
海南引种睡莲表型多样性分析及评价
2021-12-08 01:21潘庆龙,付瑛格,谷佳,盛玉辉,李清雪,饶英,朱天龙,周扬,史佑海,赵莹,王健
热带作物学报 2021年10期
潘庆龙,付瑛格,谷佳,盛玉辉,李清雪,饶英,朱天龙,周扬,史佑海,赵莹,王健


摘 要:以海南引種的86份睡莲资源作为研究对象,对45个表型性状进行多样性、相关性、聚类、主成分等分析。结果表明:86份睡莲表型性状存在着丰富的变异。13个数量性状的多样性指数范围为0.612~2.041,最高的是花瓣宽度(2.041),最低的是萼片枚数(0.612);32个质量性状的多样性指数范围为0.259~1.648,最高的是花色(1.648),而柱头盘颜色(0.259)和中轴突颜色(0.259)两个性状的多样性指数最低。13个数量性状的变异系数范围为19.98%~122.06%,其中具有强变异的性状有两个,分别为雄蕊个数(122.06%)、花瓣总数(109.87%)。相关性分析表明,叶长与叶宽等6对性状呈极显著性正相关(P<0.01)。聚类分析将86份睡莲分为4个类群,依次为:广热带睡莲品种群、澳大利亚睡莲品种群、古热带睡莲品种群、广温带睡莲品种群。主成分分析共提取出特征值大于1.5的8个主成分,累积贡献率为60.838%。86份供试睡莲综合评分排名最高的为‘澳洲白巨睡莲,最低的为‘小白子午莲睡莲。
关键词:睡莲资源;表型性状;聚类分析;主成分分析
中图分类号:S682.3 文献标识码:A
Analysis of Phenotypic Diversity of Nymphaea L. in Hainan, China
PAN Qinglong1, FU Yingge1, GU Jia1, SHENG Yuhui2, LI Qingxue1, RAO Ying3, ZHU Tianlong4, ZHOU Yang1, SHI Youhai1, ZHAO Ying1*, WANG Jian1*
1. Key Laboratory of Genetics and Germplasm Innovation of Tropical Forest Trees and Ornamental Plants, Ministry of Education / Key Laboratory of Germplasm Resources Biology of Tropical Special Ornamental Plants of Hainan / College of Forestry, Hainan University, Haikou, Hainan 570228, China; 2. Hengxing University, Qingdao, Shandong 266100, China; 3. Guizhou University, Guiyang, Guizhou 550025, China; 4. Hainan Fodu Lianyuan Ecological Agriculture Co., Ltd., Haikou, Hainan 570228, China
Abstract: 45 phenotypic traits of 86 water lily (Nymphaea L.) resources in Hainan were surveyed, and the diversity, correlation, clustering and principal component analysis were conducted. The phenotypic traits of the 86 water lilies varied greatly, and the diversity indexes of 13 quantitative traits ranged from 0.612 to 2.041, with the highest value of petal width (2.041) and the lowest one of sepal number (0.612). The diversity indexes of 32 quality traits ranged from 0.259 to 1.648, in which the highest value was the flower color (1.648), and the lowest value was the stigma color (0.259) and the middle axon color (0.259). The variation coefficient of the 13 quantitative traits ranged from 19.98% to 122.06%, among which the number of stamens (122.06%) and the total number of petals (109.87%) were the two most varied traits. Correlation analysis showed that significant positive correlations (P<0.01) existed between the length of leaf and width of leaf, and other 5 couples of characteristics. Clustering analysis of the 86 water lilies were divided into four groups: Brachyceras, Anecphya, Lotus and Nymphaea. Eight principal components were extracted by principal component analysis with characteristic values greater than 1.5, and the cumulative contribution rate was 60.838%. Among the 86 water lilies N. gigantea ‘Albert de Lestang ranked the highest value in comprehensive score, and N. ‘Pygmaea Alba got the lowest one.
