
超亲优势数量遗传学理论推导及其在玉米育种中的应用
2021-08-05 13:51陈泽辉王安贵祝云芳郭向阳刘鹏飞
西南农业学报 2021年6期
陈泽辉,吴 迅,王安贵,祝云芳,郭向阳,刘鹏飞
(贵州省农业科学院旱粮研究所,贵州 贵阳 550006)
【研究意义】杂种优势是遗传基础不同的2个亲本的杂交一代在生长势、存活力、生殖力和抗性、品质等方面均优于双亲的现象[1]。杂种优势是非常重要的生物学现象,超亲优势则作为一种杂种优势表现形式,被广泛用于生产实践,但对其认识仍处于不断加深中,对超亲优势数量遗传学理论进行推导并应用于实践,对丰富玉米育种理论具有重要的意义。【前人研究进展】我国北魏《齐民要术》一书对马和驴杂交产生骡的现象进行了详细记载,骡表现出较马和驴强壮和高大,且骡的寿命比马和驴都长。这种子代表现超过亲本的现象很好地体现了超亲优势,为人类认识和利用超亲优势开辟了成功案例。在农业上,杂种优势已被广泛应用于水稻、玉米、高粱、油菜等植物及家畜和家禽等动物育种上,并已取得了明显效果。【本研究切入点】关于杂种优势形成的遗传基础较为复杂,其理论研究较多,对杂种优势理论的探索却远远落后于应用实践,杂种优势形成的遗传基础仍停留在假说阶段。关于杂种优势理论尤其是超亲优势的认识仍然处于不断加深中。【拟解决的关键问题】在综述前者研究成果的基础上,结合多年的玉米自交系和杂交种选育实践,基于加性-显性模型对超亲优势数学表达式进行推导,并对超亲优势可能的遗传基础及其在指导玉米自交系和杂交种选育方面的应用进行探讨,以期提高玉米育种效率,为玉米育种的理论指导提供参考。
1 超亲优势数量遗传学理论推导
1.1 中亲优势的数量遗传学理论
数量性状是由多个位点基因控制,往往表现出很强的杂种优势现象。为推导出杂种优势的数学表达式,以具有2个等位基因G和g的单个位点为例,A和B两个随机交配群体,杂交产生的F1往往表现出杂种优势,其中一个群体等位基因G和g的频率分别是p和q;另一个群体同一位点上的等位基因频率分别为p′和q′;两群体间同一位点同一等位基因频率之差为y,则y=p-p′=q′-q。把p′和q′写作p-y和p+y。按基因型尺度(图1)理论,GG、Gg和gg的基因型值分别为a、d和-a[2-3]。
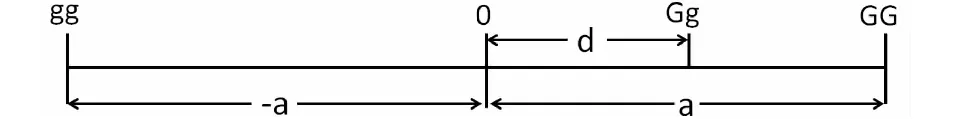
图1 基因型尺度
按基因型尺度,A群体的平均值:
MPA=a(p-q)+2dpq
B群体的平均值:
MPB=a(p-y-q-y)+2d(p-y)(q+y)=a(p-q-2y)+2d[pq+y(p-q)-y2]
即2个群体杂交产生F1时,从一个群体随机得到的个体与从另一个群体随机得到的个体交配获得F1的基因型频率列于表1。

表1 2个群体杂交后代的基因频率和基因型频率

表2 在2个Hardy-Weinberg平衡群体杂交组合1个位点2个等位基因下的基因型值和基因型频率
F1的平均基因型值:
