
桃蛀螟卵巢发育过程及其与温度的关系
2021-10-12 13:15张胜男李国平田彩红黄建荣封洪强
植物保护 2021年5期
张胜男 李国平 田彩红 黄建荣 封洪强


摘要 本文明確了桃蛀螟的雌性生殖系统由1对卵巢、1对侧输卵管、1根中输卵管、1个交配囊和1个受精囊组成。依据雌性生殖系统发育的典型特征,即卵巢小管形态、卵粒的成熟度、脂肪体的多少及形态,将卵巢发育过程分为乳白透明期(Ⅰ级)、卵黄沉积期(Ⅱ级)、成熟待产期(Ⅲ级)、产卵盛期(Ⅳ级)及产卵末期(Ⅴ级)5个发育时期。在温度为20、25℃和30℃下,桃蛀螟初羽化成虫完成卵巢发育的全部过程(Ⅰ级~Ⅴ级)所需时间分别为11.43、938、8.62 d。经推算,桃蛀螟卵巢各级(Ⅰ级~Ⅴ级)的发育起点温度分别为22.88、21.1、16.55、18.44、1119℃,有效积温分别为0.37、4.45、23.22、35.10日·度和125.91 日·度。根据不同温度下卵巢发育的速度,建立了卵巢发育速率的预测模型,这为桃蛀螟的短期精准预报提供了科学依据。
关键词 桃蛀螟; 卵巢解剖; 分级标准; 卵巢发育速率预测模型
中图分类号: S 435.132
文献标识码: A
DOI: 10.16688/j.zwbh.2020370
Ovarian development of Conogethes punctiferalis (Guenée) and its relationship with temperature
ZHANG Shengnan, LI Guoping, TIAN Caihong, HUANG Jianrong*, FENG Hongqiang*
(Henan Key Laboratory of Crop Pest Control, Key Laboratory of Integrated Pest Management on Crops in
Southern Part of Northern China, Ministry of Agriculture and Rural Affairs; International Joint Research
Laboratory for Crop Protection of Henan; Institute of Plant Protection, Henan Academy of Agricultural
Sciences, Postgraduate T&R Base of Zhengzhou University, Zhengzhou 450002, China)
Abstract
This article clarified the female reproductive system of Conogethes punctiferalis, which was composed of a pair of ovaries, a pair of lateral oviducts, one middle oviduct, one copulatory pouch and one bursa copulatrix. According to the typical characteristics of the developing female reproductive system, including the fine morphology of the ovarian tubules, the maturity of eggs, the number and morphology of the fat body, the ovarian developmental process was divided into five developmental stages: the transparent stage (level Ⅰ), the vitellogenesis stage (level Ⅱ), the egg maturation stage (level Ⅲ), the egg-laying stage (level Ⅳ) and the late egg-laying stage (level Ⅴ). At the temperatures of 20, 25℃ and 30℃, the developmental durations of level Ⅰ to level Ⅴ were 11.43, 9.38, 8.62 d, respectively. After calculation, the developmental temperature thresholds and sum of degree-day accumulation required for level Ⅰ- level Ⅴ of ovarian development were 22.88, 21.11, 1655, 1844, 11.19℃, and 0.37, 4.45, 23.22, 35.10, 125.91 degree-days, respectively. According to the speed of ovarian development at different temperatures, an ovarian development prediction model was established, which provides a scientific basis for the accurate short-term forecast of C.punctiferalis.
