
不同栽植年限小叶杨与胡杨杂交种(小×胡杨)光合响应特性比较
2021-08-16 01:50黄雅茹马迎宾迟悦春赵英铭刘禹廷
西北林学院学报 2021年4期
黄雅茹,马迎宾,迟悦春,赵英铭,褚 洋,孙 非,刘禹廷,张 帅
(1.内蒙古磴口荒漠生态系统国家定位观测研究站/中国林业科学研究院 沙漠林业实验中心,内蒙古 磴口 015200; 2.中国林业科学研究院 华北林业实验中心,北京 102300)
光照对于植物生长发育具有非常重要的影响,如果植物长期生长在不同的光强下,即使是相同的植物,为了适应不同的光环境,其形态指标、生理指标也会发生相应的改变。植物生长发育、生物量的积累、有机物质的积累重要基础是光合作用[1],光合作用随着光强的变化而变化。光照强度增加,光合速率也增强,反之亦然,因此光照强度显著影响植物的光合作用[2-3]。光合作用的生理过程非常复杂,植物自身生理特性和环境因素共同影响光合作用,光合作用的过程时刻受光照、温度、水分等环境因子的影响,其中,光合作用的先决条件是光照[4]。测定光合作用后,通过公式计算求出光饱和速率(Amax)、光饱和点(LSP)、光补偿点(LCP)、表观量子效率(AQY)、暗呼吸效率(RD)等生理参数[5]。研究表示,通过上述生理参数有助于判定植物光合作用能力和光合作用效率及其受环境变化的影响程度[6-7]。在干旱区,植物的水分生理及光合生理特性与人们采取的植被管理措施密切相关,因此,植物的水分生理及光合特性直接决定了植物的生存和发展状况[8]。了解荒漠植物在特定生境中的光合以及水分生理特性非常重要。
乌兰布和沙漠绿洲是内蒙古河套地区防风固沙的重要屏障,目前,防护林体系主要的造林树种有新疆杨(Populusalbavar.pyramidalis)、二白杨(Populusgansuensis)、小叶杨(Populussimonii)、毛白杨(Populustomentosa)等,但是,绿洲防护林的部分退化引发了一些环境问题及社会矛盾,因此,林业部门对防护林体系的更新与重建非常重视,尤其是树种选择问题,是目前急需解决的难题[9]。我国干旱沙漠地区乔木造林树种非常少,是生态建设中亟待解决的重大技术问题;而用干旱沙漠区特有的树种材料,通过杂交育种的方式,选育出适宜干旱荒漠区的高抗性新品种是我国的重要课题。小×胡杨(Populussimonii×P.euphratica)是小叶杨(Populussimonii)和胡杨 (Populuseuphratica)进行多次杂交试验选育出的优良品种;小叶杨(♀)和胡杨(♂)有性杂交后,完全具有双亲融合性状[10]。前人研究显示,小×胡杨比小叶杨和胡杨生长迅速,耐盐碱性强于小叶杨,比胡杨更容易进行繁殖,是干旱沙漠区防护林造林的新品种[10]。目前,有关小×胡杨的研究报道较少,主要包括小×胡杨幼苗抗旱性[10]、育种[10]、引种[10]、硬枝扦插技术[10]、无菌培养[11]方面的研究,而有关小×胡杨气体交换参数对光强的响应方面的研究报道还很缺乏。鉴于此,本研究以乌兰布和沙漠东北部不同栽植年限的小×胡杨为研究对象,测定气体交换参数,分析不同光强对小×胡杨气体交换参数的影响,阐明其对环境光合特性的适应和生理响应,以期为乌兰布和沙漠东北部防护林体系的更新及造林树种选择提供一定的理论依据,为绿洲前沿生态环境的改善提供理论依据,为进一步评价小×胡杨新品种适应性提供基础数据。
1 材料与方法
1.1 研究区概况
