
枸杞果胶酯酶基因LbPME克隆及表达分析
2021-09-28 09:11李浩霞尹跃杨斌安巍赵建华
江苏农业科学 2021年18期
李浩霞 尹跃 杨斌 安巍 赵建华

摘要:以宁夏枸杞(Lycium barbarum L.)'宁杞1号'果实为材料,利用RT-PCR技术,克隆出LbPME的全长序列为1 152 bp,包含完整的开放阅读框(ORF),编码有384个氨基酸,蛋白质分子量为42.63 ku,理论等电点9.20;LbPME编码的蛋白包含2个可能的N-糖基化结合位点(Asn46和Asn183),4个底物结合位点(Thr166、Gln201、Arg301和Trp303)和2个酶活性位点(Asp224和Asp245)。分析发现,该蛋白由多个呈右手螺旋的β折叠肽链组成,且含有1个中空三角柱状的PME经典结构;LbPME与茄科的烟草的同源性达到90.86%。利用实时荧光定量技术,发现不同组织中LbPME均有表达,且在茎中最高而叶中最低,两者存在显著差异;随着果实的发育,LbPME表达量呈现出先升后降趋势,在青果期表达量最高,随后显著降低,在成熟期维持较低水平。本研究结果为进一步探讨枸杞LbPME的功能奠定了基础。
关键词:枸杞;果胶酯酶基因;克隆;表达
中图分类号: S188 文献标志码: A
文章编号:1002-1302(2021)18-0033-06
收稿日期:2021-01-05
基金项目:国家自然科学基金(编号:32060359);宁夏自治区科技创新领军人才项目(编号:KJT2017004);宁夏自治区农业科技自主创新专项(编号:NKYJ-18-16)。
作者简介:李浩霞(1977―),女,甘肃景泰人,副研究员,主要从事旱作农业及栽培生理研究。E-mail:lihaoxia0943@163.com。
通信作者:赵建华,博士,副研究员,主要从事枸杞种质遗传改良与分子生物学研究。E-mail:zhaojianhua0943@163.com。
果胶酯酶又称果胶甲酯酶(pectin methylesterase,简称PME),是细胞壁代谢的关键酶之一,PME催化果胶分子去甲酯化后易与钙离子结合,使细胞壁硬化,延缓细胞的生长;同时,PME在去甲酯化过程中,释放大量氢离子,形成低pH值的胞外环境,聚半乳糖醛酸酶和果胶裂解酶活性增强,造成果胶大量降解,促进细胞加快生长[1-2]。在柑橘、香蕉、苹果、葡萄、胡萝卜、番茄、马铃薯、豌豆等植物中均有关于PME的报道[3]。拟南芥PME基因仅在花粉中特异表达,缺失该基因会造成花粉管形态异常及种子数减少[4],而拟南芥PME基因的表达受丁香假单胞杆菌番茄致病变种的诱导,当植株受到病菌侵染时,该基因表达量大幅上升[5]。豌豆根尖PME基因的表达与根边缘细胞的游离紧密相关,该基因表达会抑制根边缘细胞的正常游离[6]。休眠状态的黄杉种子未检测到PME的活性,但逐渐增加外源PME浓度,会引起种子休眠的破坏和萌发[7]。随着樱桃的果实果肉的软化,PME活性逐渐增高[8]。此外,通过QTL分析筛选出3个果胶酯酶参与番茄果实抗坏血酸含量的调控[9]。在转CbPMEI1和PMEI13基因拟南芥中,PME参与低温和盐胁迫下根系生长调控[10]。因此,PME在植物生长发育、种子萌发和果实软化等过程中发挥着重要作用[11]。
