遗传特点
粒长是粒型中最为重要的指标,最能反映籽粒形状。水稻粒长在不同品种中变化多样,其波动范围在6~15 mm。如典型的粳稻品种川7 的粒长仅有7.32 mm;而明恢63 是一个重要的籼稻品种,其粒长达到10.2 mm。关于水稻粒长的遗传特点因其研究材料不同所得结论也不尽相同,但近年研究表明,水稻粒长受多基因(主效基因或微效基因)调控。石春海等[10]以3 个粗短粒品种和3 个细长粒品种配制的3 个杂交组合为研究对象,结果发现粒长是以多基因为主的数量性状,在遗传效应中其加性效应占据的比重较大,拥有较高的广义遗传率和狭义遗传率。周清元等[11]对水稻粒长进行遗传分析发现,杂种F1代的籽粒平均长介于亲本之间,F2代中出现大的分离且呈连续分布,尺度测验结果表明粒长符合加性-显性模型,可见粒长的遗传受多基因体系控制。利用两个粒长上存在显著差异的籼稻品种为亲本构建重组自交系群体,对其粒长进行QTL 定位分析,贾小丽等[12]在探索水稻粒长的遗传机制中发现粒长性状在RILs 群体中呈连续变异,并表现出显著的超亲分离。
3 粒长与籽粒相关性状间的关系
3.1 水稻粒长与粒重的关系
粒重是水稻产量潜力的重要决定因素,也是水稻育种的主要目标之一[13],决定水稻的粒型等性状,而粒长是粒型的重要组成成分。许多研究表明,水稻粒长与粒重紧密相关。徐建龙等[14]对水稻籽粒性状进行遗传解析时发现粒重与粒长呈显著正相关。潘国庆等[15]以60份粳稻种质资源为材料,对粳稻粒型和粒重进行相关性分析,表明粳稻粒长的增加极显著提高了千粒重,且从相关系数上看,粒型对粒重的影响程度为粒长>粒厚>长宽比>粒宽。张颖慧等[16]以籼粳重组自交系群体为研究材料,将千粒重分为5组(≤25.0、25.1~30.0、30.1~35.0、35.1~40.0、≥40.1 g)进行粒型性状分析,结果发现粒长与粒重间存在极显著正相关,当粒重≤25.0 g时,粒重的增加主要依赖于增加粒长和粒厚,但随着粒重增加,粒长的作用逐渐减弱。李孝琼等[17]在对水稻粒型相关性状进行QTL定位时发现千粒重与粒长呈极显著正相关。通过对鄂晚10号/大粒王的F7代重组自交系群体进行千粒重和外观品质性状的考察,段丽丽等[18]发现粒重的增加与粒长呈极显著正相关,通径分析结果表明,粒宽对千粒重的作用最大,粒长次之。谢婷婷等[19]研究发现粒长与粒质量间的相关性达到显著水平。因此,水稻育种中改良粒型性状和粒重可以同步进行。
3.2 水稻粒长与品质的关系
