
枣和酸枣果实可溶性糖积累规律差异研究
2022-01-05 11:00邓群仙张慧芬林立金吕小平廖文飞
四川农业大学学报 2021年6期
邓 倩,王 羊,邓群仙*,张慧芬,夏 惠,林立金,吕小平,廖文飞
(1.四川农业大学园艺学院,成都 611130;2.四川省广安市邻水县经果技术推广所,四川邻水 638500;3.四川农业大学果蔬研究所,成都 611130)
‘罗江调元枣’原产四川盆地高温高湿寡日照地区的德阳市罗江区,果实长圆柱形,糖高酸低,味甜,是四川主栽鲜食枣品种[1]。‘圆形小酸枣’是广泛分布于我国酸枣产区的灌木,果实圆球形,酸高糖低,味酸,为常用药用植物及栽培枣砧木[2]。
糖是果实品质和风味物质的主要成分之一。糖产生水果最重要的口味感觉——甜[3]。但是果实的甜味是由糖的种类、含量及比例共同决定的。不同种类,甚至同一种类不同品种的果实,可溶性糖构成都可能存在差异,多数果实以蔗糖、果糖和葡萄糖为主要可溶性糖[4]。桃中蔗糖含量约占总可溶性糖的73%[5],苹果中果糖含量最高[6]。荔枝有蔗糖积累型、单糖积累型、蔗糖与单糖含量相当等3种类型[7]。‘北京鸡蛋枣’和‘延川酸枣A’(灌木型)成熟时果糖含量最高,为己糖积累型;但‘延川酸枣B’(乔木型)、骏枣和冬枣在成熟期果实蔗糖含量最高[8-10],为蔗糖积累型。
目前果实中可溶性糖代谢的研究主要集中在苹果、葡萄、柑橘以及梨等树种上,北方栽培枣品种‘冬枣’和‘骏枣’的全基因组测序和可溶性糖代谢研究也已见报道[9-12]。但南方鲜食枣品种的可溶性糖积累规律及其相关基因表达的研究较少,且由于南方光照弱、阴雨天多等原因导致南方栽培枣果实中可溶性糖的积累量低于北方栽培枣[13],这对鲜食枣风味的影响较大。新疆和北京地区种植的鲜食枣中可溶性糖的含量最高可达33.35%和37.15%[14-15],而四川种植的‘罗江调元枣’可溶性糖含量仅为22%左右[2]。本试验以四川主栽鲜食枣品种‘罗江调元枣’为材料,以砧木品种‘圆形小酸枣’为对照。通过测定不同发育时期可溶性糖含量与组分,及相关代谢基因的表达量,进一步分析‘罗江调元枣’和‘圆形小酸枣’的可溶性糖积累规律差异,为进一步研究南方栽培枣品种中可溶性糖代谢及枣果实品质与风味的形成提供理论依据。
1 材料和方法
1.1 试验地概况
试验园位于四川省德阳市罗江区白马关镇万佛村五组(N 31°26′34′,E 104°46′18′),海拔约 660 m,属于亚热带湿润型,气候温和,四季分明,年均气温16.5℃,最高气温36.6℃,最低气温-6.7℃;年均降水量910 mm,年无霜期278 d,年均日照时数1 260 h。
1.2 试验材料
试验树为露地栽培鲜食枣品种‘罗江调元枣’(根蘖苗,用TYZ表示),株行距为2 m×3 m;露地栽培砧木酸枣品种‘圆形小酸枣’(根蘖苗,用SZ表示),株行距为2 m×3 m。枣树生长结果正常,树势健壮、栽培管理基本一致。
每份材料分别选择3株树,单株为一小区,3次重复。于第二批花的盛花期,在树冠外围由枣股抽生的枣吊中部挂牌。选择树冠外围与挂牌枣吊(由枣股抽生)上大小相似的果实,置冰盒中带回实验室后立即放入-80℃超低温冰箱中保存。材料皆于花谢后20 d开始采样,前期(幼果期)每隔14 d采样一次,中后期(果实膨大期至半红期)至果实商品成熟期(果面至少有一半为赭红色)每隔7 d采样一次。每次采样每份材料约60个果实。
1.3 可溶性糖的测定
