
红松不同生活史阶段的枝叶权衡
2021-05-21 08:07尹凤娟王明琦金光泽刘志理
林业科学 2021年4期
尹凤娟 王明琦 金光泽 刘志理
(东北林业大学生态研究中心 森林生态系统可持续经营教育部重点实验室 哈尔滨 150040)
植物的枝叶大小关系反映了构型塑造和空间资源利用能力,是生态策略的主要维度之一(Westobyetal., 2006)。枝结构影响叶片大小和展叶强度的变化,从而决定叶片大小和数量的权衡以及枝叶间生物量分配(Yangetal., 2008),叶片的大小及数量分布等影响植物对碳的获取,并进一步影响枝的生长和生存(Xiangetal., 2009)。叶片和枝的性状在生物力学和形态结构上高度协调,决定了植物对光照、水分等资源的利用和适应策略(Kawaietal., 2019)。研究发现,植物在资源紧张的环境中会随着生活史梯度的上升由资源获取型转向资源保守型(Daminetal., 2018; Dayrelletal., 2018)。此外,不同枝叶年龄的枝叶资源分配策略和净光合作用存在差异(Tangetal., 2019),枝叶性状的调整对树木生长和森林净初级生产力产生影响(Maetal., 2020)。因此,揭示枝叶大小关系随生活史梯度和枝叶年龄的变异规律能客观地反映植物对环境的适应机制,对深入了解植物资源权衡策略具有重要意义。
目前对于植物枝叶关系主要集中于“枝-叶大小关系”(Westobyetal., 2003; Sunetal., 2006)和“叶大小-数量关系”(Kleimanetal., 2007; Yanetal., 2013)。White(1983)提出的“Corner’s rules”假说认为横截面积较大的枝,其单叶面积较大,但分枝较小。对“Corner’s rules”的进一步研究证明枝-叶大小关系存在着显著的正相关关系(Sunetal., 2006; Yangetal., 2009),然而枝-叶大小关系的异速和等速生长依然存在争议。比如不同功能类群叶和枝的异速生长与不同的环境条件有关(Mengetal., 2015); 当小枝的功能局限于维管供应和生物力学支持时,枝横截面积与枝上的叶片面积呈等速生长关系,而其他压力作用于树枝时,枝-叶的大小关系将成为异速生长(Brouatetal., 2001)。植物为适应环境往往通过权衡来分配有限资源,因此Kleiman等(2007)提出了“出叶强度优势”假说,认为高出叶强度与大的单叶面积不可兼得。后续研究表明单叶面积和出叶强度的权衡在植物中普遍存在,比如不同海拔、不同坡向的单叶面积与出叶强度呈显著负相关关系(杨冬梅等, 2012a; 李曼等, 2017)。然而,刘长柱等(2015)对48种常见树种的研究表明,不同树种、生活型和森林类型的叶大小和出叶强度之间的权衡关系并不一致。
在木本植物个体发育的不同阶段(幼苗、幼树、成年树和老龄树),其成熟叶片在生理和形态特征上往往不同。矮小的幼苗光照受到严重限制,资源获取困难、养分利用有限,对水热因子的变化敏感(Wawrzyniaketal., 2020)。拥有成熟根系的老龄树,更易获得土壤水分和氧分,分配给组织水平上更多的生物量(Hendriketal., 2012),但同时随着茎干粗度和树高增大,水分运输距离增加,导致水分上行受阻,树冠顶层叶片可能面临着缺水问题(Gleasonetal., 2016)。不同生活史阶段的植物性状的可塑性存在差异,某一生活史阶段植物枝叶性状的权衡策略并不一定适用于其他生活史阶段(Lumetal., 2019)。探究不同生活史阶段的植物功能性状的变化,有助于更深刻地认识植物枝叶的发育规律及对环境的适应机制。
