
小球藻蛋白/海藻酸钠互穿纤维的制备与特性
2021-05-08 08:41
食品工业 2021年4期
1.华东理工大学食品科学与工程系,生物反应器工程国家重点实验室(上海 200237);2.中国农业大学信息与电气工程学院(北京 100083)
小球藻(Chlorella)是一种单细胞绿藻,也是首先实现商业化利用的微藻之一[1]。小球藻不仅以胶囊、片剂、口服液等形式作为保健品销售,同时也被广泛应用于饮料、面条、饼干之中[2]。小球藻中含有大量优质蛋白质,占小球藻粉的40%~70%,其氨基酸组成超过FDA(食品及药物管理局)颁布的用于人类食品的蛋白质标准,研究证明小球藻蛋白及其水解物具有良好的抗氧化能力[3-5]。然而关于海藻的科学研究及商业化应用多集中于海藻多糖,对于海藻蛋白质的研究有限[6],限制了小球藻蛋白的应用领域。
互穿网络结构(IPN)是由2个或2个以上的聚合物网络组成的聚合物结构,这些聚合物网络在分子层面上相互交错穿插,2种聚合物之间并无共价键连接[7]。如果聚合物体系中只有一种聚合物交联形成网络,所得结构称为半互穿网络结构(semi-IPN)[7]。与单一聚合物形成的结构相比,IPN结构及semi-IPN结构使产品在机械强度、热稳定性及吸水能力等多个方面得到改善[8-10]。海藻酸钠是一种从褐藻中提取出的海藻多糖,是由β-D-甘露糖醛酸和α-L-古罗糖醛酸组成的线性聚合物,在Ca2+等二价阳离子作用下可迅速形成具有“蛋壳结构”的凝胶[11-13],常用于构建互穿网络结构。对于IPN结构的研究主要集中在组分添加量对于产品性能的影响,对于组分交联度的研究较少。
试验以小球藻蛋白和海藻酸钠为原料,使用谷氨酰胺转胺酶(TG酶)和氯化钙分别交联两种原料形成网络结构,通过湿法纺纱制备具有互穿网络结构的凝胶纤维,并探究TG酶添加量对于凝胶纤维的结构、物化特性及降解过程的影响。通过傅里叶红外光谱(FT-IR)及小球藻蛋白交联度的测定验证制备凝胶纤维的可行性,进而对凝胶纤维中小球藻蛋白的二级结构、凝胶纤维的吸水能力与持水能力进行评价。探究凝胶纤维在蛋白酶作用下的降解过程,并对小球藻蛋白及其酶解物的释放过程进行分析。试验首次将小球藻应用于凝胶纤维制作,可为拓展小球藻蛋白的应用领域提供理论基础。
1 材料与方法
1.1 材料与试剂
海藻酸钠(南京都莱生物技术有限公司);破壁小球藻粉(蛋白质含量78%,青岛科海生物有限公司);谷氨酰胺转胺酶(2 100 U/g,酷尔化学科技(北京)有限公司);碱性蛋白酶(200 000 U/g,上海腾骞生物科技有限公司);其他常用化学试剂均为分析纯。
1.2 仪器与设备
CP213电子分析天平(奥豪斯仪器(上海)有限公司);HHS-I1-1电热水浴锅(上海一恒科学仪器有限公司);SHA-BA恒温振荡器(常州朗越仪器制造有限公司);DL-5-B离心机(上海安亭科学仪器厂);STARTER2100 pH计(上海华联医疗器械有限公司);MOV-212F鼓风干燥箱(上海天呈实验仪器制造有限公司);UV-2000紫外分光光度计(尤尼克(上海)仪器有限公司);FTIR5700傅里叶红外光谱仪(尤尼克(上海)仪器有限公司)。
