
利用染色体片段置换系定位低温影响水稻萌芽期根长和芽长QTL
2021-12-17 07:20余镁霞邓浩东谭景艾宋贵廷吴光亮陈利平刘睿琦邹安东贺浩华边建民
作物杂志 2021年6期
余镁霞 邓浩东 谭景艾 宋贵廷 吴光亮 陈利平刘睿琦 邹安东 贺浩华,2 边建民,2
(1作物生理生态与遗传育种教育部/江西省重点实验室,330045,江西南昌;2江西省水稻高水平工程研究中心,330045,江西南昌)
水稻起源于热带和亚热带,对低温敏感。随着人口数量的增加,对水稻的需求量增加,导致水稻种植面积不断向高纬度低温地区扩展[1]。低温不仅影响水稻内部代谢物的产生,如脯氨酸、蔗糖和活性氧等,还会影响根和芽的形态[2]。水稻主要依靠根和芽的通气组织输送氧气[3],根和芽发育不良会使植株整体缺氧,不利于产量形成。因此,在低温条件下,研究水稻萌发期根和芽的遗传机制对培育耐冷优良品种和促进水稻生产具有重要指导作用。
水稻耐冷性是一个复杂的数量性状,可以通过较多指标衡量[4-5]。迄今为止,已通过存活率、发芽率、发芽力、种子活力等指标定位到多个与水稻耐冷相关的QTL,如qCTS12[6]、qLTG3-1[7]、COLD-1[8]、qLTB3[9]和qCTB7[10]等。但对衡量水稻耐冷性重要指标的根长和芽长的研究相对较少。
在水稻根系众多性状中,根长是一个能反映水稻在逆境条件下生长发育健壮程度的重要指标。Steele等[11]在灌溉和干旱胁迫处理下以近等基因系(near-isogenic line,NIL)为材料,在第9号染色体 RM242-RM201间定位到 1个根长 QTL。Shimizu等[12]在磷缺乏的条件下,以籼稻品种Kasalath和粳稻品种日本晴构建的染色体片段置换系(chromosome fragment replacement line,CSSL)群体为材料,在第 6号染色体定位到影响根长的QTLqREP-6。Mitsuhiro等[13]通过更换以 NH4+为唯一氮源的营养液建立适合量取根长的生长条件,利用粳稻品种 Koshihikari和籼稻品种 Kasalath杂交衍生出的CSSL,在第6号染色体检测出控制根长的QTLqRL6.1。Wang等[14]利用短根保持系协青早B和长根恢复系 R9308杂交获得的重组自交系(recombinant inbred line,RIL)群体,在第7号染色体检测到影响根长的QTLqRL7。