稻米品质是指从稻谷生产到加工成直接消费品的全部过程中本身的物理及化学特性,是作为粮食或商品的综合反映。人们通常将其分为外观品质、碾米品质、蒸煮品质和营养品质4 类[20]。
粒型、垩白和透明度是评价稻米外观品质的3 个重要指标。粒型一般以粒长、粒宽及其比值表示,主要决定于粒长。根据米粒的长短情况将稻米分为长粒、中粒和短粒3 种。但由于地区和消费习惯差异,不同粒型的粒长绝对值并无统一标准。目前,大家较认可的是由Khush等[21]于1978 年提出的分类法,根据糙米的粒长和粒型将谷粒分为极长粒(>7.5 mm)、长粒(6.61~7.50 mm)、中粒(5.5~6.0 mm)和短粒(<5.50 mm)。垩白是指稻米胚乳中不透明的部分,垩白度和垩白率也是评价稻米外观品质的重要依据。我国优质稻谷国家标准规定:一级优质米的垩白率低于10%,垩白度低于1%[22]。此外,研究表明垩白属于复杂的数量性状,其形成受多种因素影响,除受灌浆期高温高湿条件和“库源”矛盾影响外,还与籽粒大小等性状密切相关。通过对36 个中籼杂交水稻组合的主要品质性状进行分析,表明在长米粒型、中米粒型组合中,垩白粒率和垩白度均随直链淀粉含量的增加呈显著或极显著增加,且在中米粒型组合中,垩白粒率和垩白度与米粒长宽比呈显著或极显著负相关[23]。Shomura 等[24]发现,qSW5基因在调控粒长增加的同时降低了稻米的垩白度和垩白粒率。Wang等[25]研究发现,GL7 位点拷贝数的变异导致了水稻粒长增加,进而降低稻米的垩白度和垩白粒率,从而使得产量提高、外观品质改善。
碾米品质是指稻谷在脱壳及碾磨后所保持的状态,主要用糙米率、精米率和整精米率来衡量,又称加工品质。碾米品质是粮食生产的外延,与稻米的商品价值密切相关。碾米品质除受气候环境因子、加工和收获因子等影响外,外观品质(粒长)对其也有重要影响。杨联松等[26]在对粒型与稻米品质间的相关性分析时发现,粒长与糙米率、精米率、整精米率间呈极显著负相关。刘燕德等[27]研究表明,粒长与稻米的碾米品质(糙米率、精米率和整精米率)间呈极显著负相关,即随着粒长增加,加工品质越差。相关系列研究也发现,整精米率与粒长呈极显著负相关[28-30]。在水稻加工过程中,应尽量减少暴晒,提高碾磨工艺水平。