常绿树种不同枝叶年龄的枝叶形态、生理特征均具有明显差异(Petersetal., 2008),当年生小枝容易随外界环境发生改变(杨冬梅等, 2012b),同时当年生小枝对于营养物质和碳水化合物的运输方面比多年生枝更灵活(Pickupetal., 2005)。然而,多年生枝叶所占的比例远大于当年生枝叶,多年生叶片合成更多的光合产物(张小全等, 2000; Robakowskietal., 2017),因此多年生枝叶对树冠结构和光截获能力具有重要意义。但当年生枝-叶大小关系是否能推及到多年生枝-叶大小关系尚无定论。因此,解析枝叶性状的变异及权衡在生活史阶段及不同枝叶年龄的变化规律对于掌握植物对生境变化的响应策略具有重要意义。
本研究以阔叶红松(Pinuskoraiensis)林的建群种红松为研究对象,针对其不同生活史阶段(幼苗、幼树、成年树、老龄树)的红松样树,测定其不同枝叶年龄的4个性状,包括枝横截面积、单叶面积、总叶面积和出叶强度。旨为解析枝横截面积、单叶面积、总叶面积及出叶强度在不同生活史阶段和不同枝叶年龄的变异情况; 检验生活史和枝叶年龄对性状间相关关系是否存在显著调控作用。
1 材料和方法
1.1 研究区域概况
研究调查地位于黑龙江凉水国家级自然保护区(128°47′8″—128°57′19″E, 47°6′49″—47°16′10″N),该保护区气候类型为温带大陆性季风气候,冬季漫长且寒冷干燥,夏季雨热同期,年平均气温-0.3 ℃,年平均降水量676 mm。该区域的地带性顶极植被为阔叶红松林,地带性土壤为暗棕壤。森林覆盖率96%,有原始成过熟林4 100 hm2,红松林面积占其中的80%,是目前我国保存较为完整的原始阔叶红松混交林分布区之一。
1.2 数据采集
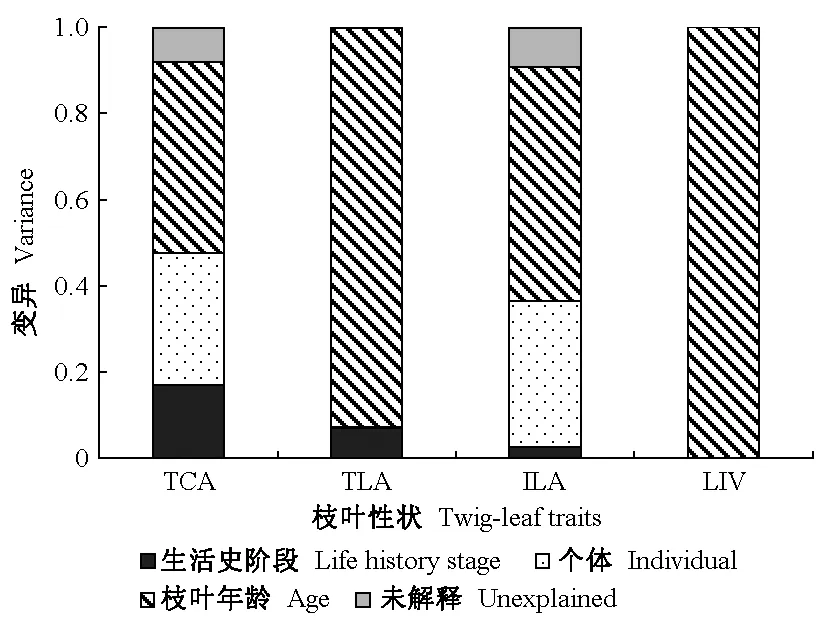
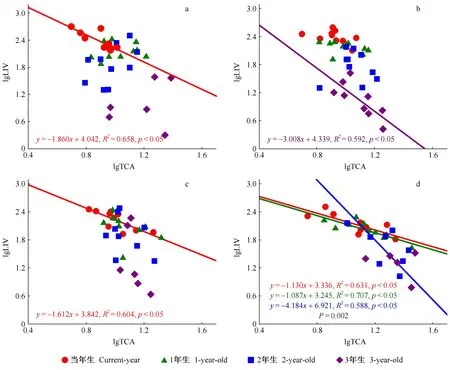
2019年10月在黑龙江凉水国家级自然保护区内,依托9 hm2阔叶红松林动态监测样地(徐丽娜等, 2012),根据红松不同个体发育阶段的特征选取4个生活史阶段的红松样树为研究对象(祖占和, 1987),包括幼苗10株(DBH≤3 cm)、幼树10株(8 cm 式中:n为针叶数量;V为针叶体积(cm3);l为针叶平均长度(cm)。 根据每枝龄样枝所支撑针叶的总叶面积和针叶数量的比值计算单叶面积; 样枝的枝横截面近似为圆形,基于基径测定其枝横截面积; 出叶强度为总针叶数与样枝体积的比值(Yanetal., 2013)。 表1 样树胸径信息①Tab.1 Information of diameter at breast height (DBH) for sample trees 用最小显著差异法(LSD)检验不同生活史阶段和不同枝叶年龄枝、叶性状的差异性,LSD检验类似于Fisher’s检验法可以检验2组数据的差异性(Hessetal., 2017),显著性水平设置为α = 0.05。为使数据符合正态分布,本研究将性状值进行(以10为底)对数转换后再进行数据分析。采用嵌套方差分析法解析枝叶性状变异,即采用‘nlme’包中的‘lme’函数针对每个性状构建线性混合模型,其嵌套水平为: 生活史阶段>个体>枝叶年龄(Paradisetal., 2004)。 枝叶功能性状间的相关关系采用函数为y=bxa,并转换成lg(y) = lg(b) +alg(x),式中:x和y为2个性状,b为性状间相关关系的截距,a为性状间相关关系的斜率,即相对生长指数或异速生长参数。当|a| = 1时,表示二者是等速变化关系; 当|a|显著偏离1时,二者间为异速变化关系(Harveyetal., 1991)。