1.3 方法
1.3.1 小球藻蛋白/海藻酸钠互穿结构凝胶纤维制备
采用Sibaja等[14]的方法并稍作修改,通过湿法纺纱制备小球藻蛋白/海藻酸钠互穿结构凝胶纤维。将小球藻粉溶于100 mL去离子水中,用考马斯亮蓝法[15]测定其中蛋白质含量,标准曲线为Y=5.66X+0.078(R2=0.999 9),调节其中小球藻蛋白质量浓度至1 g/100 mL。向溶液中加入1 g海藻酸钠,磁力搅拌30 min使其完全溶解。添加TG酶至其浓度分别为0,1.0,2.5,5.0,7.5和10.0 U/mL,置于50 ℃水浴下反应2 h。反应结束后,置于90 ℃水浴5 min灭酶。冷却至室温后,以5 000 r/min离心15 min除去气泡,所得样品即为纺丝液。
将纺丝液装入配备18G针头的注射器中,挤压至100 mL质量浓度为10 g/100 mL CaCl2溶液中。2 h后,将凝胶纤维取出,放入去离子水中浸泡1 min后取出,35 ℃下干燥24 h,得到小球藻蛋白/海藻酸钠互穿结构凝胶纤维。凝胶纤维按照其纺丝液中TG酶浓度分别标记为semi-IPN、IPN1、IPN2.5、IPN5、IPN7.5及IPN10。
1.3.2 傅里叶红外光谱(FT-IR)
将2 mg凝胶纤维与200 mg溴化钾充分混合后压片。采用FT-IR5700傅里叶红外光谱仪对凝胶纤维的结构进行表征,波数范围4~4 000 cm-1,分辨率4 cm-1。
1.3.3 二级结构分析
以所得红外光谱图为基础,参考谢孟峡等[16]的方法对凝胶纤维中小球藻蛋白的4种二级结构相对含量进行测定。
1.3.4 小球藻蛋白交联度的测定
小球藻蛋白交联度的测定按照Mi等[17]提出的方法。溶液Ⅰ配制:1.05 g柠檬酸,10 mL浓度为1.0 mol/L的NaOH溶液,0.04 g SnCl2·2H2O混合后,用去离子水定容至25 mL。溶液Ⅱ配制:1 g茚三酮溶解于乙二醇单甲醚中,定容至25 mL。将溶液Ⅰ与溶液Ⅱ混合,磁力搅拌45 min,避光储存。
将1 mL纺丝液溶解于去离子水中,定容至100 mL。将1 mL稀释后纺丝液与2 mL茚三酮溶液混合,100 ℃水浴20 min。冷却至室温后,加入3 mL 50%异丙醇,在570 nm波长下测量吸光度。与茚三酮反应后,样品中游离氨基含量与其在570 nm波长下吸光度呈正比。样品中游离氨基含量可代入甘氨酸浓度与其在570 nm下吸光度绘制所得标准曲线进行计算Y=2.805X+0.030(R2=0.999 8)。小球藻蛋白交联度按式(1)计算。
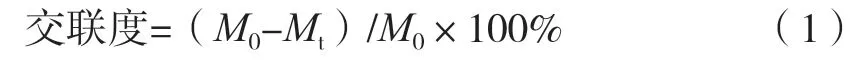
式中:M0为不添加TG酶的样品中游离氨基物质的量浓度,mmol/L;Mt为不同TG酶添加量的样品中剩余游离氨基酸物质的量浓度,mmol/L。
1.3.5 吸水能力测定
将一定质量的凝胶纤维置于去离子水中,37 ℃浸泡24 h。取出后用滤纸擦干表面水分,记录质量。凝胶纤维的吸水性按式(2)计算。
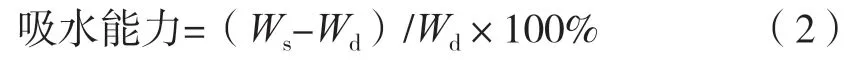
