
黄淮麦区部分小麦品种粒重基因 TaGS5-A1的等位变异分布及其效应分析
2021-05-25 07:03李静雯董亚超肖永贵李法计杨舒蓉夏先春何中虎
麦类作物学报 2021年3期
李静雯,董亚超,肖永贵,李法计,杨舒蓉,李 鸣,夏先春,何中虎,3
(1.甘肃省农业科学院生物技术研究所,甘肃兰州 730070; 2.中国农业科学院作物科学研究所国家小麦改良中心,北京 100081; 3.CIMMYT中国办事处,北京 100081)
小麦是我国第三大粮食作物,其产量的稳步提升对于保障国内粮食安全具有重大意义[1]。单位面积穗数、穗粒数和粒重构成小麦产量三要素[2]。研究表明,千粒重的提高对小麦产量潜力提升贡献较大[3-5]。籽粒大小和籽粒灌浆程度直接决定小麦粒重[6],粒重与籽粒大小、形状(粒长、粒宽、籽粒长宽比和粒厚)正相关[7],且受多个微效基因控制。因此,深入了解粒重基因及其分子标记对于提升小麦产量潜力具有重要意义[8]。
近年来小麦粒重相关基因研究取得了一定进展,已克隆出多个粒重相关基因,包括TaGW2-6A、TaGW2-6B[9-11]、TaGS-D1[12]、TaGS5-A1[13]、TaTGW6[14-15]、TaGW8-B1[16]、TaGL3-5A[17]、TaSDIR1-4A[18]、TaSus2-2B[19]、TaSus2-2A、TaSus1-7A[20]、TaCwi-A1[21]、TaCWI-D1[22]、TaCKX6-D1[23]、TaCYP78A3[24]、TaTPP-6AL1[25]、TaSAP1-A1[26]、Tabas1-B1[27]、Ta-6-SFT[28]、TaGS1[29]、TaBT1[30]、TaSST[31]、TaSnRK2.10[32-33]、TaSnRK2.9-5A[34]等。上述粒重相关基因的克隆、功能标记的开发及其与农艺性状关系的研究,为了解粒重的分子遗传机理及利用功能标记加快小麦高产和稳产育种进程提供了重要信息。
早期研究显示,水稻OsGS5编码的丝氨酸羧肽酶类蛋白(serine carboxypeptidase-like proteins,SCPLs)可影响籽粒灌浆,在调控水稻籽粒大小和粒重方面发挥着重要作用[35]。Xu等[36]研究发现,SCPLs已在多种植物中得到鉴定,表明其广泛参与植物体生长发育调控过程。Wang等[13]利用水稻OsGS5基因的生物学信息,克隆了普通小麦3A染色体上TaGS5-A1基因的全长编码序列,并针对其等位变异TaGS5-A1a和TaGS5-A1b开发了共显性酶切扩增多态性序列(cleaved amplified polymorphic sequences,CAPS) 功能标记,发现小麦TaGS5-A1基因对于籽粒大小具有正向调控作用,携带其优异等位基因TaGS5-A1b的小麦品种具有较宽的粒宽和更高的千粒重,在小麦高产育种中有较大的应用潜力。简大为等[37]和Li等[38]先后将TaGS5-A1基因CAPS功能标记用于检测新疆及黄淮麦区小麦,发现TaGS5-A1b基因型材料在新疆改良品种和黄淮麦区分布频率均高于50%,后者还验证了TaGS5-A1b基因与高粒重相关。