MF1=a(p2-py-q2-qy)+d[2pq+y(p-q)]=a(p-q-y)+d[2pq+y(p-q)]
中亲优势值的度量表示在F1与中亲值间的差值,则:
式中,d为显性效应,若该位点无显性,则不会产生杂种优势。Y为两群体该位点基因频率之差,若两群体间基因频率无差异,则不会产生杂种优势。考虑两群体与研究性状有关的所有位点的联合效应,不考虑位点间的上位效应,即,在加性-显性模型下,F1总的中亲优势如下。
1.2 超亲优势的数量遗传学理论
超亲优势指杂交组合F1的表现高于高值亲本或低于低值亲本的现象。超显性假说由Shull[4]提出,经East[5]利用基因理论将此观点具体化。按基因型尺度模型导出超亲优势的数学表达式。
1.2.1 超高值亲本的超亲优势 假定A群体为高值亲本,B群体为低值亲本。在单个位点情况下,超高值亲本的超亲优势数学推导如下。
mPA=a(p-q)+2dpq
MPB=a(p-q-2y)+2d[pq+y(p-q)-y2]
MF1=a(p-q-y)+d[2pq+y(p-q)]
HPHF1=MF1-MPA=a(p-q-y)+d[2pg+y(p-q)]-a(p-q)-2dpq=a(p-q-y-p+q)+d[2pg+y(p-q)-2dpq]=-ay+yd(p-q)=y[d(p-q)-a]
若双亲均为纯系(纯合自交系),p为1,q为0,则超高值亲本的超亲优势如下:
HPHF1=a(1-y)+dy-a=a-ay+dy=y(d-a)
从上式可看出,在单个位点时,超亲优势等于双亲同一个基因的频率之差y与该位点显性效应d和加性效应a之差的乘积。若y为0,若双亲是均含该基因的纯合系,该位点无超亲优势。y为1时,即亲本1含该基因,亲本2不含该基因,超高值亲本的超亲优势如下:
HPHF1=d-a
从上式可知,在单个位点时,只有显性效应d大于加性效应a才有超亲优势。
1.2.2 超低值亲本的超亲优势 在单个位点情况下,超低值亲本的超亲优势数学推导如下:
LPHF1=MF1-MP2=a(p-q-y)+d[2pq+y(p-q)]-[a(p-q-2y)+2d[pq+y(p-q)-y2]=a(p-q-y-p+q+2y)+d[2pq+y(p-q)-2pq-2y(p-q)+2y2]=ay-dy(p-q)+2dy2
LPHF1=MF1-MPB=a(p-q-y)+d[2pq+y(p-q)]-[a(p-q-2y)+2d[pq+y(p-q)-y2]=a(p-q-y-p+q+2y)+d[2pq+y(p-q)-2pq-2y(p-q)+2y2]=ay-dy(p-q)+2dy2
若双亲均为纯系(纯合自交系),p为1,q为0,则:
LPHF1=ay-dy+2dy2
y为1时,即亲本A群体含该基因,亲本B群体不含该基因,那么,超低值亲本的超亲优势如下:
LPHF1=a+b
从上式得出,在单个位点时,只有显性效应d的绝对值大于加性效应a才有超低值亲本的超亲优势。
双亲的基因频率差异y为0。