PME广泛存在于高等植物中,目前已从柑橘[12]、棉花[13]、草莓[14]、番茄[15]、枇杷[16]等中分离出PME基因。拟南芥PME基因在花和荚果整个发育阶段均有不同程度的表达[17]。研究发现,PME基因大多以基因家族的形式存在[18-19],水稻有59个编码基因,拟南芥有66个编码基因,白杨有89个编码基因[20-22],但目前有关于枸杞PME的研究国内还尚未见到报道,本研究以宁夏枸杞'宁杞1号果实为材料,基于枸杞果实转录组测序数据,克隆出枸杞果实中LbPME的 cDNA 全长,并分析枸杞果实发育各个时期LbPME的表达特征,为进一步研究枸杞LbPME基因的功能及枸杞生长代谢机制提供理论依据。
1 材料与方法
1.1 材料
供试材料为宁夏枸杞'宁杞1号'品种,均采自宁夏银川市西夏区芦花台园林场国家枸杞种质资源圃(106°9′ E、38°380′ N,海拔1 100 m),选取6年生'宁杞1号'树3棵,于2018年6―7月,在果实发育的幼果期(开花后约9 d)、青果期(开花后约15 d)、色变期(开花后约22 d)、初熟期(开花后约29 d)和成熟期(开花后约36 d),分别选取果粒大小均匀、果色一致的无病虫害的果实;在果实成熟期采集枸杞根、茎、叶和果实,所用材料用液氮迅速冻结后,运送回国家枸杞工程中心分子生物学实验室后立即放置于-80 ℃冰箱保存备用。
1.2 方法
1.2.1 RNA提取及反转录 采用Trizol 试剂盒(北京TIANGEN公司)提取枸杞果实的总RNA,利用RT-PCR试剂盒(美国ThermoFisher公司)对提取的总RNA进行反转录,获得单链cDNA,用于后续LbPME的克隆及表达分析。
1.2.2 目的基因克隆 依据转录组所获得的EST序列,设计全长引物:LbPME-F和LbPME-R(表1)。以枸杞果实反转录cDNA为模板,扩增目的基因LbPME的全长序列。PCR反应体系:2×PCR Buffer 25 μL,dNTP (2 mmol/L) 10 μL,LbPME-F (10 μmol/L) 2 μL,LbPME-R (10 μmol/L) 2 μL,cDNA 模板1 μL,KOD FX Neo(1 U/μL,Toyobo Life Science Cat)1 μL,Millipore H2O补足50 μL。PCR反应程序:98 ℃预变性5 min;98 ℃变性 10 s,60 ℃退火30 s,68 ℃延伸2 min,共30个循环;68 ℃延伸5 min。PCR擴增产物克隆到pMD18-T载体(德国TakaRa公司)上进行测序。
1.2.3 实时荧光定量表达分析 根据LbPME序列设计荧光定量引物LbPME-qRTF和LbPME-qRTR(表1),以18S基因为内参基因,在BIO-RAD CFX ConnectTM 荧光定量 PCR 仪中进行扩增。反应体系:Power SYBR Green PCR Master Mix (Applied Biosystems Cat)12.5 μL,LbPME-qRTF(10 μmol/L)1 μL,LbPME-qRTR(10 μmol/L) 1 μL, cDNA模板5 μL, Millipore H2O补足25 μL。内参基因18S使用的实时荧光定量PCR引物为 18S-F2 和18S-R2。反应条件:95.0 ℃预变性 3 min;循环为95.0 ℃变性10 s,55 ℃退火30 s,72 ℃ 延伸20 s,75 ℃读板5 s,共40个循环;溶解曲线从65 ℃上升到95 ℃, 每次读板上升0.5 ℃。通过计算2-ΔΔCT值來确定该基因的相对表达量。