Key words
Conogethes punctiferalis; ovary anatomy; grading standards; ovary development model
害虫测报是进行害虫精准防控的基础,而灯诱、性诱、卵巢解剖及田间调查等技术是进行害虫测报的有效措施[1]。卵巢解剖不仅能确定害虫种群的卵巢发育级别,判断产卵高峰期,推断害虫的防治时期,还可以确定种群的繁殖能力,明确害虫的迁飞潜力,是害虫短期预测的关键技术[2-3]。
在1957年初农业部制定的黏虫 Mythimna separata 测报办法中把解剖雌蛾、观察其卵巢发育进度作为测报的常规方法,这是卵巢解剖工作应用于我国害虫测报工作的开端[4]。通过近几十年来不断研究,已确定了黏虫、棉铃虫 Helicoverpa armigera、亚洲玉米螟 Ostrinia furnacalis、二点委夜蛾 Athetis lepigone、褐飞虱 Nilaparvata lugens 和稻纵卷叶螟 Cnaphalocrocis medinalis 等多种农业害虫的卵巢分级标准[5-10],构建了卵巢解剖应用技术的短期预测预报系统,已成为我国害虫测报的常规技术手段。
桃蛀螟 Conogethes punctiferalis (Guenée) 寄主植物有100余种,既为害桃、苹果、石榴、板栗等果树,也为害玉米、向日葵、高粱、蓖麻等农作物,是一种分布广泛的多食性农业害虫[11]。由于农业产业结构调整及气候变化等原因,致使该虫在国内很多地区的为害逐年加重,已成为多种果树和农作物上的一种重要害虫[12]。为准确预测桃蛀螟的发生趋势,本文将详细描述不同温度下桃蛀螟卵巢发育的形态变化过程,确定桃蛀螟卵巢发育的分级标准,构建基于温度的卵巢发育预测模型,研究结果对准确预测桃蛀螟种群动态具有重要意义。
1 材料与方法
1.1 供试虫源和饲养条件
幼虫于2019年9月在河南省农业科学院现代农业研究开发基地监测点(35.02°N,113.69°E)的玉米上采集,带回实验室后将幼虫放置在温度为(26±1)℃、湿度为(75±5)%、光周期为 L∥D=16∥8 h的培养箱内,每天使用玉米穗饲养至第二代初羽化成虫备用。
1.2 成虫的饲养和卵巢解剖
将每250头初羽化的成虫(雌雄比为1∶1.5)置于30 cm×30 cm×30 cm的养虫笼中,并为其提供10%的白糖水,将养虫笼分别置于20、25℃和30℃的培养箱内饲养,每日从3个养虫笼中各随机选取10头雌蛾对其卵巢进行解剖,同时记录不同温度和不同日龄雌蛾的卵巢发育特征,试验设置3次重复。解剖时先将桃蛀螟雌蛾用乙醚麻醉,去除翅和足,转入培养皿,再用解剖镊子从胸腹连接处向尾部逐步撕开体壁,使卵巢完整地暴露在外面,去除多余脂肪体,浸泡至蒸馏水中,最后使用超景深三维显微镜系统(VHX200,KEYENCE)进行显微观察和拍照。卵巢的观察参照苏德明等[4]的方法,确定桃蛀螟雌蛾生殖系统的结构组成,观察这些结构的形态变化特征,依据典型的特征确定卵巢的分级标准。
1.3 卵巢发育速率的预测模型
先将不同温度下桃蛀螟成虫卵巢发育到Ⅰ至Ⅴ级的历期进行加权平均,求出发育到各级别的平均发育历期D,将其换算成相应温度下的平均发育速率V(V=1/D),然后根据有效积温法则,采用最小二乘法计算桃蛀螟成虫卵巢发育各级别的发育起点温度C和有效积温K[13]。公式如下:
C=∑V2∑T-∑V∑VTn∑V2-(∑V)2
K=n∑VT-∑V∑Tn∑V2-(∑V)2
式中,T为环境温度;C为发育起点温度;K为有效积温常数;V为发育速率(1/D);n为试验温度组数。
