
基于转录组测序解析甜玉米耐低温发芽的相关基因
2021-04-23 04:34李光玉肖颖妮胡建广李春艳李余良卢文佳李高科刘建华
广东农业科学 2021年3期
李 武,李光玉,肖颖妮,胡建广,李春艳,李余良,卢文佳,李高科,刘建华
(广东省农业科学院作物研究所/广东省农作物遗传改良重点实验室,广东 广州 510640)
【研究意义】甜玉米是一类果蔬兼用的特用型玉米,其形成是由于淀粉合成代谢途径发生基因突变,导致淀粉合成途径受阻、蔗糖等糖类物质积累。由于甜玉米是普通玉米籽粒淀粉合成基因突变产生,其种子活力普遍较低,这成为制约我国甜玉米生产的主要因素之一。低温发芽率是甜玉米种子活力鉴定的重要指标,但目前有关玉米应对低温胁迫的研究尚不充分[1]。低温胁迫主要分为冷害胁迫(<15 ℃)和冻害胁迫(<0 ℃)。作为C4 植物代表的玉米,当温度低于15 ℃或者更低时,容易遭受冷害胁迫而影响发芽率[2]。伴随全球气候变化,极端天气频发,广东近5 年来的春季最低气温维持在4~5 ℃左右,平均气温约13 ℃,玉米直播常遭遇低温胁迫,不耐低温发芽的品种往往容易受灾。因此,低温发芽特性好的甜玉米品种对推广直播栽培、实现节本增效具有重要价值[3]。挖掘鉴定低温发芽相关调控基因,将为阐明甜玉米低温发芽的生物学基础、加快育种改良提供重要的理论支撑。
【前人研究进展】玉米耐冷性遗传研究与功能鉴定主要集中在发芽期和苗期。早期研究中,Mc Connell 等[4]研究6 个耐冷性差异显著的自交系及其F1、F2和回交群体,发现苗期活力受加性和显性效应控制,而发芽能力受加性、显性效应和上位性基因调控。Hodges 等[5]研究由4 个耐冷能力不同的自交系完全双列杂交得到的群体,结果表明不同群体在发芽期和苗期耐冷性的遗传因素可能不同。Revilla 等[6]利用同样的方法研究5个自交系及其后代F1的耐冷性,发现高发芽率和苗期快速生长两个耐冷性状的遗传调控符合加-显性模型。近年来,较多学者利用不同的作图群体对玉米耐低温发芽和苗期耐冷的数量性状遗传位点(Quantitative trait locus,QTL)进行鉴定。例如,利用不同作图群体(F2:4家系、重组自交系群体等)研究玉米耐冷相关QTL,多集中于分析发芽期发芽和苗期光合作用性状,如发芽指数、光合作用效率、根干重、茎含氮量等指标[7]。有学者利用ETH-DH7×ETH-DL3 构建的F2:3群体,于第6号染色体定位到低温下控制苗期光合作用的主效QTL,且该位点能在多个环境下被重复检测[8]。随后Nguyen 等[9]利用相同的F2:3群体研究低温下苗期光合系统耐冷性,但检测到的主效QTL 位于第3 号染色体上。Rodriguez[10]利用210 份F2:3家系(EP42×A661),以茎干重、存活率等为评价指标,在低温环境下检测到4 个QTL(其中1个为存活率QTL)。