1.2.4 生物信息学分析 利用DNAman软件对LbPME全长进行相应氨基酸序列分析;利用ProtParam tool(http://web.expasy.org/protparam/)完成蛋白基本性质分析;采用PSORT Prediction(http://psort1.hgc.jp/form.html)完成亚细胞定位分析;使用NCBI Conserved Domains(https://www.ncbi.nlm.nih.gov/Structure/cdd/docs/cdd_search.html)对序列的结构域和功能进行分析;使用SWISS-MODEL(https://swissmodel.expasy.org)进行蛋白质三级结构预测分析;在NCBI网站运行的Blast程序数据库中对LbPME氨基酸序列进行相似性搜索,并进行了聚类分析;利用MEGA7软件中提供的比对功能进行多序列比对并基于邻接法(Neighbor-joining method,NJ)构建系统进化树bootstrap系数为1 000次,进化距离计算采用泊松校正方法。
2 结果与分析
2.1 枸杞LbPME基因克隆分析
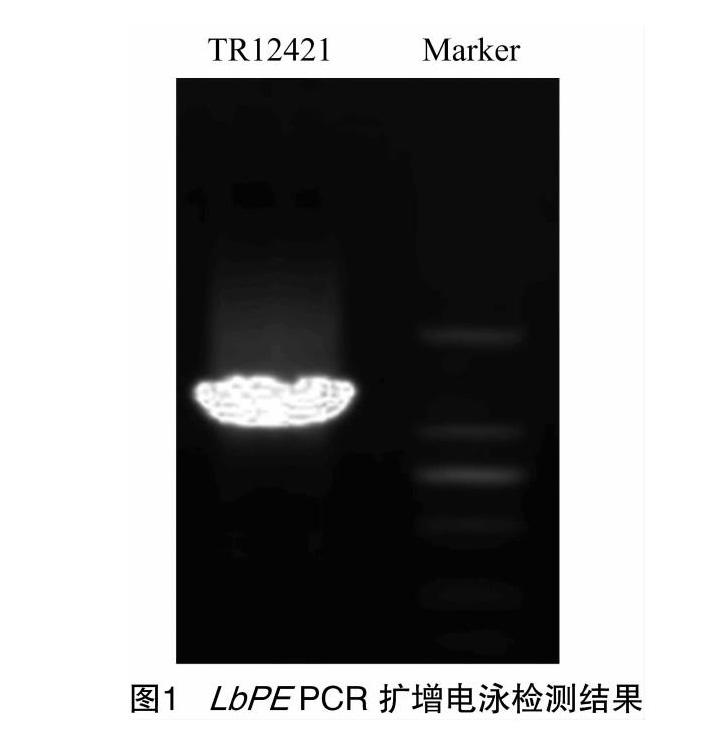
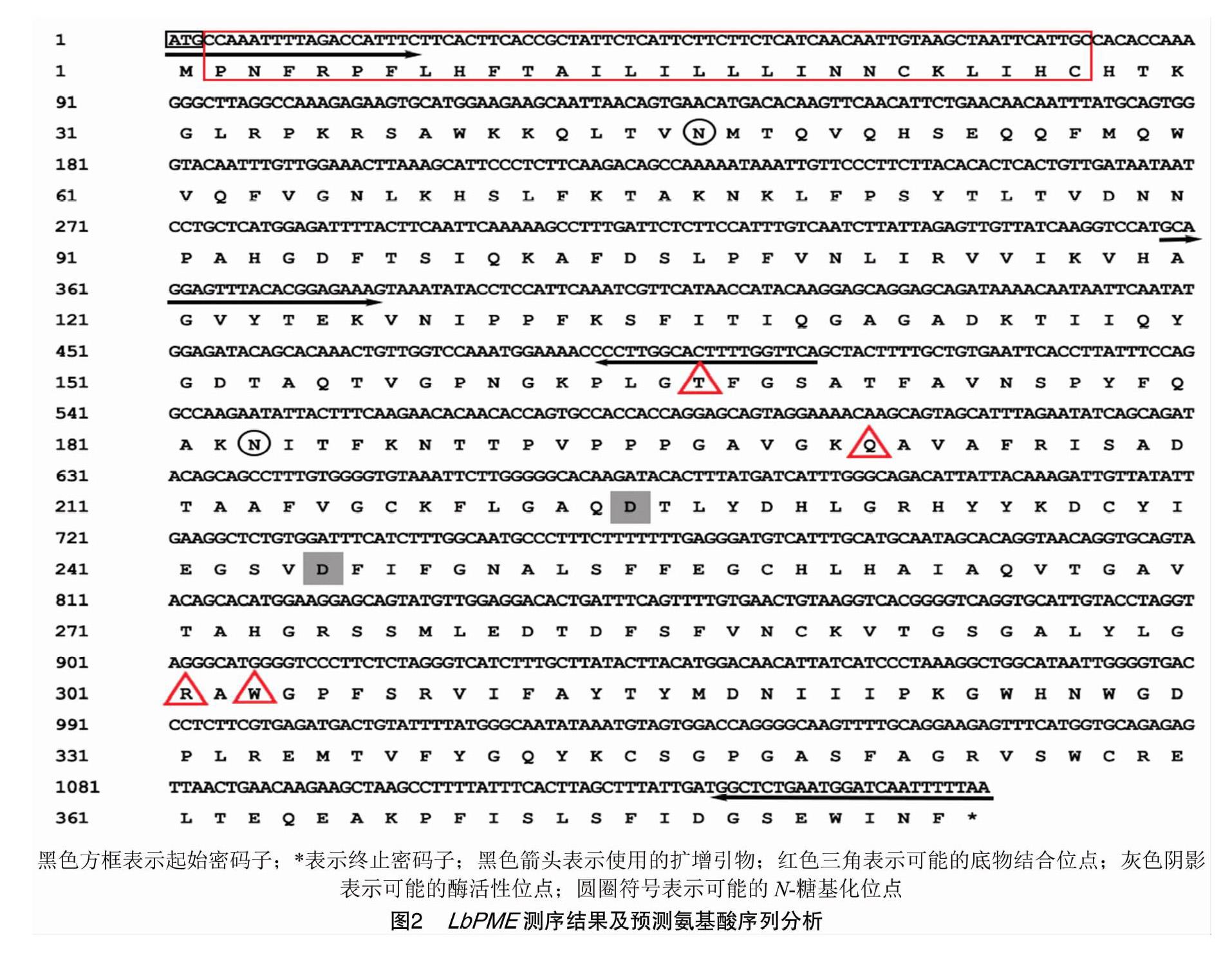
基于转录组测序数据分析,发现TR12421|c0_g1的EST基因在果实发育过程中FRKM值呈现出显著差异。以该EST序列设计特异性异物,从枸杞果实中获得枸杞果胶酯酶基因片段(图1)。该片段测序长度为1 152 bp(图2)。经Blast比对发现,该序列与果胶酯酶同源,暂时命名为LbPME,NCBI注册号为KX584484。
2.2 枸杞LbPME基因氨基酸序列及进化树分析
测序结果如图2所示。该基因含有1个1 152 bp 的开放阅读框,编码有384个氨基酸,其理论等电点和分子量分别为9.20和42.63 ku,该基因的氨基酸序列分子式为C1 957H2 954N512O533S14,包含25个碱性氨基酸(Asp+Glu)和36个酸性氨基酸(Arg+Lys)。消光系数为58 370,吸光系数为1.369。脂肪酸系数为77.68,不稳定系数为28.40,是一个较稳定的蛋白。
对其结构域进行分析,结果表明,该蛋白在Ser37~Ile381位氨基酸序列与果胶酯酶结构域(PLN02682)有较高相似性,其E值为0e+00。由图3可见,该蛋白具有果胶酯酶的显著特征,在LbPME蛋白序列中发现有2个可能的N-糖基化结合位点(Asn46和Asn183),4个底物结合位点,分别是Thr166、Gln201、Arg301和Trp303,2个酶活性位点Asp224和Asp245,以及1个疑似的过渡态稳
定位点Gln223(未在图上标示)。对其进行三级结构预测,LbPME由多个β折叠呈现右手螺旋排列,形成中空的三角形柱状果胶酯酶经典结构,其中一侧为底物结合与催化位点。无规卷曲形成了4个底物结合位点(Thr166、Gln201、Arg301和Trp303),这4个位点形成一个四边形结构。