采用标准化主轴估计法(standardized major axis, SMA)检验生活史或枝叶年龄对性状间相关关系是否存在显著影响,由软件smatr Version 2.0计算,其中性状间相关关系的斜率与1或-1的显著性检验基于Warton和Weber(2002)研究方法判定。采用R-3.6.1(R Core Team, 2018)对数据进行统计分析,用SigmaPlot 10.0 和 Excel软件绘图。 老龄树阶段枝横截面积和总叶面积显著高于其他3个阶段; 单叶面积在幼苗阶段显著低于其他3个阶段; 出叶强度随植株生长逐渐降低(图1)。不同生活史阶段总叶面积、出叶强度随叶龄增加呈下降趋势; 枝横截面积随枝龄增加呈上升趋势,但成年树阶段各枝龄枝横截面积无显著差异; 幼苗和幼树阶段单叶面积随叶龄增加呈下降趋势,成年树和老龄树阶段不同叶龄的单叶面积无显著差异(图1)。 图1 红松不同生活史阶段枝叶性状的变异Fig. 1 The variations of twig and leaf traits of Pinus koraiensis at different life history stages Ⅰ、Ⅱ、Ⅲ、Ⅳ分别代表幼苗阶段、幼树阶段、成年树阶段、老龄树阶段。大写字母和小写字母分别代表不同生活史阶段间性状存在显著差异(P < 0.05)和不同枝叶年龄间性状存在显著差异(P < 0.05)。Ⅰ, Ⅱ, Ⅲ, Ⅳ represent the seedling, sapling, adult tree, and old tree stage, respectively. The capital letters and lowercase letters indicate that twig or leaf traits are significant differences between different life history stages and between different leaf and twig ages at 0.05 level, respectively. 枝叶年龄对枝叶性状变异的解释较多,解释了出叶强度变异的99%、总叶面积变异的93%、单叶面积变异的54%和枝横截面积变异的44%(图2)。枝叶性状在不同生活史阶段间的变异较小,生活史阶段分别解释了枝横截面积、总叶面积和单叶面积变异的17%、7%和3%(图2)。 图2 红松枝叶性状的方差分解Fig. 2 Variance decomposition of twig and leaf traits of Pinus koraiensis TCA: 枝横截面积; TLA: 总叶面积; ILA: 单叶面积; LIV: 出叶强度。下同。解释程度小于0.1%在图中显示为0。TCA: Twig cross-sectional area; TLA: Total leaf area; ILA: Individual leaf area; LIV: Volume-based leafing intensity. The same below. The degree of interpretation less than 0.1% is shown zero in theFigure. 枝横截面积与总叶面积在幼苗和老龄树阶段均呈显著正相关(图3a); 枝横截面积与单叶面积仅在幼苗阶段呈显著正相关(图3b); 枝横截面积与出叶强度在幼苗、成年树及老龄树阶段均呈显著负相关(图3c); 出叶强度与单叶面积仅在幼苗阶段呈显著负相关(图3d)。