Hu 等[11]利用重组自交系群体,以相对出芽率、初生根长为指标,通过不同温度时间处理(12 ℃16 h、18 ℃8 h、28 ℃24 h)鉴定出15 个QTL,主要分布在第4、5、6、7、9号染色体上。李旭辉等[12]利用低温萌发能力高的玉米自交系220 和P9-10,以及低温出苗能力弱的自交系Y1518 和PH4CV,配制了3 个F2:3群体,通过6K 玉米SNP 芯片做基因型鉴定,确定了67 个与低温发芽相关的QTL,其中17 个贡献率大于10%。上述研究报道,均以普通玉米种子为分析对象,鲜有甜玉米种子低温萌发相关的分子遗传学研究。
【本研究切入点】 根据前期工作对甜玉米种子低温下的发芽率、发芽势、发芽指数、活力指数等指标的综合评价,我们获得了2 份低温萌发能力差异显著的甜玉米自交系Scil-H7th 和Scil-C4th。【拟解决的关键问题】利用低温萌发能力高的Scil-H7th 为供体亲本(DP)、低温萌发能力低的Scil-C4th 为轮回亲本(RP),杂交构建近等基因系(Near Iso-genic Lines,NIL),并分析不同品系的低温萌发能力。通过转录组测序分析低温萌发能力不同的材料,以鉴定甜玉米耐低温萌发的相关基因,为后续甜玉米耐低温萌发特异种质资源的创制奠定基础。
1 材料与方法
1.1 试验材料
甜玉米自交系Scil-C4th(轮回亲本,RP)和Scil-H7th(供体亲本,DP)于2014 年早季在广东省农业科学院白云试验基地种植并杂交获得F1;以Scil-C4th 为轮回亲本连续回交4 次,经过4 代自交得到稳定自交系,并通过种子标准发芽试验进行表型选择,最终获得3 个RP 背景的NIL品系,分别命名为L6、L18 及L41。筛选完整饱满的Scil-C4th、Scil-H7th 及NIL 品系甜玉米种子作为本试验材料。
1.2 试验方法
1.2.1 种子发芽处理 将Scil-C4th、Scil-H7th 及NIL 不同品系甜玉米种子浸泡12 h 后进行种子发芽试验,分别在15 ℃人工气候箱中培养,每个发芽处理3 次重复,每个重复100 粒种子。样品在12 h 光照/12 h 黑暗环境中生长,光照强度为250 μmol/m2·s,持续7 d[13]。在吸胀后1 d(时期Ⅰ)和吸胀后3 d(时期Ⅱ),从样本中随机收集5 个种子胚,液氮冻存,用于提取RNA 进行高通量测序。发芽试验期间,每天计数发芽种子,并在吸胀后7 d 测量根长、茎高以及计算发芽率[14],在80 ℃干燥24 h 后测定幼苗干重。计算发芽指数(GI)和种子活力指数(VI):
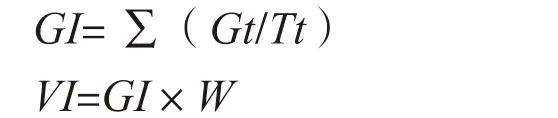
式中,Gt为第t天发芽种子的数量,Tt为对应于Gt的时间(d),W为幼苗干重。