LPHF1=ay-dy+2dy2=0
1.2.3 多位点下的超亲优势 考虑两群体与研究性状有关的所有位点的联合效应,不考虑位点间的上位效应,即在加性-显性模型下,F1总的中亲优势为多个位点超亲优势的累加。
数量性状由多个位点基因控制,如图2中A和G位点。A位点内A基因和a基因与G位点内G基因和g基因间的相互作用为上位性互作关系,多数情况下位点间的上位性不显著,或占比较小。当上位性互作为正时,优势会增大,当上位性互作为负时,优势会降低。超高值亲本的超亲优势数学表达式HPHF1=Σy(d-a),及超低值亲本的超亲优势数学表达式LPHF1=Σ(d+a),在加性-显性模型条件下这2个数学表达式才成立,但上位性可能对杂种优势表现有所贡献。在无上位性互作情况下,只有在多个位点的(d-a)的代数和为正时,才有超亲优势。则需要多个位点的显性效应是超显性效应时,杂种F1才会表现出超亲优势。说明,超亲优势主要是超显性在起作用。
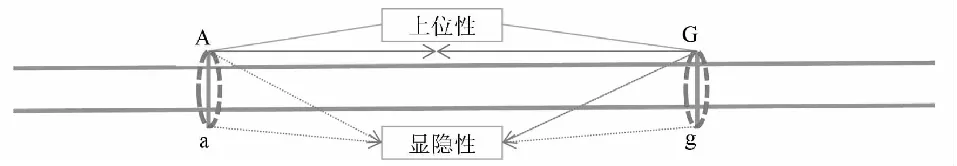
A位点内A基因与a基因间及G位点内G基因与g基因间的相互作用均为显隐性关系,A位点内A基因和a基因与G位点内G基因和g基因间的相互作用为上位性互作关系
上位性效应在杂种优势表现中存在潜在作用[6-7]。但是上位性互作效应只能解释玉米群体中微不足道的遗传变异,而加性和显性方差能够解释除此之外的大部分遗传变异。上位性似乎并不是遗传变异的主要成分,上位性应该表现在由优良自交系组成的特定杂交种中,因此,上位性的相互作用似乎在特定的,而不是在杂交组合间普遍存在[8]。
Hallauer等[8]为获得加性a、显性d组分和3种上位性组分(aa、ad和dd)的方差,利用从玉米群体BSSS中获得的一系列随机自交系进行的研究结果表明,加性组分占总产量遗传方差的93%;加性与显性共占99%。说明,上位性对产量的遗传变异为次要贡献因素[9]。
考虑到上位性对杂种优势可能存在的影响,超亲优势的数学表达式为HPHF1≈Σy(d-a)和LPHF1≈Σ(d+a)。除基因频率之差外,还考虑双亲互补性,在此基础上形成杂种优势的基因频率差异互补的理论观点,可用于指导玉米育种实践。
如具有4个位点开展的数量性状,不同基因型AABBccdd和aabbCCDD的2个自交系杂交,F1的基因型为AaBbCcDd。假设显性基因A对某一数量性状的贡献为12,B的贡献为10,C的贡献为8,D的贡献为6;相应的隐性等位基因的贡献分别为6、5、4、3。在不存在上位性时,亲本AABBccdd的表现型值为12+10+4+3=29,另一亲本aabbCCDD的表现型值为6+5+8+6=25。根据显性基因的效应可知F1的表现型值。若无显性效应,杂合的等位基因Aa、Bb、Cc、Dd的贡献值均等于相应的等位显性基因和隐性基因的平均值,(12+6+10+5+8+4+6+3)/2=27,与双亲的平均值相同,无杂种优势。若表现部分显性,则P1表现型值大于中亲值偏向高值亲本,表现出部分杂种优势,即AaBbCcDd>27。若表现为完全显性,则F1大于高值亲本,杂种F1为AaBbCcDd=12+10+8+6=36,表现出超亲杂种优势。利用超亲优势的数学表达式HPHF1=y[d(p-q)-a]分别计算各位点的超亲优势。