Keywords: waterlily resource; phenotypic trait; clustering analysis; principal component analysis
DOI: 10.3969/j.issn.1000-2561.2021.10.005
睡莲为睡莲科(Nymphaeaceae)睡莲属(Nymphaea L.)多年生草本花卉[1],具观赏性、食用性及多种生物活性等特点,通常为园林水景材料[2],用作食品及饮品[3],为抗菌[4]、抗焦虑[5]、抗肝毒[6]等抗性药物[7-10]的重要活性成分,在多领域具有广泛的应用。世界上包含1000余个不同品种,分布广泛,但主要集中于热带、亚热带及温带地区[2]。睡莲属分为广热带睡莲亚属(Brachyceras)、古热带睡莲亚属(Lotos)、澳大利亚睡莲亚属(Anecphya)、新热带睡莲亚属(Hydrocallis)、广温带睡莲亚属(Nymphaea)5个亚属[11]。
种质资源是品种培育的基础和关键[12],表型性状是最直观和基础的植物种质资源遗传多样性研究方法,是植物优良品种选育的基础[13],因此研究种质资源的表型遗传多样性,对促进种质资源的创新和高效利用具有重要意义[14]。关于种质资源表型多样性研究已在多种植物中具有相关报道,在睡莲中最早为1905年Henry Conad采用传统形态分类法建立的睡莲属植物种的分类系统[1]。此后,李淑娟等[15]利用统计学方法对引进的56个耐寒睡莲品种在关中地区的各个观赏指标和适应性进行了评价,并从中筛选出38个优良品种。吉建斌等[16]在京津冀地区从引进的30多个耐寒睡莲中选出18个性状相对比较稳定的品种,研究了其生物学特性、物候期、抗逆性等。这些研究为睡莲形态评价指标的制定奠定了的基础,但评价的品种有限,大量的热带睡莲品种、尤其是近两年广受欢迎的澳大利亚睡莲未被包纳进去。本研究以海南引进的64个热带睡莲品种及22个广温带睡莲品种为试验材料,采用表型性状分析和数量分类的方法[17]进行研究,旨在为进一步开展睡莲种质资源鉴定、利用及睡莲资源数据库的构建提供依据。
1 材料与方法
1.1 材料
本试验所用睡莲品种均为海南佛渡莲源生态农业有限公司引进品种(表1),种植地点在海口市种质资源圃(110°2928.58 E,19°5355.33 N)。均为地栽,其中广热带亚属睡莲株行距为2 m× 2 m,水深0.2 m;古热带亚属及澳大利亚睡莲株行距均为2 m×3 m,水深0.3~0.5 m;广温带睡莲株行距为1 m×1 m,水深0.3 m,所有睡莲均每年更换底肥,施用复合肥25 kg/666.67 m2、有机肥100 kg/666.67 m2,其中每3个月追施两次叶面肥磷酸二氢钾50 g/666.67 m2,常规技术管理,所用样本且均已栽培、观察近2年,表现良好,各项表型性状趋于稳定。
1.2 性状评价标准制定
主要参照中华人民共和国农业农村部制定的睡莲DUS测试指南及国内外文献资料[15-25],在前试验者评定睡莲、荷花等多种花卉的多样性状评价体系上,优化并增加了部分评价性状,制定了一套含45个主要表型性状指标的评价体系(表2),包含花色、叶长、叶宽、花蕾形状、根茎类型等。各指标观察样本数均不少于10个,均根据表2标准进行各项指标测定记录。
1.3 数据处理