1.2.2 RNA 提取、文库制备及测序 将种子胚用液氮匀浆,用植物核糖核酸提取试剂盒(中国天根生物技术有限公司,北京)提取总RNA。通过在1%琼脂糖凝胶上电泳分析RNA 质量,并用NanoDrop 2000(赛默菲舍科学公司,美国)鉴定RNA 的纯度和浓度[15]。随后,每个样品取1.5 μg总RNA,按照HiSeq 2500 测序仪(Illumina 公司,美国)说明书进行cDNA 文库构建、质量控制和高通量测序。
1.2.3 数据拼装、功能注释与分类 对测序产生的原始数据进行过滤,删除包含poly-N 和低质量的reads,以获得干净的reads。然后使用bowtie2[16]或Tophat[17]将干净的reads 比对到参考基因组。每个基因的FPKM 值使用Cufflinks 计算[16],每个基因的reads 数通过HTseq-coun 计算[18]。P<0.05 被设置为显著差异表达的阈值。对筛选出来的差异表达基因(Differentially expressed genes,DEGs)进行基于R 软件包的GO 富集和KEGG 通路富集分析。
1.2.4 qRT-PCR 验证候选基因表达量 采用qRT-PCR 技术从转录组DEGs 中选择7 个候选基因表达量进行验证[15]。基因序列信息从Ensembl Plants(http://plants.ensembl.org)获得,引物采用Primer3 设计(https://bioinfo.ut.ee/primer3-0.4.0/)。使用cDNA 第一条链反转录超级试剂盒反转录获得样本的第一链cDNA,并根据AceQ qPCR 混合试剂盒操作说明对反转录获得的cDNA 进行qRTPCR验证。本试验采用Step-One-Plus系统(Applied Biosystems 公司,美国)进行测定,每个基因采用3 个生物学重复和3 个技术重复测定表达水平[15]。Actin基因被用作获得Ct 值标准化的内源性参照基因,通过比较Ct(ΔΔCt)方法计算基因的相对表达值。
2 结果与分析
2.1 甜玉米品系DP、RP 及3 个NIL 的发芽性能
为明确不同品系的低温萌发能力,测定自交系Scil-H7th、Scil-C4th 及3 个NIL 的发芽相关指标。由图1 可知,DP(Scil-H7th)的种子发芽和成苗显著快于RP(Scil-C4th);而衍生自DP 和RP 的NIL 中,L18 表现出与DP 相似的成苗特性(图1A)。相比随机选择的另外两个NIL(L41和L6),L18 表现出种子发芽快和种子活力高的特点;相比亲本,L18 在发芽7 d 后发芽率达到64.54%,显著高于RP 的23.58%,其发芽势为RP 的1.20 倍,接近DP(图1B)。由以上指标分析可知,L18 的发芽指数和种子活力指数均优于RP。
2.2 甜玉米品系DP、RP 和L18 的RNA 测序文库构建、转录组测序及聚类分析
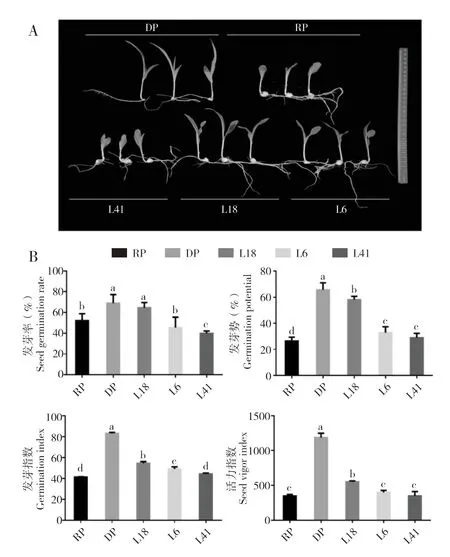