可溶性糖的提取参照柴叶茂[16]的方法,略有改动。用液氮将果实研磨成粉末,准确称取0.5 g粉末放入 10 mL 离心管中,用 5 mL 乙醇-水(80∶20,V∶V)溶解,80℃水浴30 min,取出冷却。将提取样品在常温下6 000 r/min离心10 min,上清液转移到25 mL的容量瓶中,重复提取1次,定容。定容液经过0.22 μm滤膜过滤待测。可溶性糖的测定参照马小卫[17]的方法,略有改动。色谱柱选用Innoval NH2(250 mm×4.6 mm,5 μm),流动相为乙腈-水(80∶20,V∶V),示差检测器,流速1 mL/min,检测池温度40℃;柱温35℃,进样量10 μL。
1.4 总RNA的提取与cDNA合成
RNA的提取参照改良CTAB法,cDNA的合成参照TOYOBO公司的ReverTra Ace qPCR RT Master Mix with gDNA Remover试剂盒说明书进行操作
1.5 PCR引物设计和反转录产物检测
糖代谢引物及内参基因引物序列参考张春梅[9]报道的,序列见表 1。利用Primer Premier 5.0设计定量引物,引物的合成由擎科生物技术(成都)有限公司完成。

表1 qRT-PCR引物Table 1 Primers for quantitative real-time PCR
以cDNA为模板,通过PCR扩增检测反转录产物的特异性和质量。PCR体系为2×Taq Mix 10 μL,上下游引物各 0.8 μL,模板 1 μL,dd H2O 7.4 μL。反应程序为94℃ 2 min;94℃ 30 s,退火 30 s,72℃1 min,35个循环。所得PCR产物进行琼脂糖凝胶电泳检测。
1.6 荧光定量PCR分析
荧光定量PCR反应程序按照TOYOBO公司的THUNDERBIRD SYBR qPCR Mix without ROX试剂盒说明书在CFX Connect型实时定量PCR仪(Bio-Rad,USA)上进行。反应总体积 10 μL,包含THUNDERBIRD SYBR qPCR Mix without ROX 5 μL,10 μmol/L 上下游引物各 0.4 μL,cDNA 模板 1 μL 和无RNA酶水3.2 μL。PCR扩增程序为95℃预变性30 s;95 ℃变性 5 s,退火 30 s,72℃延伸 30 s,40 个循环。基因表达水平采用相对表达量2-ΔΔCt表示。以上所有试验均设3次重复,每次试验设阴性对照。UBQ为内参基因。以‘圆形小酸枣’花后20 d的表达量为对照,其表达量为1。
1.7 数据处理方法
运用Micosoft Excel 2015进行数据处理,并结合SPSS Statistics 17.0统计分析软件。
2 结果与分析
2.1 果实发育过程中可溶性糖组分及含量的动态分析
由枣和酸枣不同发育时期果实可溶性糖含量的变化趋势可知(图 1),‘罗江调元枣’和‘圆形小酸枣’果实发育早期(花后20~48 d)的主要糖组分均为果糖和葡萄糖,占总糖含量的80%以上;在果实发育中后期(花后55~90 d)蔗糖含量迅速上升。花后90 d时‘圆形小酸枣’果实中蔗糖占总糖含量的54.26%,果糖和葡萄糖占总糖含量的45.74%;‘罗江调元枣’果实中蔗糖含量占总糖含量的77.39%,果糖和葡萄糖含量占总糖含量的22.61%。由此可见,‘圆形小酸枣’果实在发育过程中是蔗糖和己糖共同积累,而‘罗江调元枣’果实以积累蔗糖为主。

图1 枣和酸枣果实不同发育时期可溶性糖含量变化Figure 1 Changes of soluble sugar during development in jujube fruits
