
高寒草甸不同扰动斑块植物功能群和根土复合体特征变化研究
2022-03-04 12:33宋梓涵李希来李杰霞卡着才让马戈亮
生态科学 2022年1期
宋梓涵, 李希来,*, 李杰霞, 卡着才让, 马戈亮
高寒草甸不同扰动斑块植物功能群和根土复合体特征变化研究
宋梓涵1, 李希来1,*, 李杰霞1, 卡着才让2, 马戈亮2
1. 青海大学农牧学院, 西宁 810016 2. 青海省河南蒙古族自治县草原综合专业队, 河南县 811500
高原鼠兔()活动在高寒草甸上会形成各种类型扰动斑块, 扰动斑块类型的转变是导致草地退化的原因之一。目前, 斑块类型发生转变的关键影响因子尚不清楚, 因此, 对高寒草甸活动斑块、非活动斑块和恢复斑块三个扰动斑块类型植物功能类群结构、根土复合体特征和斑块分布特征进行研究, 旨在探讨斑块类型发生转变的主要因子, 说明高寒草甸退化机理。研究结果表明, 高寒草甸中活动斑块土壤表层由于放牧和高原鼠兔活动扰动强度大, 无法形成稳定的植物群落, 非活动斑块的植物群落处于次生演替早期阶段, 但是恢复斑块已具备高寒草甸优势种莎草科植物生长的基本条件。退化高寒草甸中活动斑块与非活动斑块呈现镶嵌性分布, 而恢复斑块相对独立没有镶嵌性分布现象。造成高寒草甸干扰斑块类型转变的主要原因是高原鼠兔活动强度由间歇性干扰转变为持续干扰的结果。
高寒草甸; 干扰斑块; 植物功能群; 根-土复合体
0 前言
高原鼠兔在高寒草甸上挖掘、觅食等活动会形成不同类型的干扰斑块[1], 随着这些干扰斑块的不断扩大连通, 将导致高寒草甸退化形成“黑土滩”[2]。高原鼠兔活动形成的干扰斑块在高寒草甸生态系统内起着生态调节作用还是破坏作用, 判断的主要依据是高原鼠兔活动强度是否影响了草地植物群落的稳定性[3]。以往对高原鼠兔干扰的研究主要集中于草地生态系统植物多样性和生物量的变化上[4–5], 只说明了高原鼠兔活动干扰造成的草地退化结果, 没有深入探讨干扰斑块的特征变化, 有关干扰斑块类型与其演替机理的研究非常薄弱[6]。
根据顶级群落理论将高寒草甸各类干扰斑块划分为从次生裸地到斑块不同恢复阶段[5]。尚占环认为干扰斑块是高寒草甸退化的窗口, 干扰斑块是由于长期干扰无法恢复, 其面积逐年增加的结果[2]。干扰斑块的演替以退化演替和恢复演替两种形式存在于高寒草甸上。为了研究高寒草甸在不同类型干扰斑块影响下的变化, 我们将高寒草甸中的各种类型扰动斑块划分为活动斑块、非活动斑块和恢复斑块, 通过研究它们之间的关系和动态演替规律揭示高寒草甸的退化规律。高原鼠兔干扰活动制造的活动斑块是研究高寒草甸退化的主要斑块类型。对于恢复演替方面, 主要的研究对象是有明显恢复迹象的斑块。既没有高原鼠兔活动, 也没有恢复迹象的斑块是非活动斑块。高寒草甸高原鼠兔的活动是造成草地局部土壤物理结构破坏的主要原因, 因此通过测定草地土壤的抗剪切强度, 有助于理解干扰斑块类型演替过程与机理[7]。本文采用原位十字剪切法测量不同干扰斑块土壤抗剪切强度[8], 采用无人机遥感影像解译高原鼠兔干扰斑块的分布特征, 旨在通过研究高寒草甸中活动斑块、非活动斑块和恢复斑块植物功能群和根土复合体的特征变化, 探讨高原鼠兔活动对高寒草甸稳定性的影响, 为合理控制高原鼠兔种群密度提供科学参考。