KASP技术作为一种高通量的单核苷酸多态性(single nucleotide polymorphism,SNP)或插入、缺失(insertion/deletion,InDel)基因分型平台,已广泛用于作物育种。近年来,小麦基因特异性KASP标记已被大量开发[39],并在实践中得到有效应用[40]。利用KASP 标记检测了109个长江上游小麦品种粒重等6个性状20个位点的基因型,结果发现多数材料为TaGS5-A1b基因型[41]。但是关于小麦主要粒重基因TaGS5-A1的 KASP标记在育种中的应用价值报道较少。
本研究以本课题组基于粒重基因TaGS5-A1开发的高通量KASP标记检测黄淮麦区119个小麦品种,了解其等位变异分布频率,并比较携带不同等位变异品种的粒重构成要素,探讨等位变异TaGS5-A1b和TaGS5-A1a的育种价值,以期为利用粒重分子标记进行基因聚合和培育高产稳产小麦品种提供参考。
1 材料与方法
1.1 供试材料
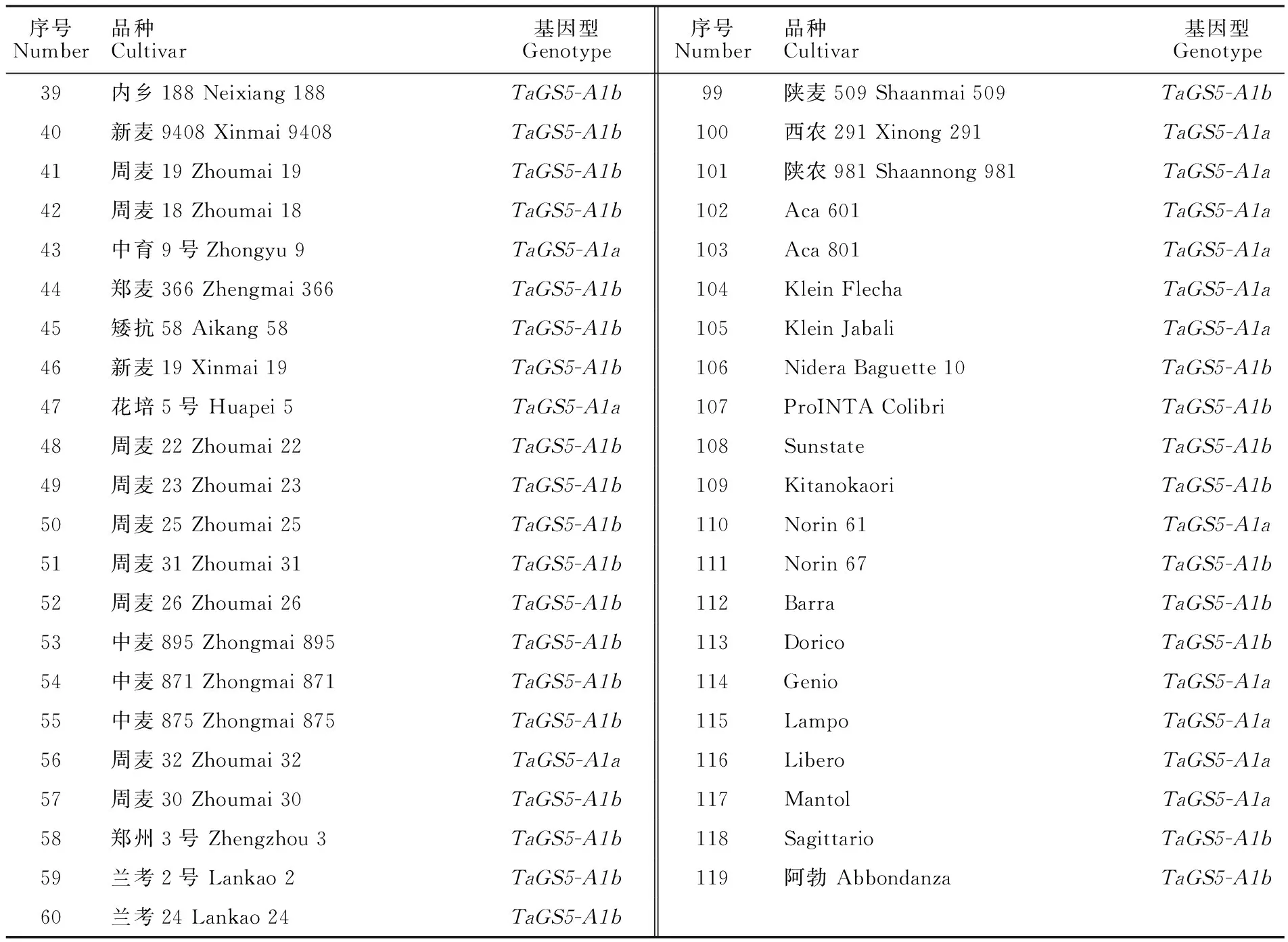
本研究采用119个小麦品种作为试验材料,其中,黄淮麦区(安徽、河北、河南、江苏、山东、山西和陕西7个省份)推广品种101个,适宜黄淮麦区生长的国外品种18个,该群体源于黄淮麦区具有代表性的166个小麦品种[42-44]。品种基本信息见表1。试验材料于2012-2013和2013-2014年度种植于中国农业科学院安阳棉花所白壁试验站(36°3′ N,114°29′ E,海拔65 m)和安徽省农业科学院濉溪试验站(33°53′ N,116°46′ E,海拔 30 m),2014-2015年度种植于中国农业科学院安阳棉花所白壁试验站和河北省石家庄市高邑县原种场(37°37′ N,114°36′ E,海拔45 m)。田间种植采用随机区组设计,3行区,行长2 m,3次重复。田间管理措施同当地大田。

表1 119个小麦品种的基本信息和基因型Table 1 Basic information and genotypes of 119 wheat varieties
(续表1 Continued table 1)
1.2 性状调查
调查性状包括千粒重、粒长、粒宽、穗长、株高和穗下节长。于小麦成熟期,每个小区随机选取20个穗子、10个单株分别测定穗长、株高和穗下节长,以小麦根茎结合处与顶部小穗之间的距离代表植株高度,以穗下部往下到第一个结节处的长度作为穗下节长。于小麦成熟后,单株收获脱粒,用万深SC-G型自动考种分析仪测定千粒重、粒长和粒宽。119个小麦品种的表型数据由Li等[44]调查提供。
1.3 小麦叶片DNA提取
每个品种挑选8粒种子,发苗7~10 d后,剪取一定量的叶片,采用高盐、低pH法提取DNA[45]。
1.4 基因型分型
采用粒重基因TaGS5-A1特异性KASP标记进行基因型分型,该基因标记由中国农业科学院作物科学研究所何中虎课题开发,具体引物序列见表2。根据KASP标记标准方法对119个小麦品种TaGS5-A1基因的目标SNP位点进行KASP分析。