蒸煮食味品质是指在蒸煮及食用过程中稻米所表现的理化及感官特性,如适口性(黏性、弹性、软硬度)、外观结构(颜色、光泽、饭粒完整性)、滋味、香味等,而直链淀粉含量、糊化温度、胶稠度是衡量蒸煮食味品质的直接指标。研究发现除三者之间密切相关外,外观品质也是影响蒸煮食味品质的重要因素。在对华南籼稻晚造稻米进行品质鉴定时,周少川等[31]研究表明,食味品质不仅与直链淀粉含量、胶稠度相关,还与粒型存在较大关联度,粒型细长、胶稠度较长的稻米具有较好的食味品质。罗玉坤等[32]对籼稻粒型和主要品质指标进行相关性分析表明,粒长与直链淀粉含量呈极显著负相关,与胶稠度呈显著正相关,即长粒型的品种往往蒸煮食味品质较好。
营养品质主要是指稻米中蛋白质、氨基酸及微量元素等含量和组成。许多研究发现,粒长与蛋白质含量、蛋白质指数、赖氨酸含量和微量元素间的相关性均达到显著水平。罗玉坤等[32]研究表明,粒长与蛋白质含量呈极显著负相关。张名位等[33]利用禾谷类作物种子数量性状遗传模型,对籼型黑米稻品种进行了分析,发现粒长与Fe、Mn 含量呈极显著正相关,而与Zn、P 含量呈极显著负相关。
综上所述可知,稻米的粒长与其品质性状密切相关。在对水稻后代选择中,通过粒长性状的间接选择,可达到改良稻米品质的育种目标。
4 水稻粒长相关QTLs 定位分析
粒长作为影响水稻粒重的一个重要成分,备受遗传学家关注。而相关基因的分子标记开发、定位和克隆也成为热门研究领域之一。随着分子生物学的快速发展,已有大量有关粒长的QTLs被报道且在水稻12 条染色体上均有分布,但控制粒长的主效QTL 主要集中在1、2、3 和7 号染色体,部分统计结果见表1。
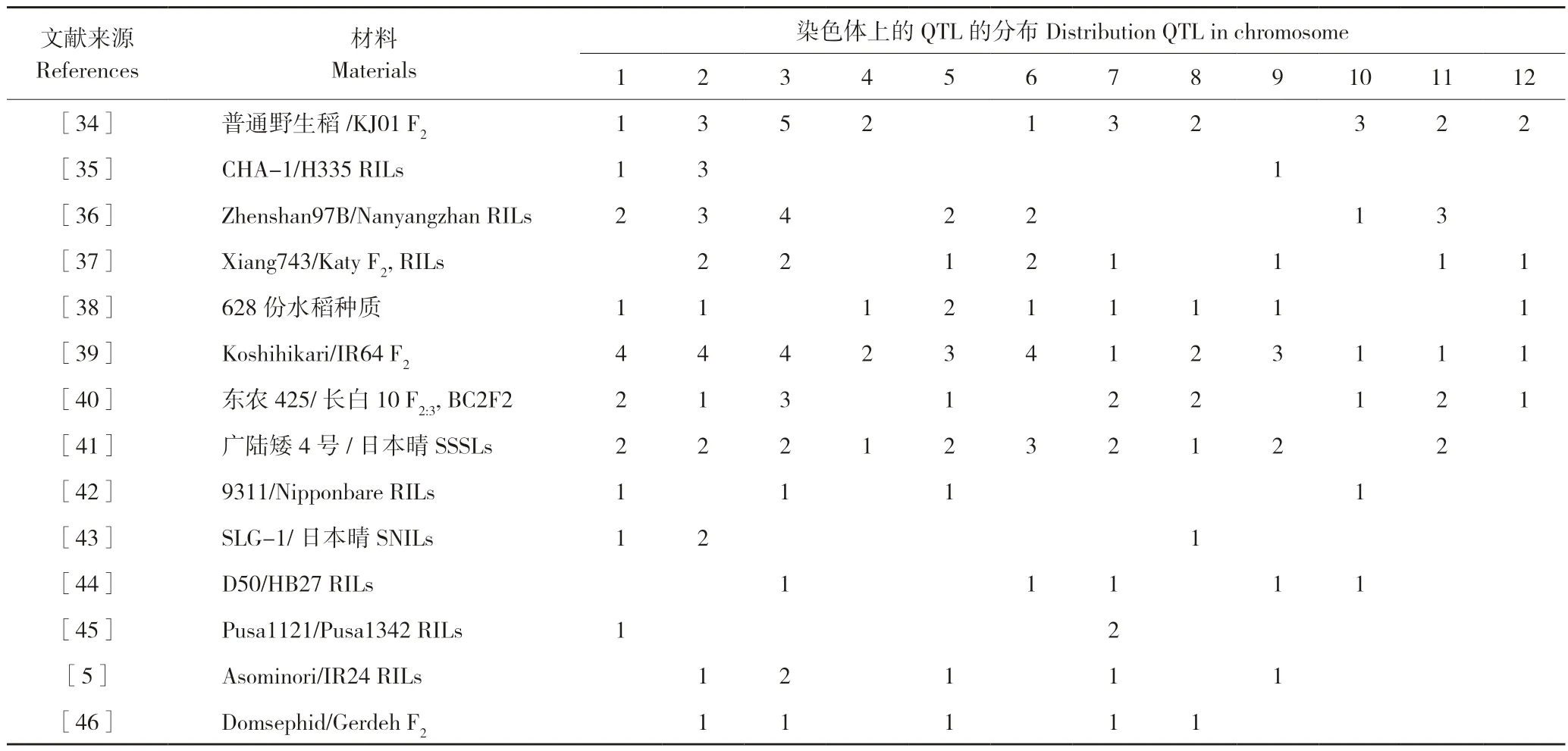
表1 部分水稻粒长QTLs 定位结果Table 1 Mapping results of QTLs for grain length in some rice varieties
5 水稻粒长主要QTLs 的克隆及功能
利用连锁和关联定位分析发现,许多与水稻粒长相关的QTLs 被鉴定,但只有少部分的QTLs被克隆(表2)。
GS3为第一个被克隆的负向调控水稻粒长、粒重的基因,位于3 号染色体上,测序结果发现大粒品种GS3第2 外显子中编码第55 位半胱氨酸的密码子(TGC)突变成终止密码子(TGA),造成蛋白翻译提前终止,缺失了178 个氨基酸,从而使得类PEBP 结构域缺少其他3 个功能域,最终导致粒长变长、粒重增加[47]。