研究区地理坐标为39°40′-41°00′N,106°00′-107°20′E,地形起伏不大,沙丘高度≤10 m,新月形沙丘或圆锥形沙丘是研究区的主要沙丘类型,沙质主要以细沙为主。研究区属于中温带半干旱大陆性气候,主要特点是云少,降水少、气候干燥,风沙活动比较多,风速较高。研究区多年平均降水量约140.3 mm(1954-2005年),沙漠昼夜温差大,平均气温为6.8℃,年日照时间为3 229.9 h。研究区主风向以西风和西北风为主,11月至翌年5月为研究区的风沙季节。土壤类型主要以风沙土为主[12]。人工林主要有柠条锦鸡儿(Caraganakorshinskii)、梭梭(Haloxylonammodendron)、花棒(Hedysarumscoparium)等,天然植被主要有油蒿(Artemisiaordosica)、白刺(Nitrariatangutorum)等,绿洲防护林体系主要栽植树种是杨树[13]。
1.2 试验材料
试验于2018年8月23日、24日进行,试验材料是小×胡杨(Populussimonii×P.euphratica),是小叶杨(Populussimonii)和胡杨 (Populuseuphratica)进行多次杂交试验选育出的优良品种,小叶杨(♀)和胡杨(♂)有性杂交后,完全具有双亲融合性状。试验以不同栽植年限的小×胡杨为研究对象,栽植立地条件一致,且栽植在同一林地。具体选择1年、2年、3年生的小×胡杨,对不同栽植年限的小×胡杨的生长状况进行测定,包括株高、胸径、冠幅、枝下高。
1.3 研究方法
不同栽植年限的小×胡杨叶片光响应曲线采用Li-6400xt便携式光合仪进行测定,在晴朗无云的天气,采用LED红蓝光源,1年、2年、3年生小×胡杨各选择3株为标准株,选择向阳无遮光部位的叶片,每株在冠层上、中、下各选择2片叶片,9:00-12:00进行测定。选取叶片为成熟且无病虫害的,CO2浓度设置为400 μmol·m-2·s-1,光强设置为1 500、1 200、900、600、300、100、50、0 μmol·m-2·s-1,重复3次。光响应曲线模拟是用光合助手软件(拟合植物叶片光合有效辐射(PAR)与净光合速率(Pn)的关系)。计算出小×胡杨的光合生理参数(光饱和速率,光补偿点及光饱和点,表观量子效率等)。非直角模型理论公式[14]为:
(1)
式中,A为表观量子效率,K为光响应曲线曲角,Q为光合有效辐射,Pnmax为净光合速率,RD为暗呼吸速率。
1.4 数据处理
数据整理及绘图使用Excel,采用函数STDEVP计算误差值。采用SPSS17.0进行多重比较分析。
2 结果与分析
2.1 不同栽植年限小×胡杨生长状况
3年生小×胡杨株高与胸径最大,分别是1年生的1.89、1.68倍,不同栽植年限树高与胸径存在显著差异(P<0.05),3年生小×胡杨冠幅也最大,与2年生差异不显著(P>0.05),与1年生差异显著(P<0.05)。3年生小×胡杨枝下高最大,与1年、2年生差异不显著(P>0.05)。
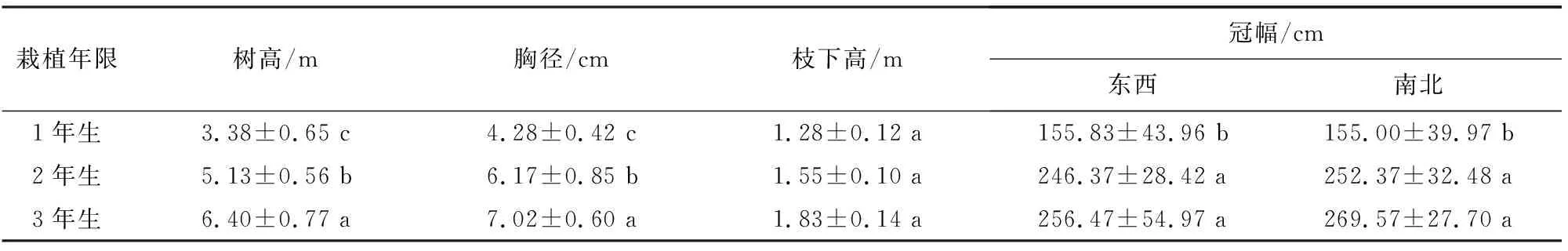
表1 不同栽植年限小×胡杨生长指标
2.2 不同栽植年限小×胡杨净光合速率光响应特征
植物光合能力的强弱是由光合速率来决定的,在一定的环境条件下,植物叶片的最大光合能力通过最大净光合速率来体现[14-15]。随着光合有效辐射的增加,不同栽植年限小×胡杨变化趋势一致。由图1可以看出,当光合有效辐射在0~900 μmol·m-2·s-1时,不同栽植年限小×胡杨净光合速率均随着光合有效辐射(PAR)的增强而迅速增加,当光合有效辐射(PAR)>900 μmol·m-2·s-1时,不同栽植年限小×胡杨净光合速率(Pn)随光合有效辐射(PAR)的增强缓慢增加,光响应曲线变化较为平缓。当光合有效辐射(PAR)为1 500 μmol·m-2·s-1时,曲线处于缓慢上升趋势,说明小×胡杨还没有达到光饱和点。经SPSS17.0方差分析与多重比较分析,1年、2年、3年生小×胡杨之间的净光合速率差异显著(P<0.05)。不同光合有效辐射,净光合速率值均表现为3年>2年>1年生。