通过SPSS 25.0软件与Microsoft Excel 2007软件进行数据处理与计算分析。
计算不同质量性状各级所占有的频率及根据不同等级数值计算遗传多样性指数(H′)进行质量性状多样性分析。数量性状多样性分析首先计算平均值( )、标准差(s),进一步计算变异系数(CV),以平均值与标准差为依据对所有的数量性状进行等级分类,共分为10级,第一级 ,直到 为第10级[3, 20, 24]。第2~9级,每级即在上一级的基础上增加0.5 s。计算每一级的相对频率(Pi),进而用Pi计算各性状的多样性指数。多样性指数公式采用 [3, 20, 24],计算公式如下:
公式中Pi为各级相对频率(某性状第i级别内包含材料数占总材料的百分比),Ln为自然对数。
聚类分析根据平方欧氏距离划分,主成分分析根据特征值大于1.5进行提取,之后根据各表型性状的新综合指标(特征向量值)为依据,进一步来确定各主成分的权重,构建综合得分函数方程,根据得分进行先后排名,此外还进行了相关性分析。
2 结果与分析
2.1 睡蓮种质资源数量性状多样性分析
由表3可知,13个数量性状多样指数范围为0.612~2.041,其中,多样性指数排在前六位的分别为花瓣宽度(WP)、叶裂深度(DD)、叶长(LL)、萼片宽度(WS)、花瓣长度(LP)和花径(DF),多样性指数分别为2.041、2.019、2.011、2.011、1.994和1.979。多样性指数排在最后一位的是萼片枚数(SN),为0.612。13个数量性状的变异系数范围为19.98%~122.06%,其中强变异的有两个,分别是雄蕊个数(NS)和花瓣总数(NP),分别为122.06%和109.87%,其中澳大利亚睡莲系列的雄蕊个数[平均值为(533±165)枚]明显高于其他品种[(81±66)枚];变异系数最小的是花瓣长度(LP),其次是萼片长度(LS),分别为19.98%和22.65%。
2.2 睡莲种质资源质量性状多样性分析
由表4可知,32个质量性状的多样性指数范围为0.259~1.648。其中柱头盘颜色(SDC)和中轴突颜色(AC)多样性指数最小,为0.259;其次为心皮着生状态(CT)和开花习性(FH),为0.262;萼片外侧主色(COS)、花朵是否挺水(EA)和花梗上下是否均匀(PDV)的多样性指数也较低,分别为0.343、0.367和0.486。花色(FC)的多样性指数最高,为1.648,其他多样性指数较高的性状还有雄蕊类型(ST)和叶型(LS),分别为1.453和1.355。总体来看,花部质量性状多样性总体大于叶片部位,性状中涉及颜色级别的多样性指数总体大于其他性状,说明花部的遗传多样性大于叶片部位的遗传多样性,其次,涉及颜色级别的性状遗传多样性较其他性状丰富。
从不同质量性状的频率分布来看,不同性状的不同类型分布差异很大。一般来说,相对频率比较大的,相对数量较多,也更普遍一些。如根茎类型(RT)共有6个级别,其中相对频率最大的为菠萝型,为0.66;其次为玛利亚克型,占比例为0.13;依次又为香水月季型、走茎型、指型,所占比例分别为0.08、0.07、0.05;结节型所占比例最小,为0.01。又如心皮着生状态(CT)中离生和合生的相对频率分别是0.93和0.07,差异更明显。
2.3 睡莲种质资源聚类分析