qGL3/GL3.1为在水稻品种‘N411’中克隆的并位于3 号染色体上另一个控制粒长和粒重的主效QTL,它编码一个与拟南芥BSU1 同源且含有Kelch-like 重复结构域(OsPPKL1)的蛋白磷酸酶[48]。其稀有等位基因qgl3在第2 个Kelch结构域处的一个保守AVLDT 构件内发生天冬氨酸向谷氨酸的转变,从而导致长粒的表型[49]。研究表明,二者虽为同源蛋白,但qGL3对BR信号通路却起到负调控作用,且与拟南芥BR 信号通路不同,qGL3是通过影响OsGSK3 的蛋白水平、是否磷酸化和OsBZR1 的分布来影响BR 信号和对粒长进行调控的。而‘N411’粒长的增加则是由于qGL3的功能缺失使其对OsGSK3 不具备脱磷酸化功能[50]。
GL3.3为通过‘珍汕97’和‘南洋占’构建的RIL 检测到与粒长有关的主效QTL,最终精细定位在15.1 kb 区间。研究表明,GL3.3对粒长起负向调控作用,‘南洋占’中GL3.3等位基因的第三外显子的碱基突变使得可读框(ORF)变小最终产生了长粒[51]。
LGY3为籼稻‘R186’与泰国品种‘RD23’通过杂交及回交所得到的与粒长相关的基因并将其最终定位到8.9kb。LGY3编码蛋白OsMADS1,G 蛋白β γ亚基二聚体通过与OsMADS1 直接作用来影响其转录,进而对水稻粒型造成影响。以‘日本晴’为背景构建了NIL NPB-lgy3 发现lgy3 增加了籽粒纵向细胞的分裂才促进了粒长的增加。OsMADS1lgy3 是OsMADS1 发生变异进而编码的一种C 端截短蛋白,对粒长也起到促进作用[52]。
GL7为能够正向调控籽粒大小的基因,其编码一种和拟南芥LONGIFOLIA 蛋白同源的蛋白质,该蛋白调控颖花纵向细胞长度。而在GL7位点存在一个17.1 kb 的串联重复片段使得该基因上调表达,附近的负调控因子下调表达,最终导致粒长增加[53]。上调表达GW7促进了颖花纵向细胞的分裂,同时减缓了横向细胞分裂,致使形成细长的谷粒[54]。
GLW7为利用全基因组关联分析获得的主效基因,通过编码一个转录因子OsSPL13来正向地调控水稻的粒长和产量,研究发现OsSPL13的5′UTR 区一个串联重复序列的改变导致基因表达水平的差异,该基因的高表达促进籽粒增大[55]。
qTGW3为位于3 号染色体上另一个控制粒长和粒重的主效QTL,通过编码GLYCOGEN SYNTHASE KINASE3 家族的成员OsSK41,负向调控水稻籽粒长度和重量。OsSK41 与AUXIN 相互作用,使其磷酸化,形成OsARF4。OsSK41 与OsARF4 共表达使OsARF4 在水稻原生质体中积累增加,从而抑制一组下游控制水稻籽粒发育过程的基因(包括一些生长素的响应基因)表达,进而导致粒形改变。OsARF4 功能的丧失导致大粒表型[56]。