图1 小×胡杨净光合速率光响应曲线
2.3 不同栽植年限小×胡杨光响应特征参数
光饱和速率(Amax)由高到低依次是3年>2年>1年生(表2)。光饱和点(LSP)用来表示植物利用光照强度的能力,光饱和点(LSP)值高,表明强光照射植物时,其生长发育不容易受到抑制,光补偿点(LCP)反映了植物利用弱光的能力,其值越小,说明植物利用弱光的能力越强,3年生小×胡杨LSP最高,说明强光照射时3年生小×胡杨生长发育不易受到抑制。1年生小×胡杨LCP最低,说明1年生小×胡杨利用弱光的能力越强。不同栽植年限小×胡杨的表观量子效率(AQY)从大到小依次是3年>2年>1年生。暗呼吸速率(RD)是指植物在没有光照条件下的呼吸速率,1年生小×胡杨的暗呼吸速率最大,3年生小×胡杨的暗呼吸速率最小,3年生小×胡杨的光饱和速率(Amax)、光饱和点(LSP)、光补偿点(LCP)、表观量子效率(AQY)均最高,不同栽植年限小×胡杨光饱和速率(Amax)差异达到显著(P<0.05),光饱和点(LSP)、光补偿点(LCP)、暗呼吸速率(RD)不显著(P>0.05),表观量子效率(AQY)2年生小×胡杨与3年生小×胡杨之间差异不显著(P>0.05),均与1年生小×胡杨差异显著(P<0.05)。
2.4 不同栽植年限小×胡杨蒸腾速率光响应特征
植物水分运输过程中,蒸腾作用起着主导作用。由图2可知,随着光合有效辐射的增加,不同栽植年限小×胡杨蒸腾速率缓慢增加,曲线趋势变化较为平缓。经SPSS17.0方差分析与多重比较分析,1年、2年、3年生小×胡杨之间的净光合速率光响应差异不显著(P>0.05)。不同光合有效辐射,蒸腾速率值均表现为3年>2年>1年生。

图2 小×胡杨蒸腾速率的光响应
2.5 不同栽植年限小×胡杨气孔导度和胞间CO2浓度的光响应特征
由图3可知,随着光合有效辐射的增加,1年、2年、3年生小×胡杨气孔导度(Gs)变化趋势一致,均呈增加趋势。不同光合有效辐射,气孔导度(Gs)值均表现为2年>3年>1年生。当光合有效辐射<600 μmol·m-2·s-1,1年、2年、3年生小×胡杨气孔导度(Gs)增加缓慢,当光合有效辐射>600 μmol·m-2·s-1,随着光强的增强,1年、2年、3年生小×胡杨气孔导度(Gs)增加较快。这说明光强的变化对小×胡杨气孔运动有着非常大的影响。随着光合有效辐射的增加,1年、2年、3年生小×胡杨气孔导度(Gs)变化趋势一致(图4),均呈下降趋势。当光合有效辐射<600 μmol·m-2·s-1,随着光强的增加,1年、2年、3年生小×胡杨胞间CO2浓度(Ci)下降较快,当光合有效辐射>600 μmol·m-2·s-1,随着光强的增加,1年、2年、3年生小×胡杨胞间CO2浓度(Ci)下降缓慢。
2.6 不同栽植年限小×胡杨水分利用效率光响应特征
在干旱荒漠地区,水分利用效率能够体现植物适应干旱环境能力的强弱。随着光合有效辐射的增加,1年、2年、3年生小×胡杨水分利用效率变化趋势一致,均呈增加趋势(图5)。光合有效辐射<600μmol·m-2·s-1,1年、2年、3年生小×胡杨水分利用效率增加迅速,>600 μmol·m-2·s-1时,随着光强的增强,3年生小×胡杨水分利用效率继续缓慢上升,但而1年、2年生的小×胡杨水分利用效率趋于平缓甚至下降。