采用PSORT Prediction(http://psort1.hgc.jp/form.html)对LbPME蛋白进行亚细胞定位,结果表明,该酶有可能是一种分泌在细胞外蛋白,并结合到细胞壁上发挥其功能。利用Blast比对分析,LbPME与茄科的同源性最高,与烟草的同源性达到90.86%。利用MEGA7软件,构建果胶酯酶氨基酸序列的系统发育树(图4),枸杞LbPME与茄目物种烟草、辣椒、马铃薯、牵牛花聚类到一个分枝,并与真子叶菊分支的其他2个目(唇形目和菊目)果胶酯酶聚类到一个大分枝上;但与真子叶蔷薇分支物种的果胶酯酶完全分离。此外,蔷薇分支物种樱桃与桑树完全聚类到不同的2个分枝中;但亲缘关系较远的金虎尾目胡杨与无患子目柑橘却聚到一起。这也说明果胶酯酶在植物进化过程中既具有很好的保守性,又在物种间呈现出少量变异。
2.3 枸杞LbPME基因表达特征分析
通过qRT-PCR 分析发现,枸杞植株根、茎、叶、果实等组织器官中的LbPME均有表达,其中,茎中表达量最高,果实中次之,叶中最低,且茎中表达量显著高于其他3个组织器官(图5-A)。随着枸杞果实发育,果实中LbPME呈现出先升后降的变化趋势(图5-B),在青果期表达量达到最大, 随后显著降低,在果实成熟时表达量维持较低水平。在果实发育的幼果期和成熟期枸杞LbPME表达量较低,且2个发育期表达量无显著差异,但显著低于其他3个时期,可见,枸杞LbPME在果实发育青果期到初熟期表现得更为活跃,但在果实成熟时可能由于细胞壁发育趋于稳定,LbPME表达量降低。
3 结论与讨论
自Balestrierii等从猕猴桃克隆出第1个PME基因后[23],对植物PME基因的研究愈加深入。 植物天然PME分子量一般在15~36 ku[9],如胡萝卜34.5 ku[24],大豆33 ku[25],脐橙36.2 ku[26]。在本研究中,成功克隆出枸杞LbPME全长为1 152 bp,包含1个完整的开放阅读框(ORF),编码383个氨基酸,其理论等电点和分子量分别为9.20和42.63 ku。而在枇杷中分离出2个EjPPME1和EjPPME2基因,均包含1个1 737 bp的开放阅读框(ORF),共编码578个氨基酸,EjPPME1理论等电点为8.94,理论分子量为63.05 ku,EjPPME2理论等电点为9.04,理论分子量为63.12 ku[16]。可见,不同植物PME蛋白分子存在一定差异,这可能是PME基因大多以基因家族的形式存在的缘故[18-19]。进一步对枸杞LbPME蛋白的三级结构域分析发现,该蛋白具有PME的显著特征,由多个呈右手螺旋的β折叠的肽链排列而成,并且形成中空三角柱状的PME经典结构,这与胡萝卜、番茄的PME三维结构基本相似[27-28]。