Gnp4被定位在4号染色体上一个10.7 kb的区间内,并发现3 个蛋白(LAX1、OsIAA3和OsIAA17)与其互作,而Gnp4与OsIAA3 或OsIAA17 互作可以调控水稻籽粒长度;在过表达Gnp4植株中,提高自由生长素含量,可以激活ARF 家族转录因子下游基因的调控,从而调控水稻籽粒长度过量表达,显著增加粒长[57]。
GS2/GL2编码一个生长调控因子(OsGRF4),当该基因第3 外显子发生稀有突变(TC487-488AA)时,影响了miR396c 对OsGRF4的剪切,进而导致GS2/GL2的过表达,使细胞变大并产生大粒的表型[58-59]。TGW6是一个调控水稻粒重的基因,该基因的突变或者表达水平的降低导致粒长增加,但不影响粒宽和粒厚,进而增加粒重[60]。
另外,还有一些水稻粒长基因与植物激素密切相关。如,D1是第一个被克隆的起主效作用的基因,编码G 蛋白亚基并参与G 蛋白介导的GA 信号传导,当该基因发生突变(外显子缺失833 bp),其突变体植株的籽粒变小,植株变矮[61]。BG1为水稻大粒基因,编码一种受生长素诱导的未知功能蛋白,过表达该基因使得生长素的极性运输能力显著提高,进而增大籽粒、提高产量[62]。D61编码BR 信号受体激酶OsBRI1,该蛋白功能的缺失导致细胞分裂、延伸受限,使得籽粒变短[63]。SRS5 编码α 微管蛋白,突变后,颖壳细胞长度降低使种子变小[64]。水稻的D2、D11、BRD1、BRD2和OsDWARF4等基因也均参与BR 合成,其突变体也具有粒长变短的特征[65-69]。OsGSK2负向调节参与调控下游BR 应答基因的表达,过表达该基因籽粒表现缩短[70],而BU1也参与BR 下游信号的传导,过表达该基因籽粒伸长[71]。此外,XIAO、SG1和SMG1等与BR 相关的基因也被分离且突变体的籽粒表现粒长变短[72-74]。
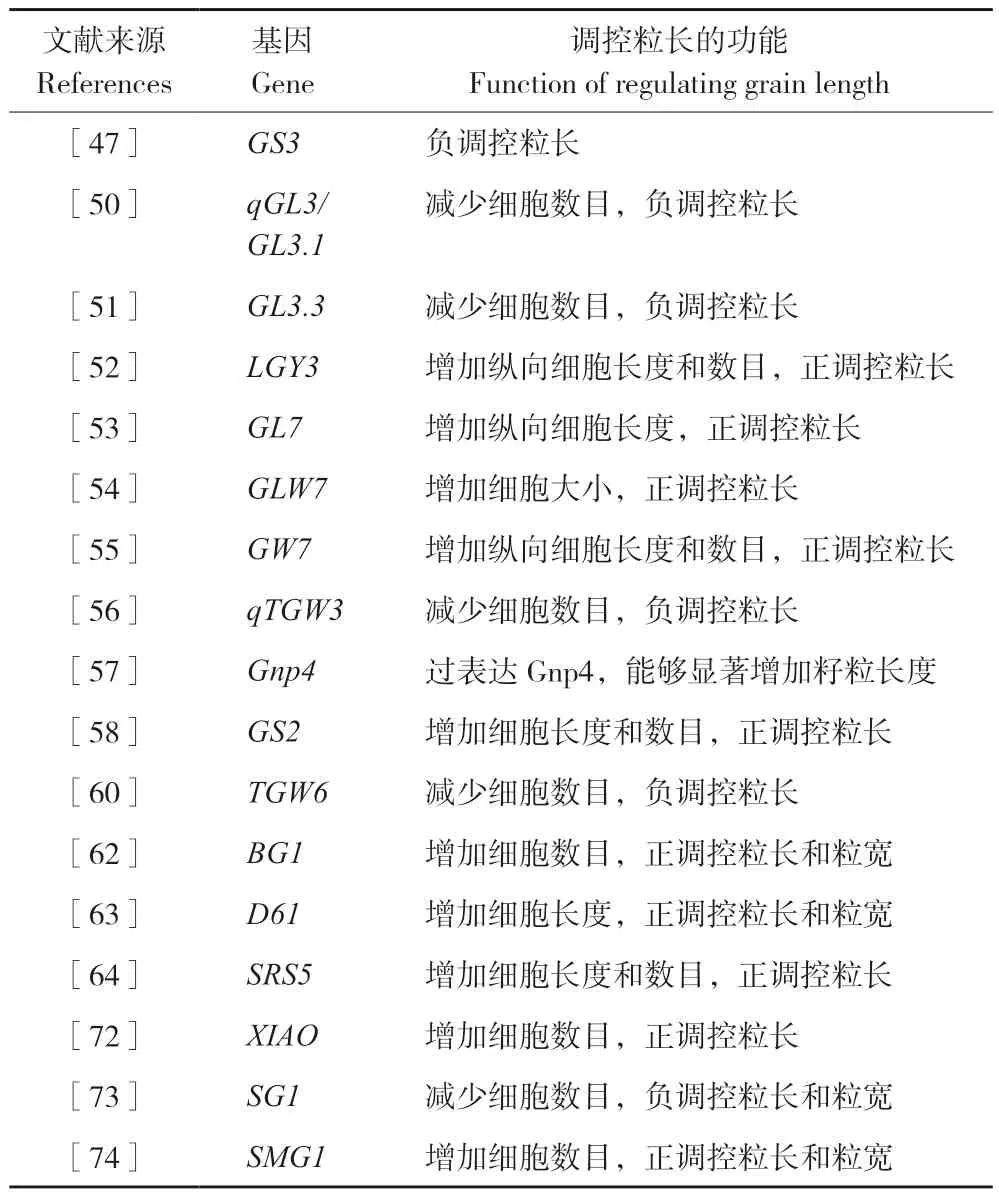
表2 调控水稻粒长的功能基因Table 2 Functional genes regulating rice grain length
6 粒长重要功能基因的应用