利用SPSS数据统计软件和离差平方和法,对86份睡莲材料表型性状进行聚类,在欧式距离14处将供试资源可分成4类(图1)。
第Ⅰ类是广热带睡莲品种群,由‘蓝蟹爪‘多贝‘绿烟‘瑞秋等52份资源组成,主要特征为心皮离生,叶缘锯齿状,白天开花,萼片多呈革质,萼片外侧主色多呈绿色,根茎类型多呈菠萝型,叶基多呈“V”型开裂,花朵多为挺水,柱头盘、中轴突、心皮附属物颜色多呈黄色。可进一步划分为6个亚类:Ⅰ-1亚类为广热带非胎生(蟹爪单复瓣)睡莲,包括5份材料,其共同特点是叶无胎生,雄蕊无瓣化或半瓣化,萼片8枚,花形态碗状,花蕾卵形;Ⅰ-2亚类为广热带非胎生(小花单复瓣)睡莲,包括7份材料,其共同特点是叶无胎生,花朵较小,雄蕊无瓣化或半瓣化;Ⅰ-3亚类为广热带非胎生(中大花单复瓣)睡莲,包括7份材料,其共同特点是叶无胎生,花朵中大型,雄蕊无瓣化或半瓣化;Ⅰ-4亚类为广热带非胎生(小花重瓣)睡莲,包括7份材料,其共同特点是叶无胎生,花朵较小,雄蕊完全瓣化;Ⅰ-5亚类为广热带非胎生(中大花重瓣)睡莲,包括6份材料,其共同特点是叶无胎生,花朵中大型,雄蕊瓣化;Ⅰ-6亚类为广热带胎生(中大花单复瓣)睡莲,包括20份材料,其共同特点是叶胎生,花朵中大型,雄蕊无瓣化或半瓣化。
第Ⅱ类是澳大利亚睡莲品种群,由澳洲IM蓝白、变色澳洲、澳洲IM紫白、澳洲白巨、‘吉祥‘海星6份资源组成,其叶无胎生,花朵大型,雄蕊无瓣化或半瓣化,心皮离生,白天开花,叶片硕大,叶无斑点,雄蕊个数多,萼片4枚,花朵香味不明显,花蕾及外侧花瓣形状均呈卵形,萼片、花瓣均无斑点,萼片外侧主色为绿色,叶梗颜色为褐色,花梗上下不均匀,根茎类型菠萝型。
第Ⅲ类是古热带睡莲品种群,由‘红色闪耀、喀麦隆、‘风月‘粉史龙‘喜庆‘泰国粉6份资源组成,其叶无胎生,花朵大型,雄蕊无瓣化或半瓣化,心皮合生,夜晚开花,萼片4枚,萼片、花瓣均无斑点,萼片外侧主色多为褐色,花朵挺出水面,花蕾形状卵形,叶梗颜色为褐色,花梗上下不均匀,根茎类型为走茎型。
第Ⅳ类是广温带睡莲品种群,由‘油画女郎‘粉莲蓬‘疯狂的雪‘粉红洒金等22份资源组成,主要特征为叶无胎生,心皮离生,叶缘全缘,白天开花,花药颜色黄色,中轴突形状锥形或圆球形,柱头盘、中轴突、心皮附属物颜色均为黄色,萼片外侧主色为绿色。可进一步划分为3个亚类:Ⅳ-1亚类为广温带亚属小花(单复瓣)睡莲,包括‘小海和‘小白子午莲两份材料,其共同特点是花朵小型,雄蕊无瓣化或半瓣化;Ⅳ-2亚类为广温带跨亚属中大花(单复瓣)睡莲,包括6份材料,均为跨亚属杂交睡莲,其共同特点是花朵中大型,雄蕊无瓣化或半瓣化;Ⅳ-3亚类为广温带亚属中大花(单复瓣)睡莲,包括14份材料,其共同特点是花朵中大型,雄蕊無瓣化或半瓣化。
2.4 睡莲资源表型性状间的相关性分析
由表5可知,45个表型性状间存在不同程度相关性。相关系数均大于0.900的有以下6组,分别为叶长(LL)与叶宽(WL)、叶长(LL)与叶裂深度(DD)、叶宽(WL)与叶裂深度(DD)、花径(DF)与花瓣长度(LP)、花瓣长度(LP)与萼片长度(LS)、心皮着生状态(CT)与开花习性(FH),均呈极显著性正相关(P<0.01),相关系数分别为0.995、0.927、0.914、0.946、0.964、1.000。其他性状中,相关系数达到0.800以上的有以下3组,分别为花径(DF)与萼片长度(LS)、叶基(LB)与叶基裂缺程度(DLF)、花瓣宽度(WP)与萼片宽度(WS),之间均呈极显著正相关,相关系数分别为0.865、0.856、0.814。因此,花瓣、萼片的大小可作为花径(DF)大小的判断依据,叶宽(WL)和叶裂深度(DD)可作为叶长(LL)的判断依据,心皮着生状态(CT)可作为开花习性(FH)的判断依据。