PCR反应在SNPline水浴PCR热循环仪Hyfrocycler2(LGC,英国)384孔PCR板进行,每个PCR反应体系为5.0 μL,包含2.0 μL 50 ng·μL-1的DNA,2.5 μL 2×KASP Master Mix,0.056 μL引物混合液,用ddH2O补充至5 μL。两个上游引物和共用下游引物的浓度分别为12 μmol·L-1和30 μmol·L-1。PCR反应程序为:94 ℃变性15 min;94 ℃变性20 s,65~55 ℃退火/延伸1 min,10个循环(从第二个循环开始每个循环降1 ℃);94 ℃变性20 s,57 ℃退火/延伸1 min,32个循环。用Synergy H1型多功能酶标仪(BioTek,美国)读取荧光值,用KlusterCaller软件(KBioscience)分型。若荧光信号弱、分群较分散,则影响数据分析,可以加3个循环(94 ℃变性20 s,57 ℃退火/延伸1 min),直至结果满意为止。

表2 TaGS5-A1位点、染色体、等位基因及引物序列Table 2 TaGS5-A1 locus,chromosome,alleles and primer sequences for detecting allelic variations
1.5 统计分析
用Excel 2010对每种基因型对应小麦品种的千粒重、粒长、粒宽、株高、穗长、穗下节长平均值分别进行统计分析。用SPSS 20.0对上述性状3年2点数据[44]进行方差分析。KASP结果判读中排除DNA 样本检测显示为缺失的数据,不予分析。采用t测验进行TaGS5-A1位点不同等位变异的显著性差异分析。
2 结果与分析
2.1 籽粒等主要农艺性状的表型分析结果
从表3可以看出,基因型、环境及基因型与环境互作对所有检测性状均有极显著影响(P< 0.01),说明基因型、环境及基因型与环境互作均影响籽粒相关农艺性状。

表3 籽粒及穗部部分性状的方差分析Table 3 Analysis of variance for grain and spike related traits
2.2 119个小麦品种 TaGS5-A1基因等位变异的分布频率
从图1可以看出,含有TaGS5-A1b等位基因的品种有91个,分布频率为76.5%;含有TaGS5-A1a等位基因的品种有28个,分布频率为23.5%,等位基因TaGS5-A1b的分布频率明显高于TaGS5-A1a。
2.3 TaGS5-A1基因等位变异的效应分析
从表4可以看出,含有TaGS5-A1b等位基因品种与含有TaGS5-A1a等位基因品种间的千粒重和粒宽差异均达到极显著水平,前者较后者增幅分别为6.6%和3.0%,说明TaGS5-A1b等位变异可增加小麦籽粒的粒重和粒宽。此外,含有TaGS5-A1b等位基因品种较含TaGS5-A1a基因的株高显著降低,降幅为6.6%,但其粒长(7.0 mm)、穗长(9.3 cm)、粒长/粒宽(2.0)和穗下节长(26.8 cm)均与含有TaGS5-A1a等位基因的品种间无显著差异,说明TaGS5-A1主要是通过增加粒宽来提高千粒重。
2.4 黄淮麦区不同省份小麦品种 TaGS5-A1基因等位变异的分布频率
对黄淮麦区河北(9个)、河南(42个)、山东(22个)、安徽(9个)、陕西(15个)小麦品种TaGS5-A1基因等位变异的分布频率进行分析,结果(表5)表明,TaGS5-A1b在安徽、河南、山东小麦品种的分布频率分别为100%、88.1%、 81.8%,在河北和陕西小麦品种的分布频率均为 66.7%。河北、安徽、陕西三省的小麦品种数目有限,统计结果仅供参考。但总的来看,TaGS5-A1b等位变异在黄淮麦区5省的分布频率明显高于TaGS5-A1a。