对A位点,双亲平均值m=9,a=3,d=3,y=1,p=1,q=0,超亲优势:HPHF1=1×[3(1-0)-3]=0,F1=P1+HPHF1=12+0=12,对B位点,双亲平均值m=7.5,a=2.5,d=2.5,y=1,p=1,q=0,超亲优势:HPHF1=1×[2.5(1-0)-2.5]=0,F1=P1+HPHF1=10+0=10,对C位点,双亲平均值m=6,a=2,d=2,y=-1,p=0,q=1,超亲优势:HPHF1=-1×[2(0-1)-2]=4,F1=P1+HPHF1=4+4=8,对D位点,双亲平均值m=4.5,a=1.5,d=1.5,y=-1,p=0,q=1,超亲优势:HPHF1=-1×[1.5(0-1)-1.5]=3,F1=P1+HPHF1=3+3=6。
4个位点总的超亲优势:
HPHF1=∑y[d(p-q)-a]=(0+0+4+3)=7
基因型AaBbCcDd的表现:
总的F1=12+10+8+6=36,从单个位点看,超亲优势的遗传基础是超显性效应。多个位点控制的数量性状,显性效应对超亲优势的表现起主导作用,其中包括超显性和完全显性,或者部分显性;不同位点的显性程度可能不一,一个位点可能是显性效应,另一位点可能是超显性或部分显性。另外,显性效应的方向也可能不一,总的效应是多个位点显性方向的代数和。
与中亲优势一样,超亲优势大小与双亲基因频率有关,在显性效应和加性效应相对固定时,双亲基因频率差异越大,超亲优势越强,这对指导玉米自交系选育和杂交组合组配具有重要意义。
2 超亲优势理论在作物杂种优势上的应用
2.1 几种基因作用方式对杂交种表现的影响
在单个位点,杂种F1的表现等于高值亲本群体的平均值,加上中亲优势。若在多个位点情况下,可能存在上位性。因此几种基因作用方式,即加性效应、显性效应和上位性对杂交种的表现有重要影响。
基因的加性对双亲群体的平均值起作决定作用。超亲优势HPHF1≈Σy(d-a)和LPHF1≈Σ(d+a),含有加性效应a。一个群体或自交系的一般配合力主要由基因的加性效应决定。因此,在作物育种中通过测定一般配合力来确定材料(群体、自交系等)具有加性基因及其效应的多少。反之,要提高一个纯系或自交系的一般配合力,则需用优势强的杂交组合进行二环选系、复合杂交选系和轮回选择后选系等方法来提高优良基因数量,提高总的遗传效应。即在一个自交系中集中更多的加性基因来提高亲本自交系的平均值[2]。