然而,这些相关关系在不同生活史阶段无显著差异(图3)。 在幼苗和老龄树阶段枝横截面积与总叶面积存在共同斜率1.693,且斜率与1存在显著差异,即呈异速生长关系(图3a); 枝横截面积与单叶面积在幼苗阶段相关关系的斜率为0.682 6,呈异速生长关系(图3b); 幼苗、成年树及老龄树阶段的枝横截面积与出叶强度存在共同斜率-1.478,与-1存在显著差异,呈异速生长关系(图3c); 出叶强度与单叶面积仅在幼苗阶段相关关系斜率与-1存在显著差异(图3d)。 图3 红松不同生活史阶段当年生枝叶性状关系的差异性Fig. 3 Diversities in relationships between current-year twig and leaf traits of Pinus koraiensis at different life history stages 图3c中枝横截面积(TCA)与出叶强度(LIV)在幼苗、成年树和老龄树阶段均存在相关关系。InFig.3c, there is a correlation between the twig cross-sectional area (TCA) and volume-based leafing intensity (LIV) at seedling, adult tree and old tree stages. 幼苗阶段当年生枝横截面积与总叶面积显著正相关(图4a); 幼树和成年树阶段,仅1年生枝横截面积与总叶面积显著正相关,且相关关系斜率与1均存在显著差异(图4b、c); 老龄树阶段,当年生及1年生枝横截面积与总叶面积均显著正相关,且当年生与1年生存在共同斜率1.377(图4d),呈异速生长关系(图4)。 幼苗和成年树阶段,当年生枝横截面积与出叶强度均呈显著负相关(图5a、c); 幼树阶段,仅3年生枝横截面积与出叶强度显著负相关(图5b); 老龄树阶段,当年生、1年生和2年生枝横截面积与出叶强度均呈显著负相关关系(图5d)。此外,枝横截面积与出叶强度的相关关系斜率在幼苗、幼树和成年树阶段与-1均存在显著差异,呈异速生长关系(图5a、b、c); 而在老龄树阶段,当年生和1年生枝横截面积与出叶强度的相关关系斜率与-1不存在显著差异,2年生枝横截面积与出叶强度相关关系斜率为-4.184,与-1存在显著差异(图5d)。 图4 不同枝叶年龄枝横截面积(TCA)与总叶面积(TLA)相关关系Fig. 4 Relationship between cross-sectional area of twigs(TCA) and total leaf area(TLA) across leaf and twig ages a、b、c、d分别代表幼苗阶段、幼树阶段、成年树阶段和老龄树阶段。a, b, c and d respectively represent seedling, sapling, adult tree and old tree stages. 图5 不同枝叶年龄枝横截面积(TCA)与出叶强度(LIV)相关关系Fig. 5 Relationship between cross-sectional area of twigs(TCA) and volume-based leafing intensity(LIV) across leaf and twig ages a、b、c、d分别代表幼苗阶段、幼树阶段、成年树阶段和老龄树阶段。a, b, c and d respectively represent seedling, sapling, adult tree and old tree stages. 