式中:Wd为凝胶纤维干质量,g;Ws为凝胶纤维吸水后质量,g。
1.3.6 持水能力测试
将一定质量的凝胶纤维置于去离子水中,37 ℃浸泡24 h。以5 000 r/min离心10 min,取出纤维,用滤纸擦干表面水分,记录质量。凝胶纤维的持水性按式(3)计算。
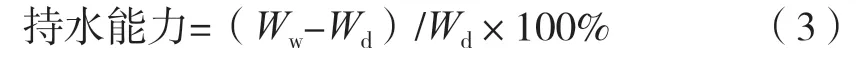
式中:Wd为凝胶纤维干质量,g;Ww为凝胶纤维离心后质量,g。
1.3.7 凝胶纤维的酶促降解
凝胶纤维的酶促降解参照Bidault等[18]的方法并稍作修改。将50 mg凝胶纤维浸泡于100 mL生理盐水中,调节pH至8.5,加入碱性蛋白酶至400 U/mL。55 ℃水浴下酶解,每隔1 h,吸取2 mL酶解液,于90 ℃水浴5 min灭酶。每次收集样品后,将2 mL酶溶液加入反应体系来保证酶浓度。试验结束后,用考马斯亮蓝法[15]测定酶解液中释放的蛋白质以及蛋白质片段含量。一定时间内小球藻蛋白及其酶解物的释放率为溶液中释放蛋白质片段质量与蛋白质初始质量的比值。
1.3.8 数据统计与分析
采用SPSS 19.0分析软件和Origin 8.0软件处理所得试验结果,数据均为3个平行样本,以平均值±标准差表示,采用ANOVA对数据的差异显著性进行分析(p<0.05表示差异显著)。
2 结果与分析
2.1 凝胶纤维红外测试分析
运用傅里叶红外光谱从定性的角度分析凝胶纤维制备的可行性,小球藻粉、海藻酸钠、凝胶纤维的红外光谱如图1所示。在海藻酸钠的红外光谱中,3 420 cm-1的吸收峰是—OH的伸缩振动;2 929 cm-1的吸收峰是饱和C—H的伸缩振动;1 635与1 417 cm-1的吸收峰则分别对应—COO—的非对称伸缩振动和对称伸缩振动[10]。
在小球藻粉的红外光谱中,酰胺Ⅰ带(位于1 700~1 600 cm-1代表C==O伸缩振动)与酰胺Ⅱ带(位于1 590~1 500 cm-1代表N—H弯曲振动与C—N伸缩振动)分别位于1 651与1 554 cm-1处[19]。3 419 cm-1的吸收峰是—OH伸缩振动与N—H伸缩振动重叠的结果[10]。
TG酶添加量不同时凝胶纤维的红外光谱均表现出海藻酸钠与小球藻粉的特征吸收峰,说明两者具有良好的相容性[20]。与semi-IPN相比,具有IPN结构的凝胶纤维在1 554 cm-1的吸收峰强度略有增强,在1 631 cm-1的吸收峰强度明显增强,说明TG酶成功催化谷氨酰胺与赖氨酸进行共价交联,生成酰胺键[21],形成小球藻蛋白网络结构。具备IPN结构的凝胶纤维均在1 157 cm-1处呈现吸收峰(代表C—O伸缩振动),而semi-IPN并无此吸收峰,这可能是由于TG酶的催化作用引发小球藻蛋白构象改变[19]。
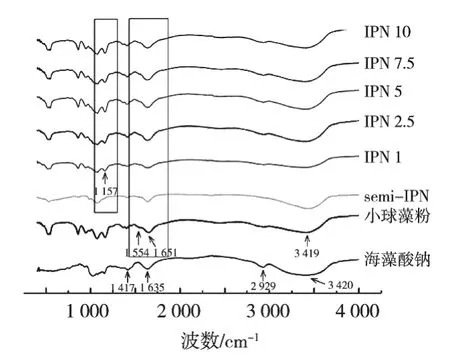
图1 小球藻粉、海藻酸钠、凝胶纤维的FT-IR曲线
2.2 小球藻蛋白二级结构分析