经验证据已表明,自交系的相对活力、抗性、产量潜力等在连续多个循环的选择中不断增加,此为美国玉米带单交种成功的原因[10],玉米二环系的产量较一环系高30.8%。另外,Collins[11]研究表明,若控制一个性状的因子对数太多,则很难获得所有基因都纯合的植株。控制一些性状,如活力、抗性、产量等的基因数量未知,但可以肯定其数目很大。在玉米育种实践中,从整个基因组的角度看,双亲间的基因差异不大。所以育种上尽可能用双亲基因差异大的自交系组配杂交组合,达到杂种优势最大化,则需建立杂种优势群。玉米育种的不同杂种优势群体是人为创造和保持的,不同国家以及同一国家的不同育种单位所采用的杂交优势群不一。在育种实践中,育种者有固定的杂种优势群是首要,并在相应优势群下对双亲不断互为改良,才能不断地培育出好品种,这一点尤其在正确利用二环系选择中具有指导意义[12]。
在孟德尔遗传中,显性是非常重要的遗传现象。显性遗传有完全显性、部分显性和超显性,呈正向显性和负向显性。显性效应的大小和方向,既因作物不同,同一作物不同性状而不同,也因控制数量性状的不同位点而不同。但同一位点,显性效应大小和方向是相对固定的。同一数量性状有关位点的定向显性,若不同位点显性方向不同,则会相互抵消,削弱总的显性表现。
显性效应的重要性因组合和性状而有很大变化。在中亲优势的数学表达式MPHF1≈Σdy2中,显性效应对中亲优势的出现非常重要,无显性效应,就无杂种优势,显性效应的正负决定杂种优势的方向。在双亲均为纯系时,各位点的中亲优势等于显性效应。超亲优势的数学表达式:HPHF1≈Σy(d-a)和LPHF1≈Σ(d+a),超显性对超亲优势起决定作用。
上位性是指不同位点基因间的相互作用,可能是2个位点,也可能是3个或多个位点基因间的相互作用。上位性效应极其复杂,具体效应值很难估算。因此,在数量遗传的研究模型中,多假定无上位性或上位性不显著;若上位性显著,也简单采用加性×加性、显性×显性和加性×显性进行分类,很难确定是哪些基因间的相互作用及效应值大小。Hallauer[8]解释上位性效应在育种中的影响认为,若上位性方差的比例较小,则可忽略上位性偏差对选择进度的影响。
2.2 超亲优势理论在作物杂种优势上的应用
超亲优势的数学表达式HPHF1≈Σy(d-a)和LPHF1≈Σ(d+a)。由于多数情况下,上位性对性状表现的作用相对较小,且上位性作用相当复杂,在此重点考虑加性和显性效应,及双亲基因频率差异对作物杂交种表现的影响,从而指导育种实践。
2.2.1 基因频率差异大或显性程度高能够提高杂种优势 要提高杂种优势,须增加双亲间基因频率差异和寻找显性程度高的基因位点。除利用商业栽培品种外,从基础群体进行选系是一个重要途径。很多育种材料可被育种者作为半成品引进,大多源自农家品种或传统的原始栽培品种,这些原始栽培品种经过数百年甚至上千年的演化被自然和人为选择所驯化,其内部多样性丰富。玉米起源于热带亚热带地区,当地丰富种质资源是实现现代玉米产业发展目标的重要育种材料来源,特别是经CIMMYT等机构改良过可应用于生产的各类品种。此外,充分利用与栽培品种关系较近的野生种为另一个重要途径。这些近缘野生种或是栽培作物的祖先,或与作物亲缘关系较近(大刍草为典型例子),这些种质资源正变得日益重要。