表4 TaGS5-A1基因等位变异品种间粒型等农艺性状的比较Table 4 Comparison of grain shape and agronomic traits between TaGS5-A1a and TaGS5-A1b allele varieties
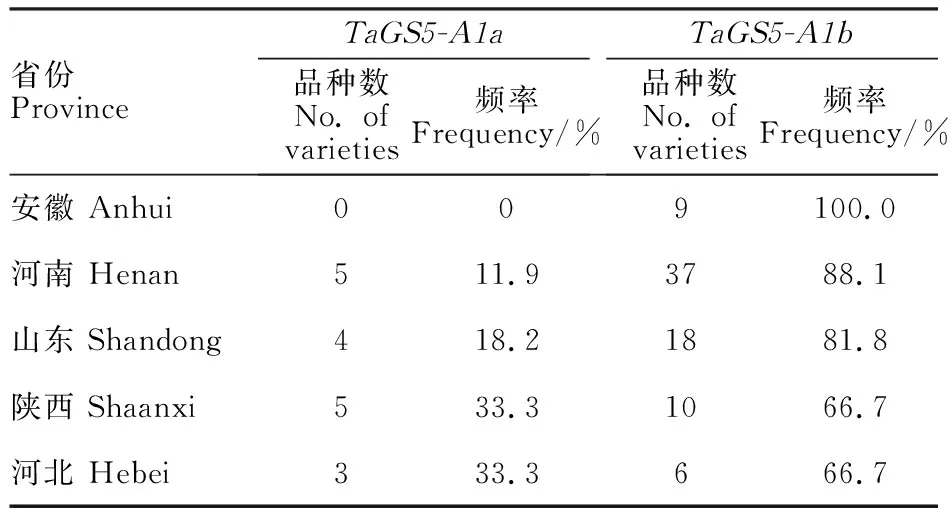
表5 黄淮麦区不同省份小麦品种 TaGS5-A1基因等位变异的分布频率Table 5 Allelic frequencies at TaGS5-A1 locus in wheat varieties from different provinces in Huanghuai wheat area
3 讨 论
3.1 TaGS5-A1基因KASP标记的验证
近十年来,模式植物水稻和拟南芥中与籽粒大小相关的遗传研究蓬勃发展。通过正向遗传学手段如数量性状位点的图位克隆和T-DNA插入突变体库已克隆出40多个控制籽粒大小的基因[46]。这些基因主要与三个遗传途径有关,包括蛋白酶体的降解,G蛋白信号和植物激素[47-49]。控制粒形的基因大多调控籽粒的细胞分裂,而控制籽粒灌浆的基因则主要影响源库关系。在细胞水平,GS5基因主要通过促进细胞分裂来增加细胞数目,进而调控籽粒大小和粒重[35]。
功能标记作为育种应用的理想标记,经验证优化后可用于所有育种材料的检测和分子标记辅助选择[50]。本研究利用粒重基因TaGS5-A1的KASP标记对黄淮麦区119个小麦品种该位点等位变异分布及效应进行分析,发现TaGS5-A1b等位基因的分布频率达76.5%,在6个环境中携带该等位基因的小麦品种具较宽的粒宽和更高的千粒重(P<0.01),这与采用该基因CAPS标记的分析结果一致[37-38],验证了TaGS5-A1基因的KASP标记的有效性和实用性,可用于小麦粒重的辅助选择。
Wang等[13]将TaGS5-A1基因定位到3AS染色体上,其侧翼标记为Barc356 和 Barc324,其中携带TaGS5-A1a等位基因材料的株高、穗长、穗下节显著高于含有TaGS5-A1b基因型的材料(P<0.05)。本研究携带TaGS5-A1b的材料株高显著低于含有TaGS5-A1a的材料,这与前人研究结果一致,但两个等位基因材料间的穗长、穗下节长无显著差异。已有研究表明,3A 染色体上与小麦千粒重相关的QTL与SSR分子标记 Barc356 紧密连锁[51],该染色体上与株高有关的 QTL位于分子标记 Barc324和 Xwmc428之间[52]。类似地,定位于2BL染色体上控制产量和株高的QTLs区域含有粒重基因TaSus2-2B,且携带该基因优势等位变异Hap-H的小麦品种在2个环境下株高均显著降低[11]。由于控制小麦粒重的主效QTL或基因经常与其他性状紧密连锁,存在一因多效的现象,那么本研究中粒重基因TaGS5-A1是否与株高QTL紧密连锁,还有待于进一步验证。
粒重性状容易受气候、土壤、生产条件和栽培措施等因素的影响,造成不同地区不同年份籽粒质地、重量差异较大。本研究通过多年多点试验,获得连续3个生长周期共6个环境的表型数据,更准确地评价了粒重基因TaGS5-A1的效应及其KASP标记辅助选择的应用价值。