图1 甜玉米品系DP、RP 及3 个NIL 在低温下的萌发特性Fig.1 Germination characteristics of sweet corn lines DP,RP and three NILs at low temperature
为了筛选与甜玉米低温萌发的相关基因,利用高通量测序对NIL L18 及其亲本进行转录组分析。RP、DP 和L18 种子经低温催芽,在吸胀后1 d(时期Ⅰ,以-1 表示)和吸胀后3 d(时期Ⅱ,以-2 表示)取样,得到RP-1、DP-1、L18-1 和RP-2、DP-2、L18-2 样品,提取RNA 进行转录组测序,每个时期3 次重复,共测序获得18 个玉米样品平均8.54 Gb 的原始碱基,经质量控制过滤后的干净碱基平均为8.14 Gb,Q30 值介于91.71%~92.44%之间。
测序原始数据过滤后,总共51809565~55432742 个reads 用于比对分析。结果(表1)显示,能够比对到参考基因组的reads 比例在72.92%~74.84%之间。对于RP、DP 和L18,具有唯一匹配的reads 比例分别为70.57%、70.45%和70.46%。利用FPKM 值(每百万份映射读数转录本的每千碱基片段)进行定量分析,基因表达量分布如图2A 所示,结果表明各样品的基因表达水平分布范围较为一致;基于相似性和主成分分析对每个样本进行区分和聚类,结果(图2 B和图2 C)表明重复样品间相似度高,重复性好。
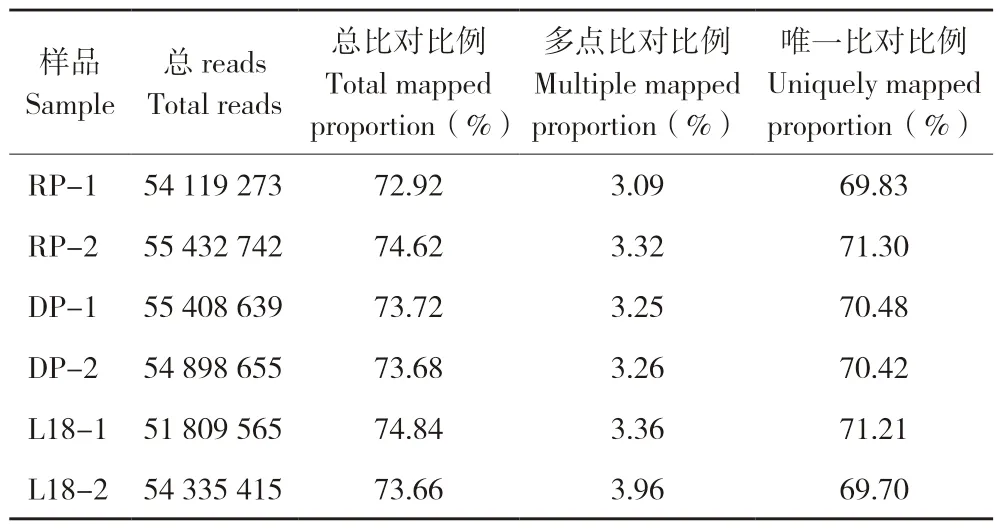
表1 甜玉米品系DP、RP 和L18 的转录组测序数据Table 1 Transcriptome sequencing data of sweet corn lines of DP,RP and L18
2.3 甜玉米品系DP、RP 和L18 在不同萌发阶段的差异表达基因
为了筛选与低温萌发相关的基因,对DP、RP 和L18 在不同萌发时期以及同一萌发时期3 个品系之间进行比较分析。自交系内不同萌发时期的分析结果(图3A)表明,DP-2 和DP-1 共有1629 个DEGs,其中780 个基因表达上调,849个基因表达下调;RP-2 和RP-1 共有2834 个DEGs,其中731 个基因上调表达,2103 个基因表达下调;L18-2 和L18-1 共有1823 个DEGs,其中1002 个基因表达上调,821 个基因表达下调;3 个材料在两个萌发时期共有的DEGs 为447 个,其中RP 和L18 共有的DEGs 有986 个,高于L18与DP 之间共有的DEGs(630 个)。分析DP 和RP、L18 与RP 之间相同萌发时期的DEGs,结果(图3 B)发现,DP 与RP 在两个萌发时期共有的DEGs 达到了3484 个,而L18 与RP 共有的DEGs 只有627 个,说明L18 遗传背景已与RP 较接近;取两者共有基因的交集,获得410 个共有DEGs,推定为低温萌发过程中不同材料间表达差异的基因集。