徐晓明等[15]以超级稻协优 9308衍生 RIL群体与轮回亲本中恢9308回交多代的高代回交群体,在第4号染色体定位到影响根长的QTLqRL4。赵春芳等[16]在低磷条件下,以 9311为背景亲本、日本晴为供体亲本衍生的染色体片段置换系为材料,在第5号染色体定位到1个与根长相关的QTL。
与根长相比,芽长作为另一个反映水稻生长发育健壮程度的重要指标,前人对其研究甚少。Redona等[17]利用粳稻Labelle和籼稻Black Gora衍生的RIL群体,在18℃和25℃温度条件下,定位到4个影响芽长的QTL。Fukuda等[18]通过籼稻和粳稻杂交的近交系在低温16℃条件下,检测到改善芽生长的QTLqSL3和qSL8。Zhang等[19]利用粳稻Lemont和籼稻Teqing杂交衍生的RIL,在18℃条件下定位到4个影响芽长的主效QTL。此外,班超等[20]通过整理水旱条件下水稻根系性状QTL,得到水稻第1、2、3、5和11号染色体共11个区域很可能存在“一因多效”位点。这表明水稻根和芽可能存在协调发育的可能。但是对水稻根长和芽长的遗传表型及其二者关系,特别是低温条件下的关系知之甚少。
基于此,本研究利用9311(受体)/日本晴(供体)衍生的CSSL群体,在正常温度(25℃)和低温(15℃)条件下,分析水稻萌芽期根长和芽长的遗传表现,并定位其中影响水稻萌芽期根长和芽长QTL。同时分析QTL间的关系,为后续研究其遗传机制提供参考。
1 材料与方法
1.1 材料与试剂
从受体亲本籼稻 9311和供体亲本粳稻日本晴杂交、回交的后代群体中选出121个性状稳定的株系,利用全基因组重测序技术进行基因分型,获得高质量的Bin标记;同时结合JoinMap 4.2软件构建CSSL群体的高密度图谱。
1.2 试验方法
从亲本和每个株系所收获的种子中选取 40粒饱满健康的种子,经3.5%次氯酸钠消毒后,用蒸馏水漂洗干净。随后将种子置于底部放有一层滤纸的9cm无菌培养皿中,每个培养皿中加入恰好浸没种子的蒸馏水。并放置于28℃条件下催芽。待种子萌发后,选择其中整齐一致的30粒种子,置于15℃、12/12h(昼/夜)恒温培养箱处理 7d,然后在 25℃恢复生长7d,期间每天随机选取10粒幼芽测量根长(cm)和芽长(cm),测量后放回培养皿。
1.3 统计分析