PME是细胞壁代谢的关键酶之一,主要作用是催化植物细胞壁中的果胶去甲酯化,生成果胶酸和甲醇,进而调控植物的生长发育。PME广泛存在于高等植物中,其表达存在时空差异特性,一些PME基因在整个植株的生命周期能持续表达[29]。但也有一些PME基因只在植株生长发育的特定阶段或特定的组织器官中表达。在草莓上发现,FaPE1仅在果实中表达,FaPE2仅在叶中表达,FaPE3-4在整个植株中均有表达[15]。在本研究中,LbPME在整个植株均有表达,其中,在茎中的表达量最高,在叶中的表达量最低。随着果实不断发育,果实中LbPME表达量呈现出先升后降变化趋势,在整个发育阶段青果期表达量最高,幼果期最低,这与草莓果实发育过程中PME基因变化趋势基本一致。这就表明LbPME在枸杞果实发育的青果期发挥重要作用,但随着果实不断发育,LbPME的表達量逐渐降低,在果实成熟时维持着较低表达水平,其参与枸杞果实细胞壁成熟表达水平明显减弱,但LbPME在果实发育过程中参与果实细胞壁合成调控还有待于进一步研究。
参考文献:
[1]于海伟,陈宝刚.白桦果胶甲酯酶抑制剂(PMEI)基因克隆及序列分析[J]. 防护林科技,2012(1):64-67.
[2]龙海蓉,王齐玮,李晓平,等. 生长期和植株性别对工业大麻杆果胶含量及质量的影响[J]. 西北林学院学报,2016,31(2):244-248.
[3]叶 华,马 力,高秀容.果胶酯酶的研究进展[J]. 生物学杂志,2005,22(6):5-7.
[4]Jiang L X,Yang S L,Xie L F,et al. Vanguard1 encodes a pectin methylesterase that enhances pollen tube growth in the Arabidopsis style and transmitting tract[J]. Plant Cell,2005,17(2):584-596.
[5]张 倩,鲍依群,谭小云. 拟南芥果胶甲酯化酶基因PME17在抵御丁香假单胞杆菌番茄致病变种DC3000株中的功能[J]. 植物生理学报,2015,51(7):1061-1066.
[6]Wen F S,Zhu Y M,Hawes M C. Effect of pectin methylesterase gene expression on pea root development[J]. Plant Cell,1999,11(6):1129-1140.
[7]Ren C,Kermode A R. An increase in pectin methyl esterase activity accompanies dormancy breakage and germination of yellow cedar seeds[J]. Plant Physiology,2000,124(1):231-242.
[8]Barrelt D M,Gonzalez C. Activity of softening enzymes during cherry maturation[J]. Journal of Food Science,1994,59(3):574-577.
[9]Ruggieri V,Sacco A,Calafiore R,et al. Dissecting a QTL into candidate genes highlighted the key role of pectinesterases in regulating the ascorbic acid content in tomato fruit[J]. Plant Genome,2015,8(2):1-10.