卵巢发育至Ⅰ~Ⅴ级历期经对数转换后采用Tukey多重比较法比较其在不同温度下发育历期的差异。使用一元线性回归法建立不同温度下成虫卵巢各级别的发育速率的预测模型(V=a+bT,a、b为模型参数,V为各发育阶段的发育速率,T为温度),通过F检验确定线性模型的拟合效果,所有分析均使用R 3.5.3软件完成。
2 结果与分析
2. 1 桃蛀螟卵巢的基本构造与分级
桃蛀螟雌蛾的生殖系统主要由1对卵巢、1对侧输卵管、1根中输卵管、1个交配囊和1个受精囊组成(图1)。1对卵巢分别位于腹腔消化道的两侧,被脂肪体包围粘连在一起,每个卵巢由4根卵巢小管组成,卵巢小管开口于侧输卵管,两根侧输卵管汇集为1根中输卵管,中输卵管开口于第九腹节。在中输卵管前端着有1个短而弯曲的受精囊,在中输卵管末端处连接着长圆形的交配囊,阴道位于排卵口前端,受精囊开口于阴道顶部,受精囊根部连接一根较长的附腺(图1)。
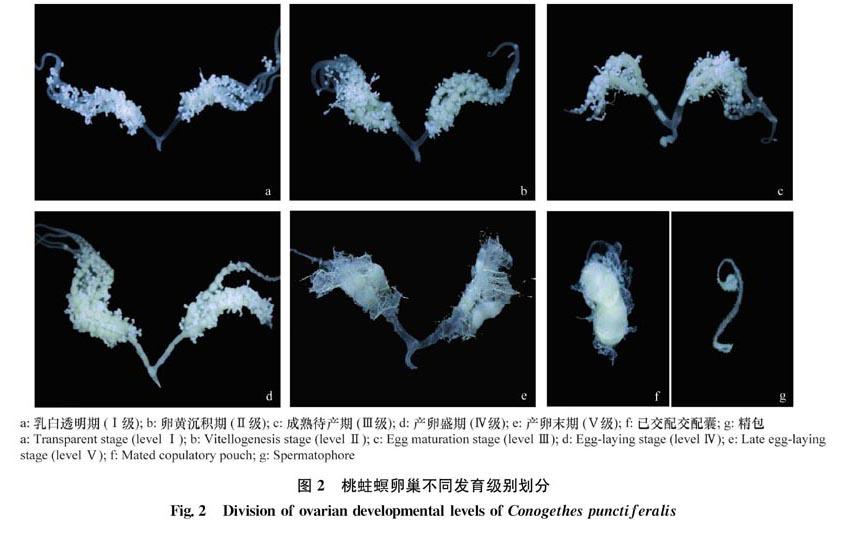
雌蛾从初羽化至死亡,其卵巢小管由光滑细小发育至粗大充满卵粒而后萎缩,附着在卵巢小管上的脂肪体由丰富至逐渐消失,侧输卵管和中输卵管由透明光滑至畸形,交配囊由乳白色球状至不规则扁球状。参考亚洲玉米螟、稻纵卷叶螟等螟蛾科昆虫按腹腔颜色、器官特征、脂肪体形态等各种器官发育特征把卵巢发育过程划分为5个级别(Ⅰ~Ⅴ级),本文也把桃蛀螟卵巢发育过程划分为5个级别。以卵巢小管细小透明作为Ⅰ级卵巢的典型特征;卵巢小管内有成熟卵粒作为Ⅱ级卵巢的典型特征;侧输卵管充满卵粒作为Ⅲ级卵巢的典型特征;整个卵巢充满卵粒作为Ⅳ级卵巢的典型特征;卵巢周围几乎没有脂肪体和卵粒作为Ⅴ级卵巢的典型特征(表1、图2)。
2.2 不同温度下桃蛀螟卵巢的发育历期
根据桃蛀螟卵巢的分级标准,判断初羽化成虫在不同温度下卵巢的发育进度。结果表明,20℃下桃蛀螟雌蛾卵巢完成Ⅰ至Ⅴ级的平均天数分别为(0.49±0.087)、(1.95±0.18)、(4.06±0.24)、(6.62±0.23)d和(11.43±0.24)d;25℃下桃蛀螟雌蛾完成Ⅰ至Ⅴ级的平均天数分别为(0.34±0.045)、(1.50±0.15)、(2.93±2.72)、(579±023)d和(9.38±0.21)d;30℃下桃蛀螟卵巢完成Ⅰ至Ⅴ级的平均天数分别为(030±0.05)、(1.29±016)、(2.73±0.30)、(4.86±020)d和(8.62±0.18)d。3个不同温度下卵巢达到Ⅰ级的历期差异不显著(P =0.2834),达到Ⅱ 級至Ⅴ级卵巢发育的历期差异均显著(P <0.05;P <0.001;P <0.001;P <0.001),在30℃下桃蛀螟卵巢发育最快(图3)。
2.3 桃蛀螟雌蛾卵巢各级别发育起点温度和有效积温
桃蛀螟卵巢发育至Ⅰ级~Ⅴ级的发育起点温度分别为(22.88±1.41)、(21.1±1.20)、(16.55±172)、(18.44±1.23)、(11.19±1.52)℃,有效积温分别为(0.37±0.34)、(4.45±1.42)、(23.22±500)、(35.10±6.33)日·度和(125.91±14.46)日·度。结果表明,桃蛀螟卵巢发育到各个级别的发育起点温度和所需要的有效积温不同。
2.4 卵巢发育的温度模型及预测
应用线性模型拟合桃蛀螟卵巢发育级别的发育速率(V)与温度(T)的关系,模型参数如表2所示,桃蛀螟卵巢达到各级别的发育速率与温度显著相关(P<0.05),这表明可以根据温度使用线性回归模型预测出桃蛀螟卵巢发育各个级别的发育历期。
3 讨论