从糖含量整体变化来看,‘圆形小酸枣’中果糖、葡萄糖和蔗糖随着果实的发育呈现逐渐上升的趋势;而‘罗江调元枣’中果糖和葡萄糖随着果实的发育呈现上升-下降-上升的趋势,蔗糖则在花后55 d开始迅速积累。花后76 d开始,‘罗江调元枣’和‘圆形小酸枣’的蔗糖、果糖和葡萄糖含量出现较大差异。
2.2 果实发育过程中可溶性糖代谢相关基因的表达分析
2.2.1 蔗糖代谢途径中相关基因的表达分析
图 2为‘圆形小酸枣’和‘罗江调元枣’不同发育时期果实蔗糖代谢途径相关基因的表达结果。从图 2A和2C可知,‘圆形小酸枣’中Zj SS1和Zj SS3的表达模式极为相似,花后20~62 d变化不大,花后69~76 d相对表达量急剧上调,花后83 d相对表达量下调,花后90 d相对表达量再次上调;‘罗江调元枣’中Zj SS1和Zj SS3的表达模式较为相似,果实发育前中期相对表达量上调,后期则相对表达量下调。‘圆形小酸枣’和‘罗江调元枣’果实中Zj SS2主要在花后76~90 d有高表达(图 2B)。图 2D~图2F中,‘圆形小酸枣’和‘罗江调元枣’果实中Zj SPS1和Zj SPS2主要在果实发育后期(花后76 d之后)开始高表达;而Zj SPS4的表达趋势为上调-下调-上调-下调,整体呈现“M”型变化。图 2G~图2J中,‘圆形小酸枣’和‘罗江调元枣’果实中Zjc INV3和Zjn INV1在花后20~48 d和花后90 d表达量较高,其他时期则基本不表达。且‘圆形小酸枣’果实中Zjc INV3和Zjn INV1在花后90 d的表达量远高于‘罗江调元枣’;Zjv INV2则在花后34 d有较高表达,其他时期表达量极低。‘圆形小酸枣’中Zjv INV1在花后48~55 d有高表达,其他时期表达量较低或基本不表达,‘罗江调元枣’中Zjv INV1基本不表达。

图2 枣和酸枣果实蔗糖代谢途径中相关基因的表达分析Figure 2 The relative expression level of sucrose metabolism genes during developmental stages of jujube fruit
2.2.2 己糖代谢途径中相关基因的表达分析
图3为‘圆形小酸枣’和‘罗江调元枣’果实不同发育时期己糖代谢途径相关基因的表达量。由图3A和3C可知,‘圆形小酸枣’和‘罗江调元枣’果实中Zj HK1的相对表达量随着果实的成熟,表达逐渐下调;而Zj HK3在果实成熟期相对表达量最高,随着果实的发育,其相对表达量上调-下调-上调,呈现“N”型。图 3B和图3D中,‘圆形小酸枣’果实中Zj HK2和Zj HK5的相对表达量整体呈现下调再上调的变化趋势;‘罗江调元枣’果实中Zj HK2和Zj HK5相对表达量无明显变化规律。‘圆形小酸枣’果实中Zj HK6的相对表达量随着果实的成熟逐渐上调,‘罗江调元枣’果实中Zj HK6的相对表达量在成熟期有高表达(图 3E)。‘圆形小酸枣’果实中UGP的相对表达量随着果实的成熟整体呈上调趋势,而‘罗江调元枣’果实中UGP的相对表达量在花后20~69 d逐渐上调,花后76 d迅速上调后再下调(图 3F)。图 3G和图3H中,‘圆形小酸枣’果实中Zj FK2在成熟期相对表达量较高;‘罗江调元枣’果实中Zj FK2的相对表达量随着果实的成熟略微下调。‘圆形小酸枣’和‘罗江调元枣’果实中Zj FK4的表达则在果实发育前期上调,果实成熟期下调。

图3 枣和酸枣果实己糖代谢途径中相关基因的表达分析Figure 3 The relative expression level of hexose metabolism genes during developmental stages of jujube fruit