2.5 睡莲种质资源表型性状的主成分分析
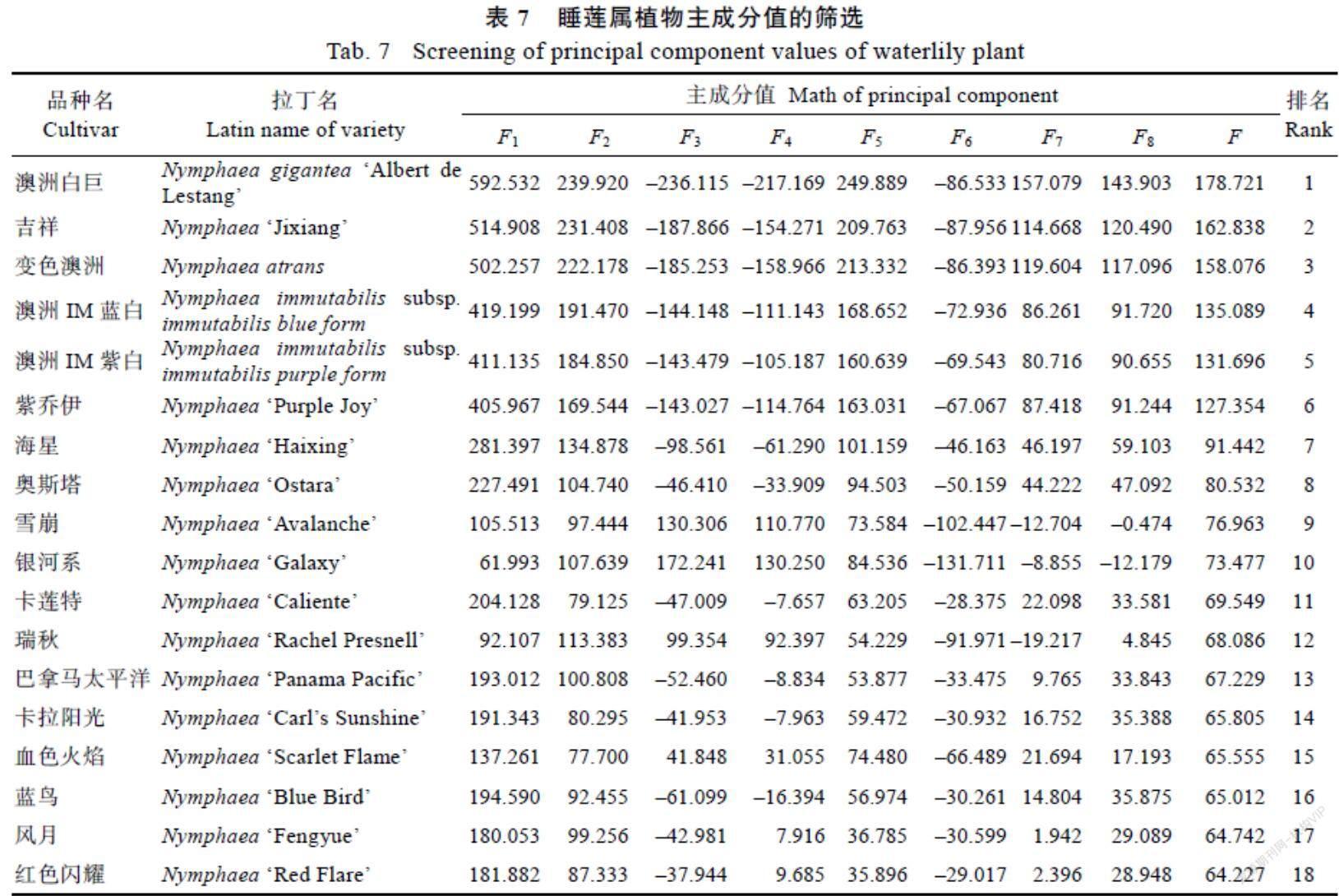
将供试86种睡莲的45个性状数据作为对象,进行统一标准化,以标准化后的性状数据作为主成分分析变量,主成分按照特征值提取,标准为大于1.5,结果(表6)提取主成分8个,可描述45个性状基本特征,累积贡献率60.838%。第1主成分的贡献率最大,为16.150%,特征向量值较大的性状是叶长(LL)、叶宽(WL)、叶裂深度(DD)、花径(DF)和花瓣长度(LP)、萼片长度(LS),其特征向量均为正值且高达0.700以上,此组较多反映了花和叶的大小,说明花和叶大小与睡莲长势密切相关。第2主成分的贡献率为10.702%,特征向量在0.600以上的有花形态(FS),特征向量最小的为叶基(LB),为–0.658。第3主成分的贡献率为8.149%,特征向量较大的有花瓣总数(NP)、萼片枚数(SN)、萼片宽度(WS)。第4主成分的贡献率为7.348%,特征向量较大的有叶色(LC)、叶斑点数(FL)、花梗有无绒毛(PF)。第5主成分的贡献率为6.623%,特征向量较大的有萼片外侧主色(COS)、花色(FC)。第6主成分的贡献率为4.507%,特征向量较大的有雄蕊瓣化(SP)和萼片形状(SS)。第7主成分的贡献率为3.892%,特征向量较大的有萼片内侧颜色(CIS)和叶梗颜色(LSC)。第8主成分的贡献率为3.467%,特征向量较大的有中轴突颜色(AC)、柱头盘颜色(SDC)。
2.6 睡莲种质资源表型性状的综合评价
以8个新综合指标(主成分特征向量)代替45个性状指标对86种睡莲进行综合评价,首先计算 (n为1~8主成分值)值,进一步计算各品种综合主成分值F并排序,F值权重为每个主成分对应特征值与提取主成分总特征值累加和的比值。函数表达式为:F1=0.843x1+0.825x2+ 0.799x3+…+0.367x45,并依此类推,每个品种共8个主成分值,进一步计算综合主成分值,以各主成分所对应方差贡献率为权数,F=(0.162F1+ 0.107F2+0.081F3+0.073F4+0.066F5+0.045F6+0.039F7+0.035F8)/0.6084,最后根据F值进行排名。将供试86种睡莲前20%的品种列于表7,共18个品种,分别为广热带亚属睡莲10个(占本亚属的19%),所有澳大利亚睡莲共6个,古热带亚属睡莲2个,其中‘澳洲白巨‘吉祥和‘变色澳洲排于前三名。
3 讨论
植物表型多样性是遗传多样性与环境多样性的综合体现,对表型性状的研究可以从整体上明确研究对象的多样性程度[25-26],也对综合农艺性状潜力和育种工作起到至关重要的作用[27]。目前应用表型多样性数据来进行遗传评价、构建种质资源库,筛选特定品种的资源已经在牡丹[28]、柿花[29]、山茶花[30]、栀子[31]、核桃[32]、大白花杜鹃[33]、油茶[34]等花木资源中得到了应用[17]。