随生活史梯度上升,红松的叶面积、枝横截面积呈上升趋势,出叶强度呈下降趋势(图1)。在树木生长过程中,叶片容易受到环境的影响并作出响应,而叶片大小是植物适应环境中热量和水分的结果(王常顺等, 2015)。光照和生物量分配使得红松幼苗初级生产力较低,对养分和资源的获取利用有限(Yangetal., 2010)。随着树高增加,树木需要降低顶部叶片的水势,以克服重力造成的水势差(Liuetal., 2020),叶片资源利用策略从增强光合作用转向防止机械损伤(Atsushietal., 2005),因此,随着生活史梯度的上升,植株更倾向于将资源分配于枝,以增强其抗损伤能力,有利于适应环境变化。此外,较小的单叶面积不但能够有效减少蒸腾损失,而且其较小的边界阻力也有利于热交换(Jacquelineetal., 2006)。因此植株在幼苗时期选择较小的单叶面积和高出叶强度,而在老龄树阶段选择较大的单叶面积和低出叶强度。随着植株生长,叶对光的截获能力和碳的获取能力上升(Niklasetal., 2007),总叶面积在老龄树时期相对较高。增大的叶面积和叶片蒸腾速率,也要求小枝具有更高的支持能力和供水效率(Niinemetsetal., 2005)以及较大的枝横截面积来增加机械载荷的强度(Fanetal., 2017),减少叶片之间对光的相互遮挡,因此枝横截面积在老龄树阶段显著高于其他阶段。 不同生活史阶段,随着枝叶年龄的上升,叶面积和出叶强度呈下降趋势,枝横截面积呈上升趋势(图1)。可能是因为,当年生叶片生长于植株外侧,光照条件较好,植物将有限资源更多地投入到当年生叶片中,可以最大化光合收益; 此外,当年生小枝选择较高的出叶强度,可以提升其光合同化能力(Cobleetal., 2015)。而多年生针叶位于树冠内侧,由于叶片的遮蔽作用,光照条件受限,同时光合速率逐渐减弱(霍宏等, 2007; Zhaetal., 2010),其对于结构完整性或防御干扰方面投入更多(Wangetal., 2017),因此植物对多年生的叶面积和叶数量的投入也会相应减少。 枝叶年龄水平对枝叶性状变异的解释较多,可能是因为本研究对同一枝条的不同枝龄进行取样,更好地表征植物枝叶年龄水平的性状变异。生活史阶段主要解释了枝横截面积的17%,可能是因为红松枝比叶的性状更能预测不同生活史阶段红松的变异(Poorteretal., 2008)(图2)。为了研究红松枝叶性状在不同生活史阶段、不同枝叶年龄的权衡策略,本研究针对生活史阶段和枝叶年龄对红松枝叶性状关系的影响机制进行了进一步探究。 枝叶性状在幼苗阶段均存在显著的相关关系,而幼树阶段均不显著,成年树阶段仅枝横截面积与出叶强度显著负相关,老龄树阶段枝横截面积与总叶面积呈显著正相关、与出叶强度呈显著负相关(图3)。不同生活史阶段枝横截面积与叶面积的正相关关系与“Corner’s rules”假说相符,即大枝对大叶,小枝对小叶。单叶面积与出叶强度的负相关关系,也与之前的研究(杨冬梅等, 2012a; Yanetal., 2013)相符,植物利用体内有限的生物量选择少量的大面积叶或者大量的小面积叶(Kleimanetal., 2007)。根据“Corner’s rules”假说和“出叶强度优势假说”,枝横截面积与出叶强度呈负相关关系(Milla, 2009)。仅在幼苗阶段的枝叶都存在显著的正相关关系或负相关关系,而在其他生活史阶段间不显著的原因可能是不同生活史阶段的红松碳储量不同(Maetal., 2020),尽管随着红松树龄增长,植株固定的总生物量上升,但更大比例的生物量被分配给支持组织,因此,性状之间相关性降低(Hendriketal., 2012)。