2.4 功能注释、通路富集和低温萌发相关基因筛选

图2 RNA 测序分析得到甜玉米品系DP、RP 和L18 的基因表达概况和聚类分析结果Fig.2 Overview of gene expression of sweet corn lines of DP,RP and L18 and clustering analysis result base on RNA sequencing
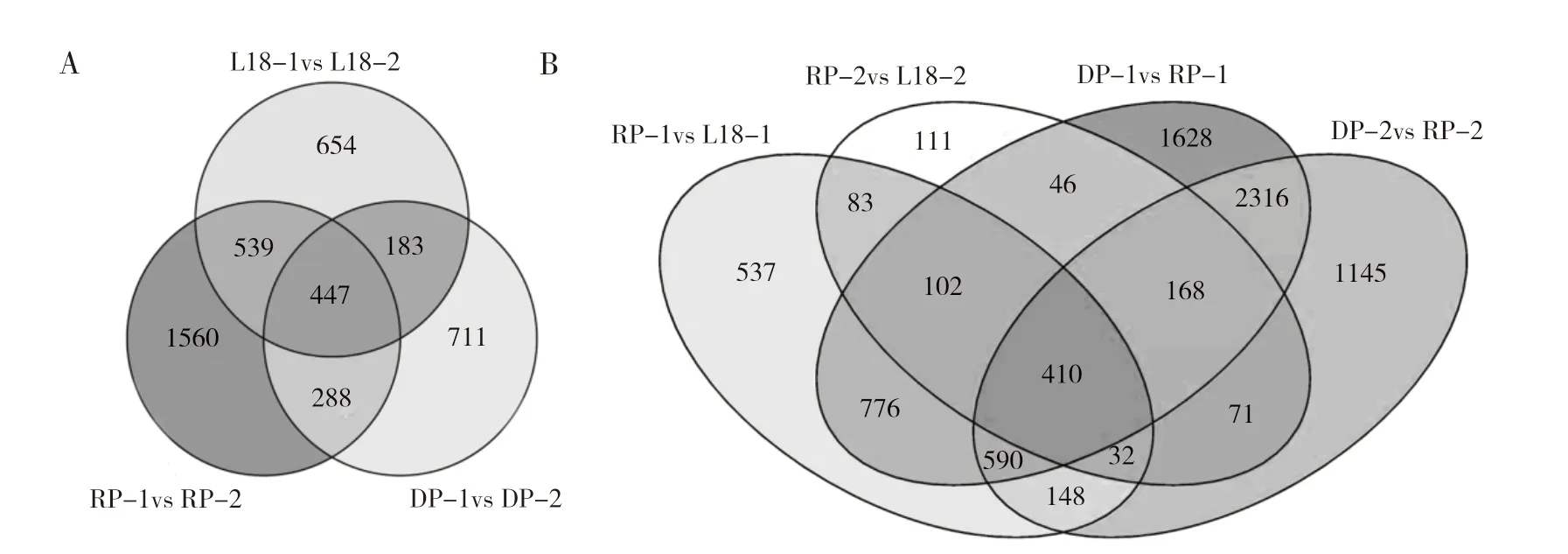
图3 甜玉米品系DP、RP 和L18 在低温萌发过程中的DEGs 韦维恩图Fig.3 Venn diagram of DEGs in sweet corn of DP,RP and L18 during low-temperature germination
为了进一步分析DEGs 所在的通路,采用KEEG 数据库对上述DEGs 进行功能注释和通路富集分析。结果显示,共有89 个和43 个涉及的通路被注释在RP-1 与L18-1 组和RP-2 与L18-2组;在3 个品系中同时鉴定了6 个关键通路,包括zma04712 昼夜节律-植物、zma01040 不饱和脂肪酸生物合成、zma00945 芪类化合物、二芳基庚酸和姜酚生物合成、zma00941 类黄酮生物合成、zma00940苯丙酸生物合成和zma00195光合作用。4个比较组(RP-1 vs DP-1、RP-2 vs DP-2、RP-1 vs L18-1、RP-2 vs L18-2)共有的410个DEGs已完成功能注释,其中286个基因被鉴定为功能性蛋白质,124个基因为未鉴定蛋白。Zm00001d005672、Zm00001d015497(线粒体乌头酸水合酶3)、Zm00001d033044(GDSL酯酶/脂肪酶)、Zm00001d033307(肌动蛋白轻链1)、Zm00001d033909、Zm00001d033948(Tubbylike F-box蛋白2)和Zm00001d037001(无静脉样蛋白)显示具有超过100倍的上调表达,Zm00001d029587(锌指蛋白AZF2)是L18中发现的下调超过100倍的蛋白。进一步分析发现,65个DEGs仅在DP和L18中表达,而不在RP中表达,其中32个基因在数据库中被注释。结合自交系在不同萌发时期的DEGs和自交系间相同萌发时期的DEGs,筛选出有交集的20个DEGs(表2),推测这些基因与低温萌发密切相关。

表2 20 个与甜玉米低温萌发密切相关的DEGs 及功能注释Table 2 Annotation of 20 DEGs highly related to low-temperature germination of sweet corn