研究昆虫生殖系统,确定卵巢发育进度是害虫预测预报的一种常用手段。鳞翅目昆虫卵巢按照发育特征一般分为5个级别,分别为乳白透明期(Ⅰ级)、卵黄沉积期(Ⅱ级)、成熟待产期(Ⅲ级)、产卵盛期(Ⅳ级)及产卵末期(Ⅴ级),例如:草地贪夜蛾 Spodoptera frugiperda[14]、棉铃虫[15]。欧洲玉米螟 Ostrinia nubilalis 卵巢则分为6个发育级别,分别为无卵黄阶段(Ⅰ级)、卵黄沉积阶段(Ⅱ级)、卵成熟期(Ⅲ级)、产卵前期(Ⅳ级)、产卵高峰期(Ⅴ级)、产卵末期(Ⅵ级)[16]。也有部分昆虫卵巢发育的乳白透明期在羽化前已经完成,因此将卵巢发育分为4个级别,如甜菜夜蛾 Spodoptera exigua[17]。草地螟 Loxostege sticticalis 的卵巢发育也分为4级,分别是初羽化期、产卵前期、产卵期和产卵末期[18]。本研究虽然没有研究蛹期的卵巢形态及发育,但研究发现初羽化桃蛀螟雌蛾已经具有成熟的卵巢小管,但无成熟的卵粒,卵粒的形成、发育及产出均在羽化后完成[19]。本研究参照鳞翅目昆虫卵巢的发育级别,根据卵巢管内有无卵粒、腹腔颜色、器官特征、脂肪体形态将桃蛀螟的卵巢发育情况分为5个等级。不同温度下,桃蛀螟卵巢发育到产卵盛期的时间不同,20℃下,需要662 d;25℃下,需要5.79 d;30℃下仅需4.86 d,温度对桃蛀螟卵巢发育有显著影响。
发育起点温度和有效积温反映了昆虫最基本的生物学特性,可用于推测其在某一地区的发生时间、世代以及各发育阶段的历期[20]。本研究得到的桃蛀螟雌蛾不同卵巢级别的发育起点温度分别为(2288±1.41)、(21.1±1.20)、(16.55±1.72)、(18.44±123)、(11.19±1.52)℃,有效积温为(037±0.34)、(445±1.42)、(23.22±5.00)、(3510±6.33)日·度和(125.91±14.46)日·度,已有研究显示桃蛀螟蛹、产卵前期和整个世代的发育起点温度分别为 14.27、7.47℃和11.85℃,有效积温依次为118.42、58.33日·度和 509.06日·度[21],与本研究结果存在一定差異,这可能由于二者试验条件和统计方法不同所致。本研究得到的桃蛀螟雌蛾卵巢各发育级别的发育起点温度和有效积温不同,是否环境温度低于发育起点温度,卵巢发育停止或发育缓慢,还需要后续更为细致的研究。
研究温度变化对昆虫生长发育的影响可以构建温度与昆虫增长速率的关系模型,用于解释温度对其发育速率的影响[22]。而研究鳞翅目昆虫卵、幼虫、蛹和成虫及全世代的发育历期,可以构建不同发育阶段的预测模型,进而还可根据温度对该虫的发生时期进行准确预测[21, 23-24]。但温度与鳞翅目昆虫雌蛾卵巢发育级别速率的关系并没有详细研究,无法准确预测鳞翅目昆虫卵巢发育至各个级别的时期,本文研究了3个温度下桃蛀螟卵巢的发育进度,建立了雌蛾羽化后卵巢各个级别的发育速率的预测模型,用于不同温度下桃蛀螟卵巢各发育等级的发育速率预测,这为准确预测产卵盛期及防治时期提供科学的依据。
雷达观测到黏虫、棉铃虫、甜菜夜蛾等多种昆虫具有迁飞习性,桃蛀螟也有跨海迁飞的习性[25]。昆虫的迁飞与卵巢发育程度密切相关,一般而言,昆虫的迁飞行为多发生在卵巢未成熟时,即本地迁出虫源卵巢发育级别及交配率均较低,卵巢发育为Ⅰ级的个体比例极高,多数未交配;昆虫的迁入种群的卵巢发育级别、交配数及交配率均较高,卵巢多为高级别[3]。因此,通过对迁飞昆虫卵巢解剖及卵巢发育进度观察,可对迁飞昆虫的虫源性质进行判别。本文在室内研究了温度对桃蛀螟卵巢发育的影响,对桃蛀螟的预测预报具有重要的参考价值,后续将进一步对桃蛀螟是否具有迁飞能力,其迁飞路径和虫源性质等进行研究。
参考文献
[1] 刘万才,黄冲. 我国农作物现代病虫测报建设进展[J]. 植物保护, 2018, 44(5): 159-167.