表2 不同栽植年限小×胡杨光合作用光响应曲线的特征参数
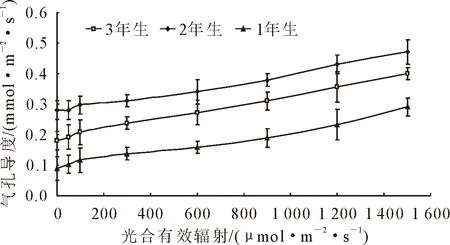
图3 小×胡杨气孔导度的光响应

图4 小×胡杨胞间CO2浓度的光响应
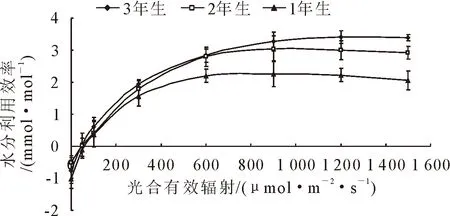
图5 小×胡杨水分利用效率的光响应
3 结论与讨论
3.1 讨论
植物生长发育和生存与光有着密切联系,植物光合作用所需能量的主要来源为光合有效辐射,光合有效辐射是影响植物光合作用重要因素[15]。表征植物光响应特性的指标主要包括光饱和速率(Amax),光饱和点(LSP)、光补偿点(LCP)等,也是研究植物光合作用的主要生理指标,同时也是研究植物适应性的重要参数[16]。不同栽植年限小×胡杨的光饱和点(LSP)变化范围1 722~1 763 μmol·m-2·s-1,比光补偿点(LCP)31.22~31.99 μmol·m-2·s-1的变化幅度大(表2),说明不同栽植年限小×胡杨进行光合作用利用强光的能力较强,利用弱光的能力较小。3年生小×胡杨的光饱和速率(Amax)最高,达到28.32 μmol·m-2·s-1,1年生小×胡杨的光饱和速率(Amax)为19.14 μmol·m-2·s-1,说明3年生小×胡杨比1年生小×胡杨利用强光进行光合作用的能力较强。该研究中小×胡杨的光补偿点为 31.22~31.99 μmol·m-2·s-1,光饱和点(LSP)为1 722.33~1 763.14 μmol·m-2·s-1,小×胡杨的光补偿点和光饱和点比小叶杨[17](光补偿点为28.97 μmol·m-2·s-1,光饱和点为1 680.60 μmol·m-2·s-1)的高,这表明小×胡杨比小叶杨对强光的适应能力更好、更喜光;小×胡杨的光补偿点比胡杨[18]的光补偿点(22.54 μmol·m-2·s-1)高,小×胡杨的光饱和点比胡杨[18](光饱和点为2 265.87 μmol·m-2·s-1)的低,说明小×胡杨对光强的利用范围还有所不足。
植物在黑暗条件(无光照)的呼吸速率称为暗呼吸速率[19]。研究显示荒漠植物能够很好的适应干旱区的环境,都具有独特的生理机制,例如暗呼吸速率(RD)一般较高[20-22]。学者们认为有的植物存在光抑制现象,随光强的增加,光合速率达到最大,光强继续增强,光合速率呈下降趋势[23];有的植物随光强的增加,光合速率达到最大,光强继续增强[24],光合速率保持平稳(一定范围内),本研究当光合有效辐射(PAR)为1 500 μmol·m-2·s-1时,曲线处于缓慢上升趋势,说明小×胡杨没有达到光饱和点。下一步需要继续增加光合有效辐射(PAR),探究小×胡杨达到光饱和点后的光响应特性。
光合有效辐射(PAR)0~1 500 μmol·m-2·s-1,1年、2年、3年生小×胡杨胞间CO2浓度随着光合有效辐射(PAR)的增加变化规律一致,均呈下降趋势。这是由于仪器叶室内的CO2浓度不变,胞间CO2浓度(Ci)越低,相应的具有越大的CO2浓度梯度值,CO2进入叶片内更为有利,更好地提供源物质,从而增强光合作用,提高光合能力的潜力。净光合速率(Pn)与气孔导度(Gs)的光响应显示,1年、2年、3年生小×胡杨气孔导度(Gs)变化趋势一致,均呈增加趋势。同时期的净光合速率(Pn)变化较为迅速,说明在光和有效辐射(PAR)0~1 500 μmol·m-2·s-1下,CO2进入小×胡杨叶片后转化速率快,提高了小×胡杨光合速率。