采用 IciMapping V4.2软件完备区间作图法(ICIM)对 CSSL群体进行数据分析和 QTL定位[21-22]。QTL按照q+目标性状+所在染色体号数或连锁群代号原则命名,名称用斜体[23-24]。用 SPSS 22.0与Excel 2017进行显著性分析。
2 结果与分析
2.1 遗传图谱构建
利用多态性标记对 CSSL群体基因型进行划分,与日本晴相同的基因型转化为标记2,与9311相同的基因型转化为标记0,无法判断的基因型转化为标记-1。以100kb为一个bin窗口,滑动所有子代样品的每条染色体,根据每个样品基因型来源,以 bin为单位得到该群体每个子代 marker信息,最终获得655个Bin标记(表 1)。利用这些Bin标记构建染色体片段置换系图谱,其中约97%的基因组被日本晴片段置换覆盖(图1)。
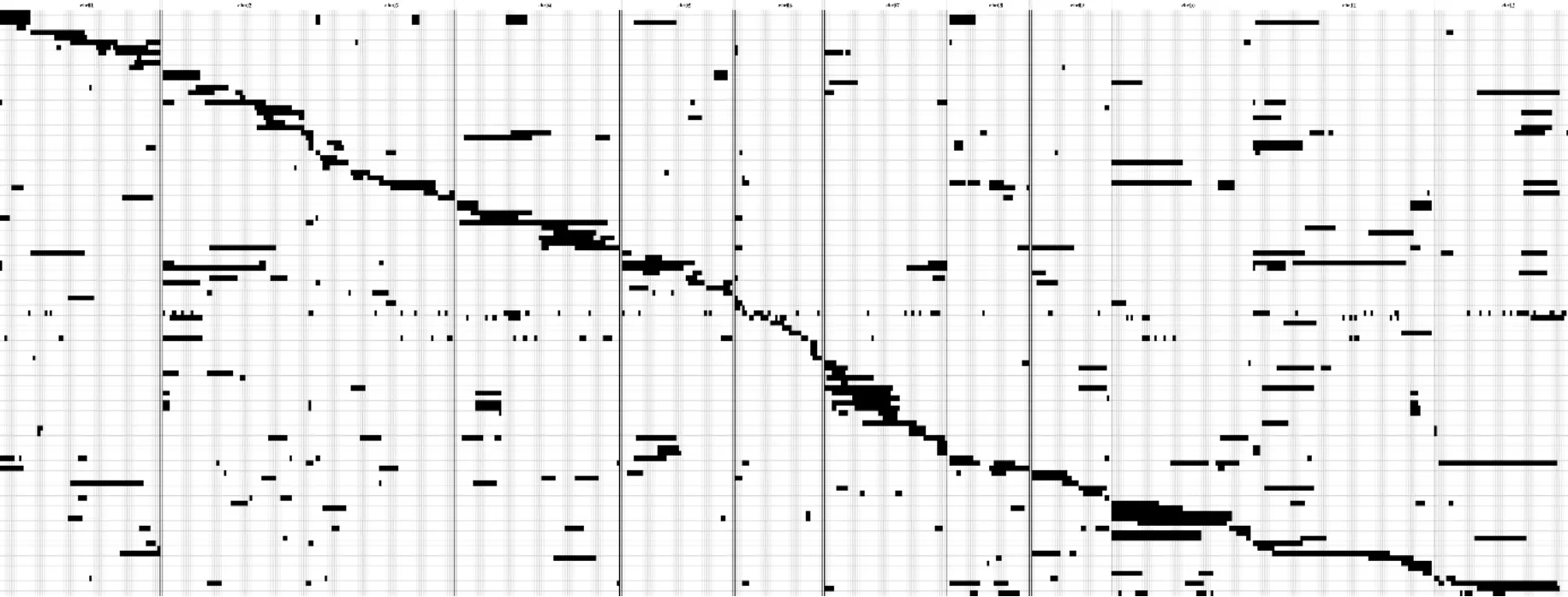
图1 染色体片段置换系图谱基因型图示Fig.1 Graphical genotypes of the CSSLs

表1 染色体标记数及bin间遗传距离Table 1 The bin marker numbers and genetic distance between adjacent bins Mb
2.2 2个温度下亲本和CSSL群体表型分析
由表2得,15℃条件下,2个亲本根长在第1天和第2天、芽长在第6天和第7天存在显著差异,其余生长时期2个亲本根长或芽长差异不显著。25℃条件下,2个亲本的根长在第1、2、3、4天、芽长在第3、4、5、6天存在显著差异,其余生长时期差异不显著。CSSL群体中,幼芽根长和芽长均表现出不同程度的差异,在2个温度条件下均呈现连续变异,具有广泛的变异幅度,均表现出明显的超亲分离现象(表2和图2)。表明15℃和25℃条件下,水稻幼芽的根长和芽长表现出明显的数量性状特征,符合QTL定位要求。

图2 15℃和25℃处理下CSSL群体表型分布Fig.2 Phenotypic distribution of CSSL population at 15℃ and 25℃

表2 亲本及CSSL群体在2个温度条件下根长和芽长表型分析Table 2 Phenotype analysis of root length and bud length under two temperatures of their parents and CSSL population