根据注释信息进行分析,进一步筛选出最有可能参与低温萌发过程的7 个上调基因(Zm00001d011006、Zm00001d018738、Z m 00001 d 028815、Z m 00001 d 020652、Z m 00001 d 037383、Z m 00001 d 029696、Zm00001d018195),利用qRT-PCR 对DP 在低温萌发过程2 个时期的胚部、根系和嫩叶进行表达水平检测。结果表明,这些基因表达量变化趋势与转录组测序结果中的变化趋势趋于一致(图4 A)。其中,Zm00001d037383(UGT1基因)随发芽进程表达显著上调,并且在胚部的表达显著高于在其他组织的表达(图4 B)。
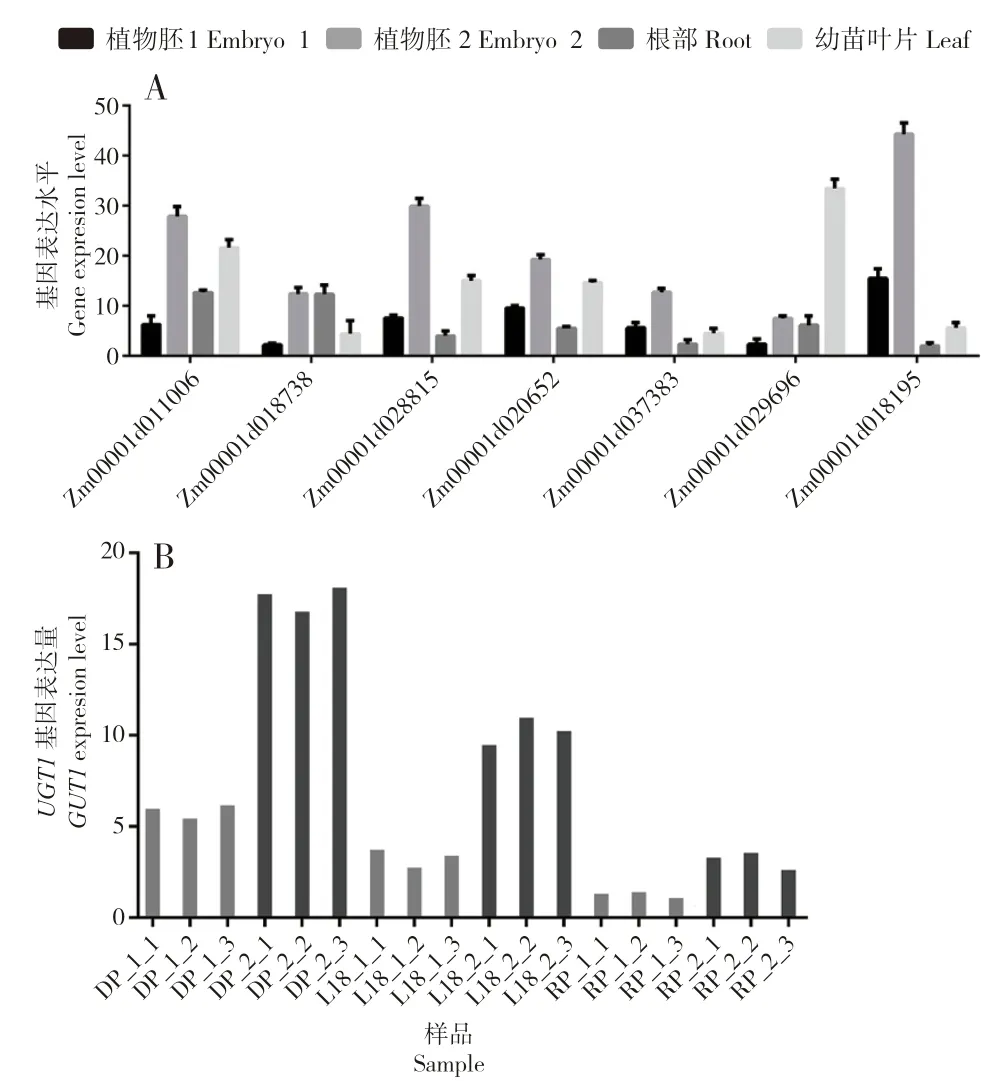
图4 甜玉米品系DP、RP 和L18 低温萌发相关基因的qRT-PCR 分析Fig.4 qRT-PCR analysis of genes related to low-temperature germination of sweet corn lines of DP,RP and L18
3 讨论
已有不少报道指出,通过构建近等基因系研究玉米生长发育和生理过程的潜在分子机制[19-20]。本研究通过将高种子活力甜玉米材料Scil-H7 的染色体片段导入低种子活力材料Scil-C4th 的遗传背景,获得了不同种子活力的近等基因系。其中,L18 的种子发芽率比RP 高23.58%,发芽势是RP 的1.20 倍,显著提高发芽指数和种子活力指数。生理实验结果表明,L18很可能携带了有利于低温条件下种子活力和发芽能力的关键基因位点,这为玉米低温萌发相关基础研究提供了有价值的种质资源。
通过Illumina 测序获得Scil-H7、Scil-C4th和L18 甜玉米胚在两个发芽阶段平均产生的8.54 Gb 原始序列数据。甜玉米种子发芽期间已建成了复杂的信号和代谢途径[21-22],影响种子发芽的因素包括不同途径的变化[23]。转录组分析表明,发芽过程中全基因组基因表达发生广泛的变化[24]。许多已鉴定的DEGs 编码蛋白质在代谢过程和核苷酸/蛋白质结合中的功能已经得到证明。KEGG 富集显示,本研究在3 个品系中同时鉴定了6 个关键通路,包括zma04712 昼夜节律-植物、zma01040 不饱和脂肪酸生物合成、zma00945 芪类化合物、二芳基庚酸和姜酚生物合成、zma00941 类黄酮生物合成、zma00940 苯丙酸生物合成和zma00195 光合作用。途径分析表明,这些DEGs 主要与次级代谢和种子发芽活力相关。转录组分析一共鉴定出447 个DEGs 存在于不同发芽阶段的Scil-H7、Scil-C4th 和L18 中,它们可能建立了一个调控网络,并随胚发芽进程而变化。