1年、2年、3年生小×胡杨气孔导度及蒸腾速率变化趋势相似。随着气孔导度增加,1年、2年、3年生小×胡杨的蒸腾速率逐渐增加。小×胡杨蒸腾速率(Tr)与气孔导度(Gs)呈正相关。这与赵辉等[5]的研究结果一致。蒸腾速率(Tr)随着光强的增加而迅速增加,光强逐渐增强,气孔导度(Gs)逐渐增加,CO2进入叶片的阻力减小,植物叶片内CO2浓度增加,植物体内水分会大量散失(通过气孔),导致蒸腾速率(Tr)上升较快。气孔能够自动调节开闭程度,使植物能够更加适应环境变化。
乌兰布和沙漠环境干旱少雨,光照强烈,荒漠植物生长发育的主要限制因子是水分,植物的水分利用效率越大,说明抗旱能力越强[25]。因此,为了适应沙漠干旱高温特殊环境条件,植物必须具有较高的光合速率和水分利用效率[26-27],本研究不同栽植年限小×胡杨水分利用效率的光响应特性相似,随着光强的增加,水分利用效率逐渐增加,不同栽植年限小×胡杨净光合速率(Pn)随着光和有效辐射(PAR)的增加而持续增加,蒸腾速率(Tr)随着光和有效辐射(PAR)的增加而缓慢增加,表明小×胡杨净光合速率随光强的增加而增大,从而对水分的利用效率也越高。
研究显示,影响植物对环境的适应性的因子非常多(植物根系、光合生理),植物自身特征也是关键因素[28]。因此,植物的适应性需要综合考虑所有的影响因子。本研究发现1年、2年、3年生小×胡杨具有高光合速率、高水分利用效率及相对较低的蒸腾速率,对乌兰布和沙漠东北缘环境的适应性较好。本研究只对1年、2年、3年生小×胡杨光合生理参数进行了探讨,今后还需要对小×胡杨进行更加详细的研究,才能对小×胡杨的适应性进行综合评价。
3.2 结论
在光合有效辐射(PAR)0~1 500 μmol·m-2·s-1范围内,不同栽植年限小×胡杨净光合速率均随着光合有效辐射(PAR)的增强而增加,当光合有效辐射(PAR)为1 500 μmol·m-2·s-1时,曲线处于缓慢上升趋势,小×胡杨还没有达到光饱和点。在光合有效辐射(PAR)0~1 500 μmol·m-2·s-1范围内,不同栽植年限小×胡杨气孔导度(Gs)随光合有效辐射(PAR)的增加而增加,蒸腾速率随着有效光辐射的增强而逐渐增加,变化趋势都较缓慢平稳,蒸腾速率对光合有效辐射(PAR)的变化并不敏感,而胞间CO2浓度(Ci)均呈逐步下降的趋势。当光合有效辐射<600 μmol·m-2·s-1,1年、2年、3年生小×胡杨水分利用效率增加迅速,当光合有效辐射>600 μmol·m-2·s-1,随着光强的增强,3年小×胡杨水分利用效率继续缓慢上升,但1年、2年生小×胡杨水分利用效率趋于平缓甚至下降。
1年、2年、3年生小×胡杨具有高光合速率、高水分利用效率及相对较低的蒸腾速率,对乌兰布和沙漠东北缘环境的适应性较好。需要指出的是,今后还应该对小×胡杨生理生态特性进行深入细致的研究,才能综合评价小×胡杨的适应性。
猜你喜欢
农业工程学报(2022年3期)2022-04-16
西南农业学报(2021年7期)2021-08-23
节水灌溉(2021年5期)2021-05-26
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
江苏农业科学(2020年12期)2020-07-20
中国农业科技导报(2020年12期)2020-03-15
西安科技大学学报(社会科学版)(2019年1期)2019-09-10
新课程·中旬(2014年10期)2014-12-26