2.3 2个温度条件下根长和芽长的QTL定位
2个温度条件下共定位32个QTL。其中,18个QTL与根长相关,14个QTL与芽长相关。这些QTL分布在水稻的第1、2、3、6~12号染色体上,贡献率为0.58%~38.26%。大部分QTL(75%)分布在水稻的第3、7~10号染色体(表3和图3)上。有9个QTL增效等位基因来自日本晴,其余均来自9311。

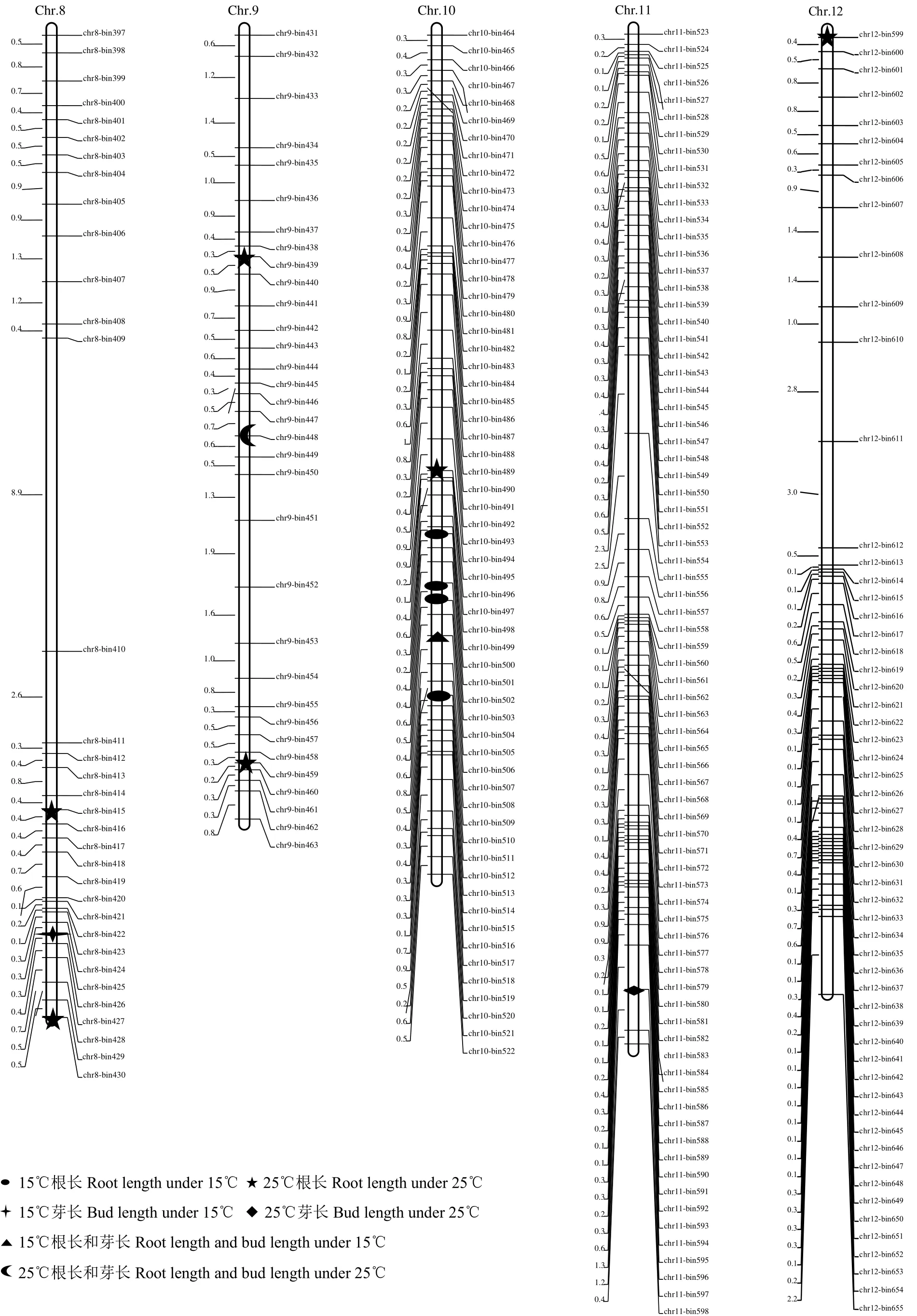
图3 CSSL群体定位的2个温度处理后影响根长和芽长QTL染色体分布Fig.3 Distribution of QTL related root length and bud length on genetic map under two temperature treatments of CSSL population
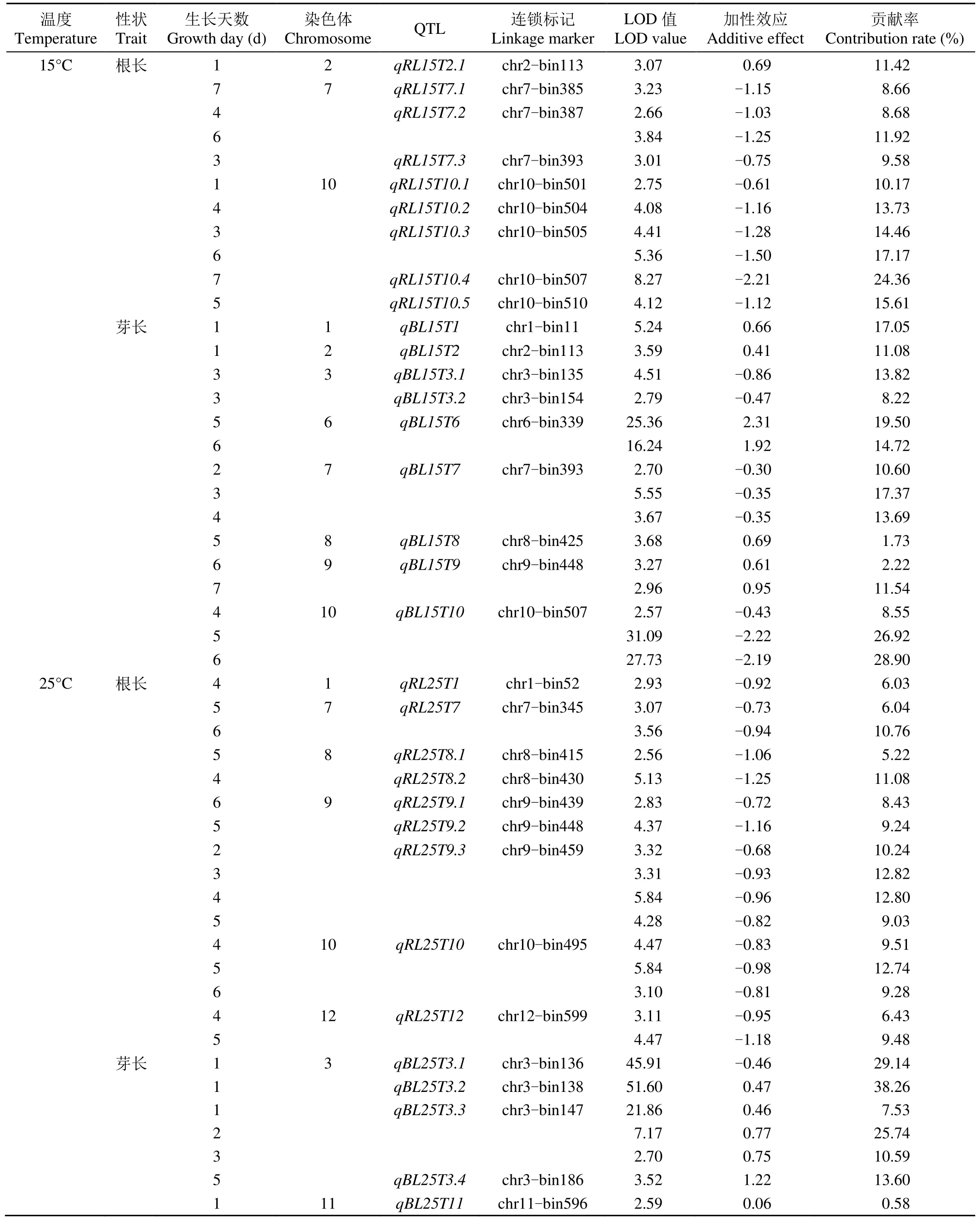
表3 CSSL群体定位2个温度条件下幼芽生长不同时期影响根长和芽长的QTLTable 3 QTL mapping for root length and bud length under two temperatures using CSSL population at different stages of sprout growth
2.3.1 15℃条件下 QTL定位 15℃条件下,共定位到 9个根长 QTL:qRL15T2.1、qRL15T7.1、qRL15T7.2、qRL15T7.3、qRL15T10.1、qRL15T10.2、qRL15T10.3、qRL15T10.4和qRL15T10.5,贡献率为8.66%~24.36%。其中,qRL15T7.2和qRL15T10.3在不同时期被重复检测到。除qRL15T2.1增效等位基因来自于日本晴外,其余均来自9311。
15℃条件下,定位到9个芽长QTL,分别位于第 1~3、6~10号染色体上,贡献率为 1.73%~28.90%,其中qBL15T6、qBL15T7、qBL15T9和qBL15T10被重复检测到。qBL15T3.1、qBL15T3.2、qBL15T7和qBL15T10促进芽生长的等位基因来自9311,而qBL15T1、qBL15T2、qBL15T6、qBL15T8和qBL15T9促进芽生长的等位基因来自日本晴。