基于L18 和轮回亲本Scil1-C4th 之间相似的遗传性状,通过高通量转录组测序的方式捕获与种子活力相关的DEGs 作为候选基因。将Scil-H7 设为参照,在L18、Scil-H7 和Scil1-C4th的两个发芽阶段比较鉴定出410 个DEGs,其中65 个仅在Scil-H7 和L18 中表达,而不在Scil1-C4th 中表达,这些 DEGs 被认为是与种子发芽紧密关联的调节因子,可用于后续功能分析。通过对不同组织样本的qRT-PCR 验证,发现Zm00001d037383(UGT1基因)与低温发芽高度相关。
对UGT1基因进行功能注释与同源基因功能搜索,发现在拟南芥中UGT1 可以催化单螺旋藻,促进葡萄糖结合;UGT1 的敲除导致木质素单体生物合成,木质素聚合和细胞壁相关转录因子表达水平大幅提高,表明UGT1 催化的单螺旋葡萄糖基化对正常细胞壁木质素是必需的,从而在一定程度上阐明了细胞壁发育和细胞壁木质化的分子机制[25]。然而,细胞壁的发育是植物种子早期发芽的重要特征,细胞壁相关基因表达的活跃对植物种子更强的抵抗逆境胁迫能力、快速发芽与出苗能力具有重要意义。因此,推测UGT1基因可能是Scil-H7 中响应低温发芽的重要目标基因。分析UGT1基因在转录组测序样品中的表达量发现,该基因在玉米种子低温发芽过程中表达量迅速增加,表现为低温胁迫下的迅速响应。UGT1基因在玉米中耐低温发芽的作用及其分子机制有待进一步探索。
4 结论
本研究利用高通量转录组测序技术,对耐低温发芽差异的母本及其自交系后代进行分析,从不同材料在相同时期的4 个比较组(RP-1 vs DP-1、RP-2 vs DP-2、RP-1 vs L18-1、RP-2 vs L18-2)鉴定出共有DEGs 410 个。其中,65 个DEGs 仅在Scil-H7 和L18 中特异表达,32 个基因功能注释完整,从中进一步筛选出20 个可能与低温萌发密切相关的基因。对其中最有可能参与低温萌发的7 个上调表达基因进行表达量验证,其中编码UDP-葡萄糖基转移酶的基因Zm00001d037383(UGT1)在DP 胚部表达明显高于其他组织,推测该基因在甜玉米耐低温萌发过程中起关键作用。研究结果,为后续创制低温萌发能力更强的甜玉米种质资源奠定了基础。
猜你喜欢
中国种业(2022年8期)2022-08-19
今日农业(2022年6期)2022-07-05
湖南农业科学(2022年1期)2022-06-06
江苏农业科学(2022年6期)2022-04-15
北京农学院学报(2022年1期)2022-03-10
广西蚕业(2021年3期)2021-10-28
当代水产(2021年1期)2021-03-19
科学导报(2020年8期)2020-03-12
安徽农学通报(2017年24期)2018-01-12
农民致富之友(2016年5期)2016-10-21