通过对小球藻蛋白二级结构进行表征,进一步了解不同TG酶添加量对凝胶纤维结构的影响,结果如表1所示。数据表明,TG酶催化效应对小球藻蛋白二级结构产生显著影响。与semi-IPN相比,随着TG酶添加量增加,β-折叠含量从22.83%逐渐下降至17.00%,无规则卷曲含量从15.58%逐渐上升至24.41%。加入TG酶之后,α-螺旋含量总体呈现下降趋势,TG酶浓度7.5 U/mL时达最低值28.51%。TG酶添加量的改变并未对β-转角含量造成显著影响(p>0.05),只是当TG酶浓度1 U/mL时达最高值34.41%,显著高于其他样品(p<0.05)。上述现象与Song等[22]研究结果相似。研究显示,二级结构与蛋白质柔韧性以及强度密切相关。Zhao等[23]研究发现α-螺旋含量下降说明分子内氢键含量降低。Liang等[24]研究发现β-折叠的稳定性高于α-螺旋,蛋白质结构的强度很大程度上取决于β-折叠含量。Chen等[25]研究发现无规则卷曲含量增加则代表蛋白质中的高度有序结构展开。结果表明,随着TG酶添加量增加,凝胶纤维中小球藻蛋白的结构从有序逐渐向无序转变。
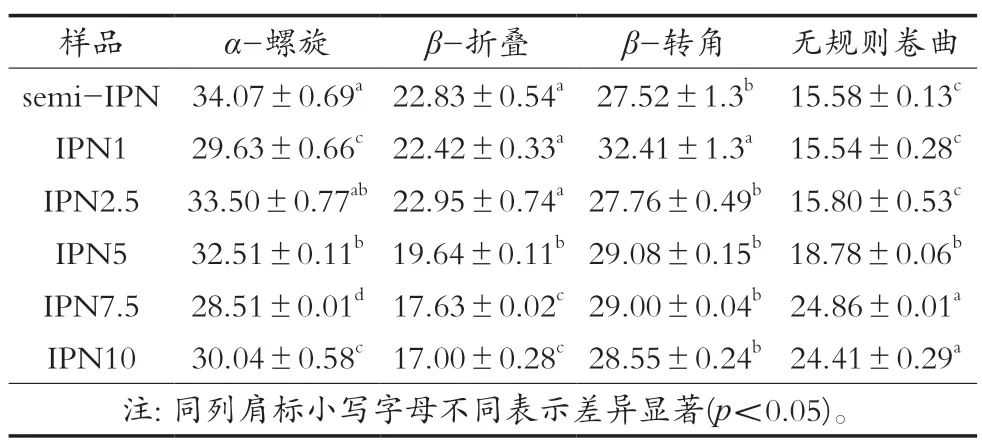
表1 不同TG酶添加量下小球藻蛋白二级结构相对含量 单位:%
2.3 小球藻蛋白交联度分析
通过测定小球藻蛋白的交联度,从定量角度分析IPN凝胶纤维制备的可行性。TG酶催化蛋白质之间生成ε-(γ-Glu)Lys共价键,从而减少赖氨酸中ε-氨基含量[26]。茚三酮可以测得样品中游离氨基含量,进而确定交联度。如图2所示,随着TG酶添加量从1 U/mL上升至10 U/mL,小球藻蛋白交联度从9.67%上升至16.26%,表明TG酶成功催化蛋白质交联并形成网络结构,与FT-IR中结果一致。与此同时,海藻酸钠导致纺丝液黏度增高,阻碍TG酶的催化效应,导致交联度有限[10]。
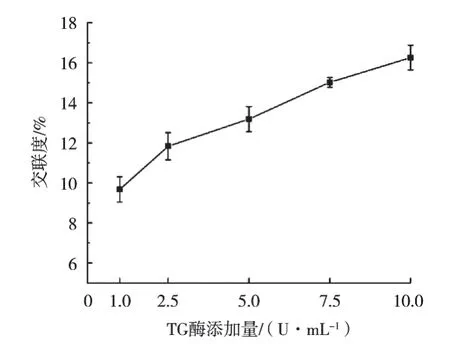
图2 不同TG酶添加量下凝胶纤维中小球藻蛋白交联度
2.4 吸水能力分析
吸水能力是衡量材料可否应用于创面敷料的重要参数,不同TG酶添加量的凝胶纤维的吸水能力为82.64%~62.76%,如图3所示。Shi等[27]认为凝胶纤维的吸水性与其中亲水基团的数量及基团亲水性强弱密切相关。在凝胶纤维中,水分子与羟基、羧基等亲水基团密切结合,表现出较好的吸水能力。随着TG酶添加量从0上升至10 U/mL,凝胶纤维的吸水性下降24.06%。尽管TG酶的催化作用使小球藻蛋白结构从有序向无序转变,但是高TG酶添加量使小球藻蛋白的交联度增加、网络结构中网格孔径降低[28]。致密的网络结构阻止水渗透进入凝胶纤维之中,从而降低吸水能力。
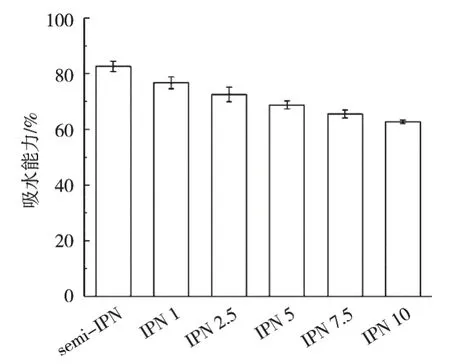
图3 不同TG酶添加量对凝胶纤维吸水性的影响
2.5 持水能力分析