2.3.2 25℃条件下 QTL定位 25℃条件下,在 6条染色体共检测到9个影响根长的QTL,贡献率为5.22%~12.82%。所有位点上促使根生长的等位基因均来自9311。qRL25T7、qRL25T9.3、qRL25T10和qRL25T12被重复检测到。
25℃条件下,有5个影响芽长的QTL被检测到。其中qBL25T11位于第 11号染色体上,qBL25T3.1、qBL25T3.2、qBL25T3.3和qBL25T3.4分布于第3号染色体。QTL贡献率为0.58%~38.26%。只有qBL25T3.3在芽生长不同时期被重复检测到。除qBL25T3.1促进芽生长的等位基因来自9311,其余位点增效等位基因均来自日本晴。
2.4 多效性QTL分析
由表4可知,第2、7、9、10号染色体的4个Bin标记区域存在同一温度影响不同性状或不同温度影响不同性状的位点。标记bin113、bin393和bin507所在的位点在15℃条件下对根长和芽长生长均有影响,这3个位点的增效等位基因均来自9311。标记bin448所在的位点对15℃芽长和25℃根长产生影响,其等位基因分别来源于日本晴和9311。

表4 对性状有多效性的染色体标记对应区域Table 4 Chromosomal markers corresponding to regions with pleiotropic effects on traits
3 讨论
3.1 芽期低温胁迫后水稻恢复生长的遗传机制
为探究芽期低温胁迫后水稻恢复生长的遗传机制,本研究利用 9311/日本晴衍生的CSSL群体在低温(15℃)和正常温度(25℃)下对根长和芽长进行了分析。结果显示,不同亲本在同一温度下存在差异。CSSL群体根长和芽长在萌发期不同阶段均表现出连续变异,表明低温和正常温度下水稻根长和芽长均表现为数量性状遗传。此外,在幼芽生长不同阶段,只有少数 QTL被重复检测到,并且在不同时期其加性效应值不同,表明水稻幼芽期不同阶段基因是选择性表达,其耐低温性受多个QTL(基因)控制,这与陈利华等[25]研究结果类似。
2个温度条件下,生长前期9311根长显著长于日本晴,后期由于粳稻日本晴生长速度加快[26-27],根长差异不显著。生长前期2个亲本间芽长无显著差异,生长后期 15℃条件下日本晴芽长显著长于9311,25℃条件下表现不明显,这可能与粳稻耐冷性强有关。在同一生长时期,2个温度条件下所定位的 QTL均不同,即低温和正常温度下,控制水稻芽期生长的基因(QTL)均存在差异,表明低温处理后,幼芽恢复生长遗传机制与常温条件下正常生长可能存在差异。