2.2.2 两群双向选择育种模式 利用中亲优势MPHF1≈Σdy2和超亲优势HPHF1≈Σy(d-a),以有利显性基因分散在双亲群体中,这一理论能较好地指导玉米自交系和杂交种的选育[13]。
(1)玉米杂种优势类群划分与利用。美国玉米育种者根据自交系的来源把自交系划分为Reid和Lancaster 2个优势群,发现两者间有很强的杂种优势,形成了玉米研究中最典型的杂种优势利用模式。迄今为止,美国利用时间最长、使用范围最广的仍然是这2个杂种优势群及由其组成的杂种优势模式。欧洲最常用的模式为美国马齿型×欧洲硬粒型;在热带地区则以ETO复合品种×Tuxpeňo为主要模式;巴西以马齿型×Tuxpeňo或Suwan1×Tuxpeňo 为常用模式。
国内主要杂交种的亲本自交系大致可分为7个杂种优势类群:唐四平头群、旅大红骨群、改良Reid群、Lancaster群、PA群、PB群、Suwan群和Tuxpeňo群等[14]。分别从这些类群中选育自交系,然后类群间自交系杂交,获得强优势杂种组合的频率较高。西南地区主要的玉米杂种优势模式:改良Reid群×Suwan群、改良Reid群×PB群、Tuxpeňo群×Lancaster群和Suwan1×Tuxpeňo等。
(2)两群相互轮回选择模式及其利用。两群相互轮回选择,即2个群体互为测验种,不仅改良2个群体本身,更重要的是改良了群体间杂交组合的表现。该改良方法是基于杂种优势的数学表达式,重点是增加2个群体间基因频率差异,既增加了2个群体的加性效应,也提高了群体间非加性基因效应。群体的一般配合力和2个群体间的特殊配合力均得到提高。
Chen等[15]对温热群体墨瑞(TR)和苏兰(SL)进行3轮相互轮回选择,群体间杂交组合的籽粒产量提高了13.89%,达当地推广杂交种(贵单8号)的产量水平(图3)。同时,群体间杂交组合的中亲优势从15%增至25.96%,超亲优势从10.91%增至16.49%(表3),而吐丝期表现为负向超亲优势。目前,已分别从墨瑞和苏兰改良群体中选育出自交系,并组配出优良组合。
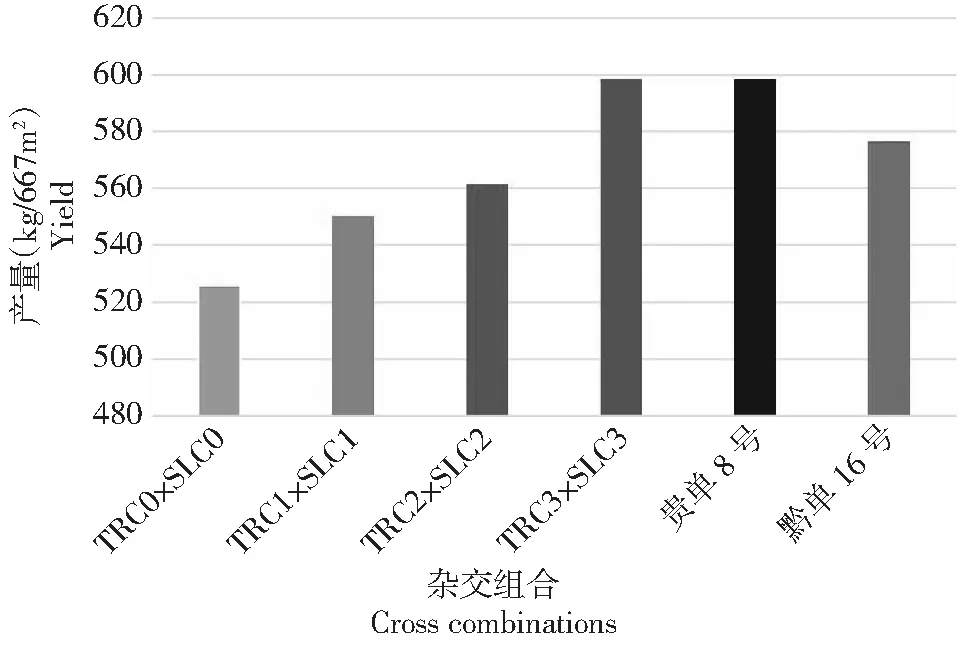
图3 玉米TR1群体×SL1群体相互轮回选择的籽粒产量
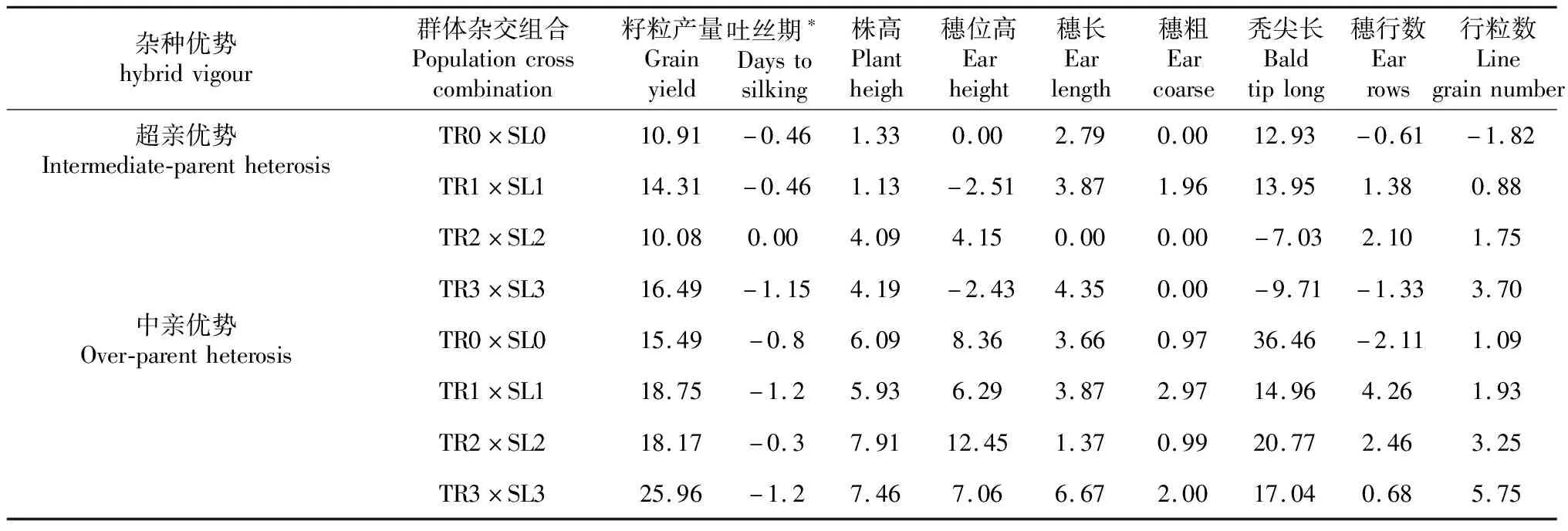
表3 玉米群体间杂交组合的超亲优势和中亲优势
(3)杂种优势的全基因组预测。基因组预测是利用训练群体的基因型结果和表型数据,建立最佳线性无偏预测模型,得到训练群体的基因组估计育种值(GEBV)。在此基础上,根据育种群体的基因型数据,基于BLUP模型和与训练群体中的等位基因同一性预测育种群体各性状表现值,从而得到各个体具有优良表型的估计值,即高育种值。