本研究对86份睡莲的45个表型性状进行分析,发现数量性状多样性大于质量性状,多样性指数与变异系数的大小呈相反趋势,与苏群等[17]、张海平等[20]所分析结果较为一致;变异系数范围为11.79%~ 122.06%,排名前两位的为雄蕊个数(NS)和花瓣总数(NP),与张海平等[20]的分析结果为花药颜色(AC)不一致;与苏群等[17]所分析的结果虽一致,但不同的是本试验中这两种性状均属于强变异类型,说明生长环境很可能对睡莲部分器官生长发育具有极显著的影响。相比前人研究[15-21],本研究选取的古热带亚属及全瓣化品种睡莲均相应增多,古热带亚属睡莲花瓣数和雄蕊数多于多数品种。因此在今后试验中应充分考虑不同亚属睡莲资源的综合性状,合理取材。
聚类分析将供试睡莲分为4个类群,依次为:广热带睡莲品种群、澳大利亚睡莲品种群、古热带睡莲品种群、广温带睡莲品种群。这与睡莲的亚属亲缘关系相一致。在亲缘关系划分的基础上,依据叶片是否胎生、花型大小和雄蕊瓣化程度进一步划分了亚类(图1)。根据分类结果,建议可以参照梅花[35]等观赏植物的二元分类法,构建睡莲的品种分类体系:第一级分类标准为睡莲亲缘关系,即广热带、澳大利亚、古热带、广温带和新热带5亚属;第二级分类标准为叶片是否具有胎生能力,分为胎生与非胎生两系;第三级分类标准为花大小,包括小花和大中花两类;第四级分类标准为花型,包括蟹爪、单复瓣(雄蕊不瓣化或少数瓣化)及重瓣(雄蕊全部瓣化)三型。此分类体系反映了睡莲总的亲缘关系,又方便园艺应用,具有科学实用的特点。若要更准确反映睡莲属内的植物亲缘关系,还需结合现代生物学技术的方法进一步验证[20]。需要特别指出的是,近年来我国大力引进的澳大利亚睡莲,在本研究中表现出了独特的表型,如雄蕊数量极多,本研究中测定澳大利亚类型的睡莲雄蕊数平均为(533±165)枚,远高于其他类型(81±66)枚;同时其花药基部为丝状,上部膨大呈花药状,分化比较明显,与其他品种表型差异明显(暂命名为澳大利亞型雄蕊)。聚类分析中将其单独聚为一类,说明这些性状具有明确的分类学意义。目前睡莲属DUS测试指南尚未纳入这些性状,建议在未来的更新中,澳大利亚类型睡莲的独特性状也应纳入形态指标中。
性状相关性可以通过一种性状的表型间接反映另一种性状的表型,从而提高选择效率,加快育种进程[36]。相关性分析表明,不同品种叶片大小与花朵大小相关的性状、心皮着生状态(CT)与开花习性(FH)均呈极显著正相关(表5)。说明用叶片大小预测花瓣大小,具有一定的参考价值。心皮离生状态睡莲均为白天开花,心皮合生状态睡莲均为夜晚开花。总体而言,心皮着生状态(CT)与开花习性(FH)之间的关系符合本研究规律。通过主成分分析,提取了8个主成分,与前人研究相比[13, 15-21],主成分提取值相对增多,这是由于本试验增加了有关雄蕊、柱头盘、中轴突等部位具体性状,如第8主成分中中轴突颜色、柱头盘颜色特征向量较大,在描述总体花部特征的同时,能进一步具体描述到局部器官的特征。
总体而言,本研究通过对睡莲品种的表型性状多样性进行分析,进一步明确了睡莲表型变异的丰富程度,为睡莲优质种质资源的保存以及核心种质资源数据库的构建提供了一定的理论基础,为睡莲资源的利用提供了一定的参考,也为培育睡莲新品种提供了相应的理论依据。
参考文献
[1] 黄国振, 邓惠勤, 李祖修, 等. 睡莲[M]. 北京: 中国林业出版社, 2009: 1-10.