3.2 定位的根长和芽长QTL比较
25℃条件下,检测到9个影响根长的QTL,其等位基因均来自于9311,其中影响最大的QTL为qRL25T9.3(LOD值16.75,贡献率44.89%);检测到5个影响芽长的QTL,其中影响最大的QTL为qBL25T3.3(LOD 值 51.60,贡献率 43.86%),除qBL25T3.1外,其余QTL位点增效等位基因均来自于日本晴。结果表明,常温条件下2个亲本等位基因对根长和芽长均产生主要影响。15℃条件下,检测到9个影响根长的QTL,其中影响最大的QTL为qRL15T10.3(LOD值9.77,贡献率31.63%),除qRL15T2.1外,其余QTL位点增效等位基因均来自于9311;检测到9个影响芽长的QTL,qBL15T3.1、qBL15T3.2、qBL15T7和qBL15T10中增效等位基因来自于 9311,而qBL15T1、qBL15T2、qBL15T6、qBL15T8和qBL15T9中增效等位基因来自于日本晴,其中影响最大的 QTL为qBL15T10(LOD值61.39,贡献率 64.37%),其增效等位基因来源于9311。结果表明,籼稻 9311中也存在耐低温的基因。尽管较多的研究表明粳稻较籼稻耐冷,特别是幼苗期和抽穗扬花期,但Fujino等[28]和Miura等[29]研究结果表明,籼稻芽期耐冷性强于粳稻,同时定位了多个籼稻芽期耐冷QTL,这与本研究结果较一致,表明籼稻和粳稻中可能均存在耐冷的基因。因此,可以聚合这些耐低温的基因(QTL),并通过分子标记辅助选择技术将其转入到其他籼稻品种中,使其根和芽能够更好地在低温环境下生长。
本研究共定位了到 32个影响幼芽生长的QTL。其中qRL15T2.1(19.75~20.75 Mb)与 Courtois等[30]定位到影响每蘖根重QTL(RG171-RG157)位置接近。qRL25T1(35.9~38.35 Mb)与Zheng等[31]定位控制根粗(RZ730-RZ801)的QTL位置相近。徐吉臣等[32](C711-G103)研究中也曾提及,qRL25T9.1(6.5~7.25Mb)所在位点控制最大根长。