[10]Chen J,Chen X,Zhang Q,et al. A cold-induced pectin methyl-esterase inhibitor gene contributes negatively to freezing tolerance but positively to salt tolerance in Arabidopsis[J]. Journal of Plant Physiology,2018,222(8):67-78.
[11]李 欢,张舒怡,张 钟,等. 鲜食枣与制干枣的成熟软化机理差异研究[J]. 西北林学院学报,2017,32(5):137-143.
[12]叶 华. 柑橘果胶酯酶的基因克隆[D]. 成都:西华大学,2006.
[13]陈婷婷,李旭凯,王如意,等. 棉花GhPME1和GhPME2基因的克隆及表达分析[J]. 中国农业大学学报,2012,17(5):7-14.
[14]Castillejo C,De La Fuente J I,Iannetta P,et al. Pectin esterase gene family in strawberry fruit:study of FaPE1,a ripening-specific isoform[J]. Journal of Experimental Botany,2004,55(398):909-918.
[15]葛英亮. 樱桃蕃茄中果胶酯酶的基因克隆及在毕赤酵母中表达的初步研究[D]. 成都:西华大学,2007.
[16]唐佳佳. 枇杷果胶酯酶基因克隆及表达特性分析[D]. 重庆:西南大学,2014.
[17]张 莉,薛金爱,于浩泉,等. 调控拟南芥花瓣脱落的果胶甲酯酶基因表达和功能分析[J]. 植物生理学报,2012,48(4):350-358.
[18]Markovic O,Janecek S. Pectin methylesterases:sequence-structural features and phylogenetic relationships[J]. Carbohydrate Research,2004,339(13):2281-2295.
[19]Yokoyama R,Nishitani K. Genomic basis for cell-wall diversity in plants. A comparative approach to gene families in rice and Arabidopsis[J]. Plant & Cell Physiology,2004,45(9):1111-1121.
[20]Pelloux J,Rustérucci C,Mellerowicz E J. New insights into pectin methylesterase structure and function[J]. Trends in Plant Science,2007,12(6):267-277.
[21]Louvet R,Cavel E,Gutierrez L,et al. Comprehensive expression profiling of the pectin methylesterase gene family during silique development in Arabidopsis thaliana[J]. Planta,2006,224(4):782-791.
[22]Geisler-Lee J,Geisler M,Coutinho P M,et al. Poplar carbohydrate-active enzymes. Gene identification and expression analyses[J]. Plant Physiology,2006,140(3):946-962.
[23]Balestrierii C,Castaldo D,Giovane A,et al. A glycoprotein inhibitor of pectin methylesterase in kiwi fruit (Actinidia chinensis)[J]. European Journal of Biochemistry,1990,193(1):183-187.
[24]Ly-Nguyen B,Van Loey A M,Fachin D,et al. Partial purification,characterization,and thermal and high-pressure inactivation of pectin methylesterase from carrots (Daucus carrota L.)[J]. Journal of Agricultural and Food Chemistry,2002,50(19):5437-5444.
[25]Ricard J,Noat G. Electrostatic effects and the dynamics of enzyme reactions at the surface of plant cells[J]. European Journal of Biochemistry,1986,155(1):191-197.
[26]Versteeg C,Rombouts F M,Spaansen C H,et al. Thermostability and orange juice cloud destabilizing properties of multiple pectinesterases from orange[J]. Journal of Food Science,1980,45(4):969-971.
[27]Johansson K,El-Ahmad M,Friemann R,et al. Crystal structure of plant pectin methylesterase[J]. FEBS Letters,2002,514(2/3):243-249.
[28]DAvino R,Camardella L,Christensen T M,et al. Tomato pectin methylesterase:modeling,fluorescence,and inhibitor interaction studies-comparison with the bacterial (Erwinia chrysanthemi) enzyme[J]. Proteins,2003,53(4):830-839.
[29]Caffe J,Tiznado M E,Handa A K. Characterization and functional expression of a ubiquitously expressed tomato pectin methylesterase[J]. Plant Physiology,1997,114(4):1547-1556.
猜你喜欢
环球时报(2022-09-20)2022-09-20
学与玩(2022年2期)2022-05-03
今日农业(2020年24期)2020-12-15
阅读(低年级)(2019年9期)2019-11-15
基层中医药(2018年10期)2018-12-06
校园英语·上旬(2016年10期)2016-11-16
人间(2016年28期)2016-11-10
兽医导刊(2016年12期)2016-05-17
现代检验医学杂志(2015年4期)2015-02-06