[2] 李淑娟, 尉 倩, 陈 尘, 等. 中国睡莲属植物育种研究进展[J]. 植物遗传资源学报, 2019, 20(4): 829-835.
[3] Yin D D, Yuan R Y, Wu Q, et al. Assessment of flavonoids and volatile compounds in tea infusions of water lily flowers and their antioxidant activities[J]. Food Chemistry, 2015, 187: 20-28.
[4] Yildirim A B, Karakas F P, Turker A U. In vitro antibacterial and antitumor activities of some medicinal plant extracts, growing in Turkey[J]. Asian Pacific Journal of Tropical Medicine, 2013, 6(8): 616-624.
[5] Thippeswamy B, Mishra B, Veerapur V, et al. Anxiolytic activity of Nymphaea alba Linn. in mice as experimental models of anxiety[J]. Indian Journal of Pharmacology, 2011, 43(1): 50-55.
[6] Bhandarkar M R, Khan A. Antihepatotoxic effect of Nymphaea stellata willd., against carbon tetrachloride induced hepatic damage in albino rats[J]. Journal of Ethnopharmacology, 2004, 91(1): 61-64.
[7] Zhang Z Z, ElSohly H N, Li X C, et al. Phenolic compounds from Nymphaea odorata[J]. Journal of Natural Products, 2003, 66(4): 548-550.
[8] Dhanabal S P, Raja M K M M, Ramanathan M, et al. Hypoglycemic activity of Nymphaea stellata leaves ethanolic extract in alloxan induced diabetic rats[J]. Fitoterapia, 2007, 78(4): 288-291.
[9] Rajagopal K, Sasikala K. Antidiabetic activity of hydroethanolic extracts of Nymphaea stellata flowers in normal and alloxan induced diabetic rats[J]. African Journal of Pharmacy and Pharmacology, 2008, 2(8): 173-178.
[10] Bing S J, Ha D, Kim M J, et al. Geraniin down regulates gamma radiation-induced apoptosis by suppressing DNA damage[J]. Food and Chemical Toxicology, 2013, 57: 147- 153.
[11] 余翠薇, 陳煜初, 余东北, 等. 睡莲5个亚属花、叶、块茎和基因组大小比较[J]. 浙江农业科学, 2017, 58(8): 1353-1356, 1361.
[12] 王天友, 王有武, 曹新川, 等. 南疆陆地棉种质资源表型性状遗传多样性分析[J]. 种子, 2020, 39(4): 5-11.
[13] Sterpansky A, Kovalski I, Perltreves R. Intraspecific classification of melons (Cucumis melo L.) in view of their phenotypic and molecular variation[J]. Plant Systematics and Evolution, 1999, 217(3-4): 313-332.
[14] 王永康, 吴国良, 赵爱玲, 等. 枣种质资源的表型遗传多样性[J]. 林业科学, 2014, 50(10): 33-41.
[15] 李淑娟, 樊 璐, 原雅玲, 等. 关中地区引进耐寒睡莲品种观赏性状评价及优良品种筛选[J]. 陕西林业科技, 2010(3): 15-18.
[16] 吉建斌, 李清清, 黄娟英, 等. 京津冀耐寒睡莲品种引繁及栽培技术研究(1)——评分标准及生物学特性研究[J]. 林业实用技术, 2007(3): 3-5.
[17] 苏 群, 杨亚涵, 田 敏, 等. 49份睡莲资源表型多样性分析及综合评价[J]. 西南农业学报, 2019, 32(11): 2670-2681.