研究表明,影响全基因组预测准确性的因素有用于估计基因组估计育种值的标记数、性状遗传力、参考群体大小、统计模型、分子标记的数量和类型、连锁不平衡、有效群体大小、参考和预测群体的关系和群体结构[16],其中训练群体的结构是影响多亲本群体基因型预测准确性的重要因素,且双向抽样能够有效提高基因组预测效率。基于两群体双向选择获得材料所具有的遗传独立性,它们间杂交会产生很强的杂种优势,所获得的后代杂交群体具有系谱清晰、遗传结构简单且遗传重组较双亲分离群体多的优点,利用该群体做训练群体,可降低标记密度。加上2个类群间杂交后代与类群亲本间所产生的同胞关系,可进一步减少对训练群体大小的要求。
2.2.3 群间选系组配模式及其利用 按杂种优势的数学表达式,2个不同类群间杂交F1的表现最强,说明集中了较多的有利显性基因,双亲差异大。若用F1杂交组合选育自交系,在多次混合授粉和加大后代群体种植数量,则有可能选育出集中更多优良基因的自交系,然后与其他类群自交系组配,育出优良组合的频率会增加,此为群间选系组配模式[2]。该模式的难点是在群间选系后,寻找可组配形成优势杂交组合类群,或改良创新类群。在不同的生态类型区,如黄淮海区、东华北区、西北区和西南区,改良创新类群和组配模式可能会有所变化。目前较成功的群间选系组配创新模式有[改良瑞德(BSSS)×Lancaster]×(黄改×旅系)、[改良瑞德(BSSS)×Lancaster]×(黄改×欧洲硬粒)、[改良瑞德(BSSS)×Lancaster]×Suwan1、[改良瑞德(BSSS)×Lancaster]×[Suwan1×Iodent]及[Tuxpeo×改良瑞德(BSSS)]×(Suwan1×Lancaster)等。
笔者所在团队用PB群的自交系9195与属于改良瑞德群的丹9046杂交,从F1自交后代中育出优良自交系QB506。QB506与属于Suwan1类群自交系T32和QR273组配,分别育出优良杂交种金玉818和金玉506,在西南地区被大面积推广应用。因此,(PB群×改良瑞德)× Suwan1是适宜西南地区玉米自交系和杂交种的选育模式。
3 讨 论
对杂种优势形成的机理研究显示,显性效应、超显性效应和上位性均可能对杂好优势有贡献,并且均有试验和实验数据支撑。但都不能很好地解释整个杂种优势的形成机理。
徐云碧[17]认为,杂合性极其相关的基因互作是解释杂种优势的主要遗传学基础,因为杂交种对于亲本间有差异的所有基因位点均为杂合的。因此,杂种优势的程度取决于何种基因位点是杂合的,以及基因位点内等位基因和基因位点间等位基因彼此间如何互作。基因位点内等位基因的互作导致显性、部分显性或超显性,理论上的显性程度为0(无显性)至大于1(超显性)。基因位点间等位基因的互作导致上位性。遗传作图结果表明,杂种优势和其他数量性状中涉及的大多数QTL具有显性效应,随着能够更有效地估计上位性方法的出现,上位性已被更频繁地发现,并被证明是数量性状遗传控制中的一个普遍现象。然而,对于不同杂交组合、物种或性状,其杂种优势须通过不同程度的显性结合所有可能的基因位点间互作来解释。对杂种优势的充分理解将依赖于与杂种优势有关的所有基因的克隆和功能分析。