近年来,随着人们生活水平提高,对优质米的需求也日趋增加。如前综述,粒长不仅影响粒重性状,还与品质性状密切相关。因此,通过改良粒型来培育高产、优质新品种,不失为一条重要且有效途径。已克隆的一批控制水稻粒长的重要基因,既有助于揭示水稻产量性状复杂的遗传机制,同时也为水稻分子标记的辅助选择提供了理论依据和技术基础。分子标记辅助选择技术对现代分子育种十分重要。通过该技术结合田间杂交和回交,可以快速有效地达到转移基因和聚合多个基因。这不仅降低了育种成本,更缩短了育种时间,育种家能更好更快地选育出目标品种。GS3为控制粒长的主效基因,在自然界中存在广泛变异,该基因第2个外显子中C-A 的突变导致长粒发生,在此基础上,功能标记SF28 被筛选出[75-76]。利用该标记进行粒型改良育种的辅助选择,卓有成效。将GW8-GW7分子模块导入我国高产水稻品种后,在保证产量不降低的前提下稻米品质显著提升[55]。研究还表明,水稻品种泰丰优55 和泰丰优208 在聚合了GW7和GS3基因的优异等位变异后,其稻米产量和品质均明显提高。王军等[77]根据TGW6与其等位基因在功能区域的缺失,开发并筛选出该基因的功能标记CAPs6-1,从而快速筛选出携带TGW6的水稻品种。
总体来看,关于水稻粒长主效QTLs 的克隆还较少且其功能基因组成果在育种上利用还不够,新基因的挖掘还需不断深入,以期克隆更多的粒长基因来改良并丰富水稻种质资源,供遗传学家和育种学家加快水稻育种进程;功能基因的利用还需不断加强,对已经克隆的粒长基因的作用机制及遗传调控网络进行更深层次的研究,以期为两个及以上基因的聚合提供更加科学的理论依据;利用高效基因组编辑CRISPR 技术有效的将有利等位基因进行聚合,对粒长的负调控基因(GS3、GL3.1、SG1等)进行编辑,定向敲除,对品种中的个别不良性状进行精准改良,快速获得新材料,提高育种效率。但在基因编辑过程中粒长基因也会对水稻其他农艺性状造成影响,如何有效避免粒型基因对其他农艺性状造成的消极作用还需进一步研究。
猜你喜欢
麦类作物学报(2022年6期)2022-08-09
中国农学通报(2022年13期)2022-05-31
美食(2022年5期)2022-05-07
早期教育(家庭教育)(2021年11期)2021-12-17
少儿科学周刊·儿童版(2021年21期)2021-12-11
现代畜牧科技(2021年4期)2021-12-05
辽宁农业科学(2021年1期)2021-03-17
现代农业研究(2019年3期)2019-06-05
吉林农业(2017年7期)2017-07-12
科学种养(2017年6期)2017-06-13