[18] 张孟锦, 杨志娟, 严 海, 等. 热带地区睡莲新品种的优选及栽培技术研究[J]. 热带农业科学, 2017, 37(10): 31-35.
[19] 万里波, 王奎玲, 刘庆华, 等. 青岛耐寒睡莲品种数量分类研究[C]//中国观赏园艺研究进展(2014). 青岛: 中国林业出版社, 2014: 133-139.
[20] 张海平, 房伟民, 陈发棣, 等. 部分睡莲属植物形态性状的多样性分析[J]. 南京农业大学学报, 2009, 32(4): 47-52.
[21] 陈发棣, 崔娜欣, 丁跃生. 南京地区新引耐寒睡莲主要观赏性状初步评价[J]. 上海农业学报, 2002(3): 51-55.
[22] 原 鑫, 李文玲, 刘召强, 等. 荷花品种表型性状遗传多样性分析[J]. 江苏农业科学, 2020, 48(16): 188-193.
[23] 贾东海, 王秀珍, 侯献飞, 等. 32份油药兼用红花种质资源表型性状遗传多样性分析[J]. 新疆农业科学, 2020, 57(10): 1775-1784.
[24] Long J C, Zhang J J, Zhang X C, et al. Genetic diversity of common bean (Phaseolus vulgaris L.) germplasm resources in Chongqing, evidenced by morphological characterization[J]. Frontiers in Genetics, 2020, 11: 697.
[25] 徐 君, 李 欣, 江 君, 等. 基于表型和ISSR标记的小株型荷花遗传多样性分析[J]. 江苏农业科学, 2019, 47(19): 137-141.
[26] 袁东升, 王晓敏, 赵宇飞, 等. 100份番茄种质资源表型性狀的遗传多样性分析[J]. 西北农业学报, 2019, 28(4): 594-601.
[27] Scarano D, Rubio F, Ruiz J J, et al. Morphological and genetic diversity among and within common bean (Phaseolus vulgaris L.) landraces from the Campania region (Southern Italy)[J]. Scientia Horticulture, 2014, 180: 72-78.
[28] 张 琳, 郭丽丽, 郭大龙, 等. 牡丹杂交F1代性状分离规律及混合遗传分析[J]. 南京林业大学学报(自然科学版), 2018, 42(6): 51-60.
[29] 索玉静, 孙 鹏, 韩卫娟, 等. 柿雄花表型及花粉形态多样性研究[J]. 中国农业大学学报, 2019, 24(2): 48-60.
[30] 李 芳, 霍 达, 王 进. 西南红山茶花表型性状的变异[J]. 贵州农业科学, 2019, 47(4): 84-88.
[31] 邓绍勇, 杨 欢, 朱培林, 等. 栀子栽培品种与近缘种的数量分类[J]. 江西农业大学学报, 2020, 42(1): 92-100.
[32] 申艳红, 高兴泉, 石天磊, 等. 石门核桃种质资源坚果品质评价与分析[J]. 北方园艺, 2020(18): 43-48.
[33] 李叶芳, 马诗雨, 宋 杰, 等. 大白花杜鹃三个天然居群的表型多样性分析[J]. 北方园艺, 2019(1): 115-120.
[34] 王 海, 周 扬, 邓雅洁, 等. 海南油茶优异种质资源及其品质的分析评价[J]. 分子植物育种, 2021, 19(12): 4142-4152.
[35] 陈俊愉, 陈瑞丹. 中国梅花品种群分类新方案并论种间杂交起源品种群之发展优势[J]. 园艺学报, 2009, 36(5): 693-700.
[36] 刘子记, 申龙斌, 杨 衍, 等. 甜椒核心种质遗传多样性与亲缘关系分析[J]. 江苏农业科学, 2016, 44(5): 199-202.
责任编辑:沈德发
猜你喜欢
中国房地产·学术版(2016年10期)2016-11-18
大学教育(2016年11期)2016-11-16
大经贸(2016年9期)2016-11-16
中小企业管理与科技·上旬刊(2016年10期)2016-11-15
考试周刊(2016年84期)2016-11-11
中国市场(2016年33期)2016-10-18
科技视界(2016年20期)2016-09-29
商业经济研究(2016年14期)2016-09-14
企业导报(2016年9期)2016-05-26