2.3 果实发育过程中可溶性糖组分含量与相关基因表达量的相关分析
2.3.1 蔗糖含量与相关基因表达量的相关分析
由蔗糖含量与相关基因表达量的相关分析可知(表2),‘圆形小酸枣’的蔗糖含量与Zj SS1、Zj SS3、Zj SPS1和Zj SPS2的表达量极显著正相关,与Zj SS2和Zjc INV3的表达量显著正相关。‘罗江调元枣’的蔗糖含量与Zj SS2、Zj SPS1和Zj SPS2的表达量极显著正相关,与Zj SS3的表达量显著正相关。
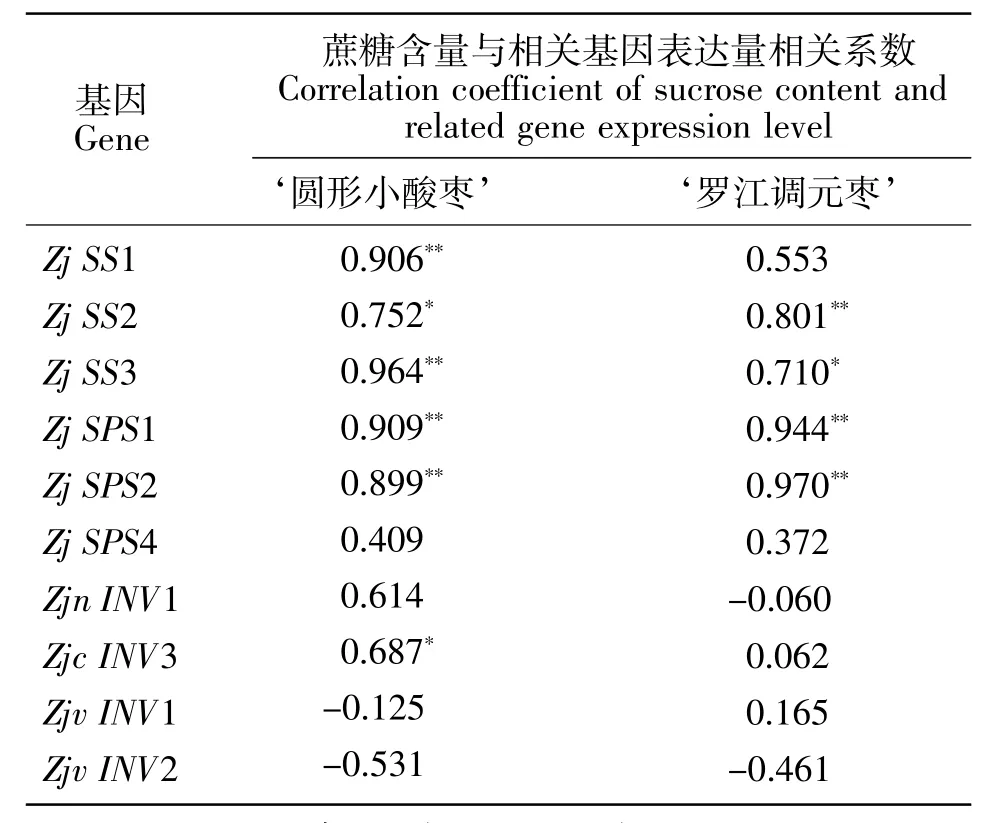
表2 蔗糖含量与相关基因表达量的相关分析Table 2 Correlation analysis between sucrose content and related gene expression level
2.3.2 己糖含量与相关基因表达量的相关分析
由己糖含量与相关基因表达量的相关分析可知(表 3),‘圆形小酸枣’的果糖含量与Zj HK3和Zj HK6的表达量极显著正相关,与Zj FK2、UGP、Zjn INV1和Zjc INV3的表达量显著正相关,与Zj HK1和Zjv INV2的表达量显著负相关。‘圆形小酸枣’的葡萄糖含量与Zj HK3、Zj HK6和Zj FK2的表达量极显著正相关,与Zj HK5、UGP、Zjn INV1和Zjc INV3的表达量显著正相关。而‘罗江调元枣’的果糖和葡萄糖含量仅与Zj FK4的表达量显著正相关。

表3 己糖含量与相关基因表达量的相关分析Table 3 Correlation analysis between hexose content and related gene expression level
3 讨论
3.1 枣和酸枣果实糖组分与含量
蒲小秋[18]检测到‘冬枣’和‘骏枣’果实中以葡萄糖、果糖和蔗糖为主,占总糖含量的99%以上。本试验通过HPLC法检测‘圆形小酸枣’和‘罗江调元枣’果实中的糖组分及含量,发现‘圆形小酸枣’和‘罗江调元枣’果实中的糖组分有蔗糖、果糖和葡萄糖,未检测出山梨糖醇。HPLC试验结果表明‘罗江调元枣’为蔗糖积累型,这与林思思等[10]在‘冬枣’和‘骏枣’上的研究结果一致。‘圆形小酸枣’为蔗糖和己糖共同积累型,而‘延川酸枣A’(灌木型)属于果糖积累型,‘延川酸枣B’(乔木型)属于蔗糖积累型[9]。
3.2 枣和酸枣果实糖代谢基因对糖组分的调控
糖代谢是一个循环的过程,果糖在蔗糖合酶(SS)的作用下可以合成蔗糖,而葡萄糖在己糖激酶(HK)、尿苷二磷酸葡萄糖焦磷酸化酶(UGP)和蔗糖磷酸合酶(SPS)等酶的作用下也可以合成蔗糖,但是蔗糖在转化酶(INV)的作用下又能降解成果糖和葡萄糖[19]。
本研究分析了可溶性糖组分、含量与糖代谢相关基因在‘圆形小酸枣’和‘罗江调元枣’不同发育阶段的表达,发现‘圆形小酸枣’蔗糖含量的变化与Zj SS1、Zj SS2、Zj SS3、Zj SPS1 和 Zj SPS2 表达量变化相似,这与Zhu X.D.等[20]在葡萄中得到的研究结果相似。且‘圆形小酸枣’蔗糖含量与ZjSS1、ZjSS2、ZjSS3、Zj SPS1和Zj SPS2存在显著或极显著相关关系,表明 Zj SS1、Zj SS2、Zj SS3、Zj SPS1 和 Zj SPS2 是‘圆形小酸枣’蔗糖代谢的关键基因。‘罗江调元枣’果实中蔗糖含量的变化与Zj SS2、Zj SS3、Zj SPS1和Zj SPS2表达量变化相似。Ji Y.等[21]利用高粱杂交实验得出SPS不仅是蔗糖积累的关键因素,也是引起甜高粱与普通高粱含糖量差异的因素的结论;且‘罗江调元枣’蔗糖含量与Zj SS2、Zj SS3、Zj SPS1 和Zj SPS2 存在显著或极显著相关关系,表明Zj SS2、Zj SS3、Zj SPS1和Zj SPS2是‘罗江调元枣’蔗糖代谢的关键基因。