3.2 TaGS5-A1b等位基因的分布频率
本研究发现黄淮麦区小麦各主产省份TaGS5-A1b等位基因的分布频率不同,但总体趋势一致,表明该粒重基因优异等位变异在黄淮麦区不同省份均被不同程度地正向选择,但是TaGS5-A1b等位基因的粒重改进潜力在不同省份有所差异。陕西省携带TaGS5-A1b等位基因小麦品种的粒重(41.9 g)高于TaGS5-A1a等位基因(39.2 g)的品种,可能是品种数偏少所致;其粒宽(3.4 mm)显著高于TaGS5-A1a基因型材料(3.3 mm),表明陕西省品种粒重改进潜力大。山东省选育的携带优势等位变异TaGS5-A1b小麦品种的千粒重(45.5 g)、粒宽(3.5 mm)均显著高于TaGS5-A1a基因型(41.2 g,3.4 mm),进一步验证了TaGS5-A1b基因型与千粒重显著正相关,而且与宋健民等[53]发现山东省1999-2010年育成的小麦品种产量构成要素中粒重呈显著上升结果相印证。
3.3 TaGS5-A1b基因功能标记在育种中的应用
粒重作为多基因控制的数量性状,以表型为主的选择可能造成优异基因的丢失,而以分子标记跟踪能确保优异基因的保留。所以常规育种与分子标记辅助选择相结合可提高对粒重选择的准确性和育种效率[54]。目前国内学者利用粒重基因特异性分子标记快速准确鉴定出不同小麦品种中粒重形成相关基因及其变异类型的分布,为小麦高产育种过程中亲本选配和后代辅助选择提供依据[8]。本研究主要基于TaGS5-A1基因对粒重的增效进行了验证,明确携带优异等位变异TaGS5-A1b小麦品种的粒重较高。粒重较高的品种如中麦875(52.8 g)、豫麦34(50.8 g)、鲁麦23(50.7 g)和中麦895(49.8 g)均携带与其高粒重表型一致的优异等位变异TaGS5-A1b。研究发现,中麦895还具有4个位点的优异等位变异,即Hap-A、Hap1/2/3、Tabas1-B1a和TaCwi-A1a;豫麦34还具有3个位点的优异等位变异Hap-A、Hap1/2/3和TaCwi-A1a;中麦875还具有2个位点的优异等位变异Hap-A和Hap1/2/3;鲁麦23仅有1个位点的优异等位变异,为Hap-A。以上小麦品种应该还存在其他粒重基因的优异等位变异,需要进一步明确这些高粒重品种中影响粒重的变异位点,评价相关位点的高通量KASP标记。因此,今后在利用本研究筛选的粒重基因TaGS5-A1高通量KASP标记时,应在综合考虑基因、环境以及植株本身优良性状的基础上,结合其他多个粒重和品质基因标记进行基因聚合,可加速培育出具有突破性的高产优质小麦新品种。
猜你喜欢
麦类作物学报(2022年6期)2022-08-09
中国农学通报(2022年13期)2022-05-31
现代畜牧科技(2021年4期)2021-12-05
智慧健康(2021年17期)2021-07-30
山东建筑大学学报(2020年5期)2020-09-16
现代农业研究(2019年3期)2019-06-05
新课程·下旬(2018年9期)2018-11-14
吉林农业(2017年7期)2017-07-12
科学种养(2017年6期)2017-06-13
青少年科技博览(中学版)(2015年10期)2015-01-11