qRL25T9.3(20.6~21.05Mb)所在位点分别影响水稻生长后期根中木质部血管数目及移栽后生根[33-34]。在qBL25T3.2位置分别定位到控制苗长的OsMRP5和oni1[35-36]。此外,目前暂未发现与其他位点相同或接近的与根和芽相关性状的QTL。这些QTL的定位为解析水稻芽期遭遇低温冷害后恢复生长的遗传机制提供了参考。通过聚合这些 QTL使水稻在芽期遭遇低温后更快地恢复生长。
3.3 多效性QTL分析
本研究表明,2个温度条件下,根和芽的生长相互影响。本试验发现标记区域bin113(qRL15T2.1和qBL15T2)、bin393(qRL15T7.3和qBL15T7)及bin507(qBL15T10和qRL15T10.4)同时在15℃条件下对根长和芽长恢复生长有影响,bin448(qBL15T9和qRL25T9.2)对15℃芽长和25℃根长有影响,为多效性QTL,影响根和芽的协同发育。因此将其视为根和芽协调发育中的多效QTL[37]。
4 结论
以9311(受体)/日本晴(供体)CSSL群体为材料,在15℃低温处理后,共检测到9个影响根长及9个影响芽长的QTL。以25℃为对照,共检测到 9个影响根长及 5个影响芽长的 QTL。除qRL15T2.1、qRL25T1、qRL25T9.1、qRL25T9.3及qBL25T3.2外,其余均可能是新的影响根和芽生长的QTL。同时,在第2、7、9和10号染色体上4个Bin标记区域检测到影响根和芽协调发育的多效性QTL。此外,在15℃定位的根长或芽长QTL与在25℃定位的根长或芽长QTL均不相同,表明低温条件下控制水稻幼芽期根和芽生长的遗传机制可能与常温下不同。
猜你喜欢
小小说月刊·下半月(2021年8期)2021-09-05
智慧健康(2021年17期)2021-07-30
国际检验医学杂志(2021年7期)2021-04-15
中国产前诊断杂志(电子版)(2020年1期)2020-05-21
现代园艺(2020年7期)2020-04-22
湖北农业科学(2019年20期)2019-12-11
动漫界·幼教365(大班)(2018年3期)2018-05-14
农业与技术(2018年5期)2018-04-25
农产品加工(2017年19期)2017-11-14
湖南大学学报·自然科学版(2016年12期)2017-05-12