现今使用的杂交种是通过杂交2个高度近交的系,即自交系而产生的F1杂种。无论采取何种育种方式,关注点都集中在F1代比2个亲本更优。多年来提出的4种解释:一是2个亲本间显性或部分显性基因的分散;二是在各个基因位点上的超显性;三是2个基因位点相斥相有利等位基因紧密连锁而导致的拟超显性;四是上位性[18]。这些解释相互不排斥,因此需确定其在任何特定杂交种中的相对重要性,但实践证明很难确定,说明这些遗传成分在杂种优势形成的遗传基础上不是相互排斥的。因而杂种优势形成虽有多个假说,但一直存在争议[19]。
随着对杂种优势研究的深入,对杂种优势的理解越来越复杂,很难将已有研究结果应用于育种实践。通过对前人研究结果的分析认为,对杂种优势的形成,基因的加性效应、显性效应、超显性和位点间基因的上位性都有作用,会因材料遗传背景不同、位点不同、基因不同而在这3个方面的基因作用方式上存在差异。基因的加性效应和显性效应普遍存在,而位点间的上位性因特定的材料和位点才显著存在,并非普遍现象;或是与加性效应和显性效应相比,其效应相对较低,大多数情况下似乎可以忽略不计。
为玉米指导育种实践,对杂种优势的理解应尽可能简单化,抓主要的或普遍存在的,并且在遗传方差成分中占比较大的部分。杂种优势利用的最终目的是育出产量高的F1杂交种,因此,加性效应的重要性是第一位的,它决定双亲的平均值;其次,显性效应也非常重要,决定F1杂交种的杂种优势,不管是中亲优势还是超亲优势,是显性效应还是超显性起作用,均是不同性状或是同一性状不同位点的差别而已。上位性对杂种优势的影响不可预见,且随遗传背景的差异,在育种中可以暂时忽略。双亲群体基因频率差异对杂种优势的影响显而易见,如基因频率差异y为零,则无杂种优势;相反,y值达最大的1时,杂种优势最大。因此认为,中亲优势和超超亲优势的数学表达式MPHF1≈Σdy2和HPHF1≈Σy(d-a)能很好地用于指导玉米育种实践。
4 结 论
在综述前者研究成果的基础上,结合多年的玉米自交系和杂交种选育实践,对超亲优势数量遗传学理论的推导结果显示,基因型尺度模型下的中亲优势和超亲优势的数学表达式分别为MPHF1≈Σdy2及HPHF1≈Σy(d-a),均能很好地用于指导玉米育种实践。
猜你喜欢
中国种业(2022年8期)2022-08-19
西南农业学报(2022年5期)2022-06-06
热带亚热带植物学报(2022年2期)2022-04-14
蔬菜(2021年7期)2021-11-27
种子(2021年8期)2021-09-23
种子(2021年7期)2021-08-19
广东农业科学(2021年3期)2021-04-23
热带农业科技(2019年1期)2019-01-14
农民致富之友(2016年5期)2016-10-21
江苏农业科学(2015年1期)2015-04-17