由此可见,幼苗的枝-叶性状关系比成年树或老龄树更强。 幼苗阶段与老龄树阶段枝横截面积与总叶面积呈现出异速生长指数均大于1的生长规律,说明枝横截面积的增长速度均小于总叶面积(图3)。这种异速生长关系反映了植物对资源利用和同化产物的分配方式。由于叶片是植株获取能量的主要器官,在植株发育过程中为了提高光合效率,减缓了小枝的生长速度(Kawaietal., 2019)。幼苗阶段枝横截面积与单叶面积异速生长指数小于1,枝横截面积的增长速度大于单叶面积,说明细枝比叶片需要更多的碳和能量(Tangetal., 2019),同时植物选择增大对枝的机械投入来抵御气候和环境压力(Gleasonetal., 2016)。本研究中红松出叶强度与单叶面积呈负相关关系(-1<异速生长指数<0),这与杨冬梅等(2012a)的研究结果一致,说明红松叶也存在这种大小和数量上的权衡。另外,红松枝叶大小关系在不同生活史阶段不存在显著差异,可能是其枝叶大小间始终维持着一个固有的平衡特征,生活史阶段的变化无法改变红松枝叶在大小和数量上的权衡策略。 幼苗阶段,当年生枝横截面积与总叶面积显著正相关,幼树和成年树阶段1年生枝横截面积与总叶面积均呈显著正相关(图4)。老龄树阶段当年生、1年生和2年生枝横截面积与出叶强度呈负相关关系(图5)。但并不是各个枝叶年龄都存在显著的“枝-叶大小关系”和“叶大小-数量关系”。不同叶龄的针叶在光合、呼吸、蒸腾速率等方面存在差异(霍宏等, 2007; Petersetal., 2008),随着叶龄的上升,针叶的需光量增加,同时多年生叶片受到更多的竞争压力和环境胁迫。当年生叶常处于树冠顶层或外围,光环境优越,营养物质更趋向于投入给当年生的枝叶(Tangetal., 2019)。而2年生及2年生以上针叶一般处于树冠的里层和下层,受光条件较差,与周围环境的热交换率较低,使其在干燥环境下可能受到热损害,或在寒冷环境中有霜冻损害的风险(Dongetal., 2020)。目前大多数研究都集中在当年生枝叶,所以这种枝-叶相关关系是否可以从当年生推及到多年生还有待验证。 植物生长发育过程中,随着资源可获得性的改变,其对资源的利用策略也会随之变化。红松“枝-叶大小关系”和“叶大小-数量关系”受生活史阶段和枝叶年龄的影响,随生活史梯度上升,红松枝横截面积、叶面积呈上升趋势,出叶强度呈下降趋势。通过分析红松枝、叶性状及其间异速生长的差异,可以更好地理解种内变异的影响机制,从而为预测植物对不同生境的响应策略奠定基础。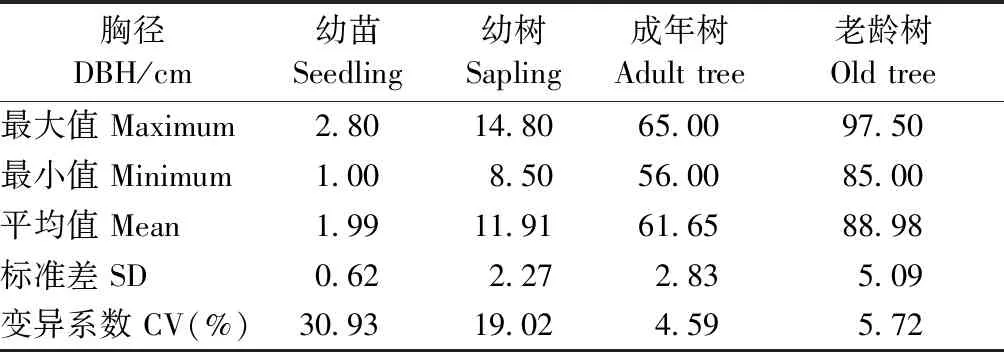
1.3 统计分析
2 结果与分析
2.1 红松不同生活史阶段枝叶性状变异

2.2 红松不同生活史阶段当年生枝叶性状的异速生长关系

2.3 枝叶年龄对枝叶性状相关关系的影响

3 讨论
3.1 枝叶性状在不同生活史阶段的变异规律
3.2 枝叶性状关系与生活史阶段的关联性
3.3 枝叶性状关系与枝叶年龄的关联性
4 结论
猜你喜欢
农学学报(2022年8期)2022-11-16
格言·校园版(2022年17期)2022-07-06
小哥白尼(野生动物)(2021年9期)2022-01-17
伦理学研究(2021年6期)2021-12-29
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
新农民(2020年5期)2020-12-10
伦理学研究(2020年4期)2020-09-07
新疆农业科学(2020年1期)2020-02-14
农民致富之友(2016年24期)2017-04-26
河北渔业(2014年11期)2014-11-17