[2] 张春辉. 卵巢解剖在害虫测报和防治上的应用[J]. 中国农学通报, 1994, 10(1): 53.
[3] 齐国君,芦芳,胡高,等. 卵巢解剖在我国迁飞昆虫研究中的应用[J]. 中国植保导刊, 2011, 31(7): 18-22.
[4] 苏德明,李汝铎,王金其. 昆虫的卵巢发育与害虫预测预报[M]. 上海: 复旦大学出版社, 1987: 31-84.
[5] 芦芳,齐国君,秦冉冉,等. 褐飞虱卵巢发育的形态变化过程及分级标准[J]. 应用昆虫学报, 2011, 48(5): 1394-1400.
[6] 李立涛,王玉强,刘磊,等. 二点委夜蛾卵巢发育分级及在预测预报中的应用[J]. 应用昆虫学报, 2012, 49(4): 1043-1047.
[7] 李学军,刘世栋,王淑贤,等. 棉铃虫雌蛾卵巢发育分级浅探[J]. 植保技术与推广, 1993, 15(5): 17.
[8] 石毓亮,张之光,李玉椿. 粘虫 Mythimna separata (Walker) 生殖系统解剖与自然交配及其在实践中的应用[J]. 山东农学院学报. 1964, 21(9): 47-58.
[9] 钱仁贵. 玉米螟卵巢解剖及应用初报[J]. 昆虫知识, 1982, 28(5): 18-20.
[10]张孝羲,陆自强,耿济国. 稻纵卷叶螟雌蛾解剖在测报上的应用[J]. 昆虫知识, 1979, 5(3): 97-99.
[11]鹿金秋,王振营,何康来,等. 桃蛀螟研究的历史、现状与展望[J]. 植物保护,2010,36(2):31-38.
[12]王振营,何康来,石洁,等. 桃蛀螟在玉米上危害加重原因与控制对策[J]. 植物保护, 2006, 32(2): 67-69.
[13]马波,江海燕. 有效积温法则及其应用[J]. 黑龙江科技信息, 2007(20): 161.
[14]赵胜园,杨现明,和伟,等. 草地贪夜蛾卵巢发育分级与繁殖潜力预测方法[J]. 植物保护, 2019, 45(6): 28-34.
[15]张韵梅,牟吉元. 棉铃虫卵巢发育的组织化学及测报分级的研究[J]. 山东农业科学, 1994, 32(3): 7-9.
[16]KUANG Xingquan, CALVIN D D, KNAPP M C, et al. Female European corn borer (Lepidoptera: Crambidae) ovarian developmental stages: their association with oviposition and use in a classification system [J]. Journal of Economic Entomology,2004, 97(3): 828-835.
[17]王宪辉,徐洪富,许永玉,等. 甜菜夜蛾雌性生殖系统结构、发育分级及在测报上的应用[J]. 植物保护学报, 2003, 30(3): 261-266.
[18]孙雅杰,陈瑞鹿,王素云,等. 草地螟雌蛾生殖系统发育的形态变化[J]. 昆虫学报, 1991, 34(2): 248-249.
[19]JING Dapeng, ZHANG Tiantao, PRABU S, et al. PBP genes regulated by the development of the ovaries, sex pheromone release, mating and oviposition behavior in Conogethes punctiferalis (Guenée) [J/OL]. Chemoecology, 2021,31:1-9. DOI:10.1007/s00049-020-00323-3.
[20]唐業忠,朱建国,况荣平. 发育起点温度的概念和应用问题[J]. 生态学杂志, 1993, 12(6): 70-72.
[21]杜艳丽,郭洪梅,孙淑玲,等. 温度对桃蛀螟生长发育和繁殖的影响[J]. 昆虫学报,2012, 55(5): 561-569.
[22]时培建,池本孝哉,戈峰. 温度与昆虫生长发育关系模型的发展与应用[J]. 应用昆虫学报, 2011, 48(5): 1149-1160.
[23]敬甫松. 大螟发育历期的温度效应研究[J]. 昆虫知识, 1985, 31(6): 247-252.
[24]何莉梅,葛世帅,陈玉超,等. 草地贪夜蛾的发育起点温度、有效积温和发育历期预测模型[J]. 植物保护, 2019, 45(5): 18-26.
[25]FENG Hongqiang, WU Kongming, CHENG Dengfa, et al. Radar observations of the autumn migration of the beet armyworm Spodoptera exigua (Lepidoptera: Noctuidae) and other moths in northern China [J]. Bulletin of Entomological Research, 2003, 93(2): 115-124.
(责任编辑:杨明丽)