持水能力是影响吸水材料性能的关键因素之一,不同TG酶添加量的凝胶纤维的吸水能力为45.98%~59.48%,如图4所示。水分析渗透进入凝胶纤维之中,与海藻酸钠、小球藻蛋白形成稳定的氢键,或与亲水基团紧密连接,表现出较好的持水能力。随着TG酶添加量从0上升至10 U/mL,凝胶纤维的吸水能力上升29.36%,这是由凝胶纤维的结构与强度共同导致[29]。TG酶添加量增加催化小球藻蛋白进一步交联形成更加致密、均匀的网络结构,可有效防止水分流失。与此同时,TG酶催化形成的ε-(γ-Glu)Lys共价键强度是氢键和疏水作用力的20余倍[30],因此保证凝胶纤维的持水性。
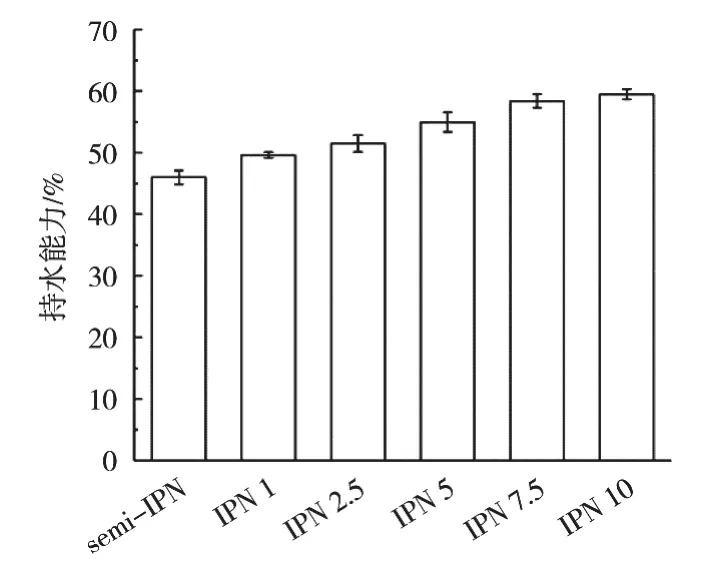
图4 不同TG酶添加量对凝胶纤维持水能力的影响
2.6 凝胶纤维的酶促降解过程
经过15 h的降解过程,凝胶纤维部分降解,最终形成细小、均匀的碎片,说明海藻酸钠与小球藻蛋白在凝胶纤维中均匀分布[18]。溶液中的Na+与凝胶纤维中的Ca2+发生置换反应,海藻酸钠网络裂解[31],由于不能被碱性蛋白酶催化降解,便以碎片的形式存在。所有样品中的小球藻蛋白在降解14 h后均完全释放至酶溶液中,说明尽管小球藻蛋白部分交联,但仍对碱性蛋白酶敏感,使其可以渗透进海藻酸钠网络结构,催化小球藻蛋白网络结构的裂解[18]。不同TG酶添加量的凝胶纤维在降解过程中的蛋白质组分(小球藻蛋白及其酶解物)释放率如图5所示。尽管不影响最终释放量,但TG酶添加量增加明显减缓蛋白质组分的释放速率(除semi-IPN凝胶纤维外)。TG酶添加量从1 U/mL上升至10 U/mL,蛋白质组分完全释放的时间从11 h增加至14 h,50%蛋白质组分释放的时间从4 h增加至11 h。这是由于随着TG酶添加量增加,凝胶纤维吸水能力降低,阻碍小球藻蛋白网络与碱性蛋白酶活性位点的结合[32]。Semi-IPN中蛋白质组分的释放速率并不是最快,可能是由于蛋白酶仅作用于小球藻蛋白,与IPN结构凝胶纤维中双网络结构同时裂解相比,其网络结构降解速度较慢。

图5 不同TG酶添加量的凝胶纤维中小球藻蛋白释放曲线
3 结论
制备小球藻蛋白/海藻酸钠凝胶纤维,通过FT-IR及小球藻蛋白交联度测试证明凝胶纤维具备互穿网络结构。二级结构分析显示,TG酶催化使β-折叠含量减少,无规则卷曲含量增加,小球藻蛋白结构从有序向无序转变。TG酶添加量的增加赋予凝胶纤维更加致密的网络结构,其吸水能力减弱,持水能力加强。所有的凝胶纤维在蛋白酶以及生理盐水作用下在14 h之内完全降解。TG酶含量从1 U/mL上升至10 U/mL,小球藻蛋白及其酶解物的释放速率逐渐下降。试验结果有助于扩展小球藻蛋白应用领域,为其深层次加工利用提供理论基础。
猜你喜欢
陶瓷学报(2019年5期)2019-01-12
电源技术(2016年2期)2016-02-27
北京信息科技大学学报(自然科学版)(2016年6期)2016-02-27
管理现代化(2016年3期)2016-02-06
管理现代化(2016年3期)2016-02-06
智能系统学报(2015年4期)2015-12-27
石油化工应用(2014年12期)2014-03-11
石油化工应用(2014年11期)2014-03-11
华东理工大学学报(自然科学版)(2014年1期)2014-02-27
断块油气田(2012年6期)2012-03-25