‘圆形小酸枣’果实中果糖和葡萄糖含量的变化与Zjv INV1、Zjc INV3和Zjn INV1的表达量变化相反。张玲等[22]的研究结果显示草莓果实发育过程中蔗糖积累与转化酶呈显著负相关,认为随着果实发育,Fa.A/N-Invs编码蛋白通过调控蔗糖分解,影响果实糖分积累,这与本试验结果相似。‘罗江调元枣’果糖和葡萄糖含量与INV表达量变化无明显相关关系。且相关分析表明,‘圆形小酸枣’果糖与葡萄糖含量变化与Zjc INV3和Zjn INV1的表达量显著相关,‘罗江调元枣’果糖与葡萄糖含量变化与任一INV的表达量不存在显著相关关系。蒲小秋[18]认为枣果实中酸性转化酶基因对调控己糖平衡具有重要作用。郭雪飞等[23]对还原糖积累型枣果实‘金铃圆枣’的研究表明,果实发育中后期中性转化酶(IN)活性与己糖含量显著正相关。因此,依据本试验的结果,可以推测Zjv INV1、Zjc INV3和Zjn INV1是‘圆形小酸枣’果糖和葡萄糖合成的关键调节基因。
花后76 d开始,‘圆形小酸枣’和‘罗江调元枣’的蔗糖含量开始出现较大差异,到花后90 d‘罗江调元枣’的蔗糖含量比‘圆形小酸枣’高1/2,然而‘圆形小酸枣’许多发育时期的Zj SS2、Zj SS3、Zj SPS1和Zj SPS2表达量却高于‘罗江调元枣’。这可能是叶片通过光合作用产生大量蔗糖,蔗糖通过长距离运输从源器官(叶片)到达库器官(果实)后造成的差异[24],这方面的推测还有待进一步研究。花后76 d开始,‘圆形小酸枣’和‘罗江调元枣’果糖和葡萄糖含量开始出现较大差距,到花后90 d‘圆形小酸枣’的果糖和葡萄糖含量比‘罗江调元枣’高1/2,这是由于在此期间‘圆形小酸枣’中Zjv INV1、Zjc INV3和Zjn INV1的表达量远高于‘罗江调元枣’所致。
4 结论
利用HPLC法测定了枣和酸枣果实中的糖组分与含量,发现本试验所用‘圆形小酸枣’为蔗糖和己糖共同积累型,‘罗江调元枣’为蔗糖积累型。通过实时荧光定量结果发现,Zj SS1、Zj SS2、Zj SS3、Zj SPS1和Zj SPS2是‘圆形小酸枣’蔗糖合成的关键基因;Zj SS2、Zj SS3、Zj SPS1 和 Zj SPS2 是‘罗江调元枣’蔗糖合成的关键基因;Zjv INV1、Zjc INV3和Zjn INV1是‘圆形小酸枣’果糖和葡萄糖合成的关键基因。‘圆形小酸枣’和‘罗江调元枣’糖积累规律的差异主要从花后76 d开始,两者己糖的积累规律差异主要由Zjv INV1、Zjc INV3和Zjn INV1的差异表达所致,蔗糖的积累规律差异还有待进一步研究。
猜你喜欢
中国果业信息(2022年1期)2022-11-28
作物学报(2022年1期)2022-11-05
果农之友(2022年9期)2022-09-27
今日农业(2022年13期)2022-09-15
当代县域经济(2022年8期)2022-08-01
当代县域经济(2022年7期)2022-07-15
农业工程学报(2022年7期)2022-07-09
三悦文摘·教育学刊(2022年7期)2022-04-27
都市(2022年1期)2022-03-08
小学生导刊(2018年20期)2018-11-30