1 材料与方法
1.1 研究区概况
研究地点在青海省河南蒙古族自治县克其合滩, 高寒草甸植物群落主要以矮生嵩草()为建群种。研究样地为退化的高寒草甸, 北纬34°41′7′′, 东经101°46′2′′, 海拔3743 m, 阳坡滩地, 坡度3°。2018年6月通过连续三天的堵洞法测定样地的有效洞口数, 有效洞口密度为300个·hm-2。
1.2 试验设计
高原鼠兔干扰斑块按照不同的土壤和植被特征进行分类, 分为活动斑块、非活动斑块和恢复斑块等3类斑块(见图1), 未有干扰斑块的高寒草甸作为对照。每种类型斑块在试验样地上随机选取5个作为重复。活动斑块是高原鼠兔活动区, 土壤裸露疏松、偶有植物生长的斑块; 非活动斑块是由高原鼠兔停止扰动活动而形成的斑块, 具有少量杂类草植物, 优势物种主要是密花香薷()、四数獐牙菜()、西藏微孔草()和细叶亚菊(); 恢复斑块是没有高原鼠兔活动正在进行自然生态恢复的斑块, 主要以莎草科植物黑褐苔草()为优势物种, 植被覆盖率小于60%; 对照高寒草甸是以矮生嵩草为建群种的草甸。
1.3 采样方法
取样时间为2019年6月, 在选取的每个类型斑块上设置1个50 cmÍ50 cm的样方, 重复5次。在样方内, 按照植物功能群划分为禾本科植物、莎草科植物和杂草类进行群落特征测定。随机选择不同植物功能群10株植物测量植物高度, 使用目测法测量每种植物盖度, 在斑块内随机抛样圆10次测量植物频度[9]。根据植物生活史将杂类草划分为一、二年生杂类草(密花香薷、四数獐牙菜和西藏微孔草等)和多年生杂类草(细叶亚菊等)[10]。
在做样方的位置上建立标记, 使用不同颜色标记不同种类的斑块。降雨结束后, 用三参数速测仪(型号: TDR350)测量土壤表面0—7 cm的土壤湿度(体积含水量)。用直径60 mm的根钻取15 cm土样, 每个斑块重复3次, 用纱布清洗法测量地下生物量。
利用自制的十字板静力探测仪测量根-土复合体的剪切强度[8]。土壤剪切强度是常用的土壤力学指标, 通过测量力学指标可以了解高原鼠兔的干扰方式。原状土剪切强度是未经扰动的土壤抵抗剪切力破坏的极限应力值, 重塑土剪切强度是完全被破坏的土壤抵抗剪切力造成位移的极限应力值。由于高原鼠兔洞口附近土壤不稳定, 因此探测点设置在距离洞口20 cm以外的地方。测量方法是: 将十字板打入测量斑块土壤表层15 cm, 静置2分钟; 扭转十字板, 转速为1—2°每10 s, 在2分钟内得到原状土剪切强度最大值; 十字板沿剪切方向旋转1圈360度, 充分破坏周围土体; 测量重塑土剪切强度, 转速1—2°每10 s, 当剪切曲线开始下降或稳定不变时, 获得重塑土剪切强度[8]。

图1 3种类型的干扰斑块及高寒草甸
Figure 1 Three types of interfering patch and alpine meadow
使用大疆PHANTOM4无人机在研究地点拍摄低空正射影像。为了获得分辨率为1 cm的照片, 飞行高度设置为39 m[11]。使用皮尺在研究地点标记10 m单位长度, 建立比例尺。拼接无人机影像, 获得整个研究区的影像。在无人机影像上根据样方上的标记识别出各类斑块, 使用Arcgis10.4软件获得斑块面积及鼠兔洞口等信息。
1.4 数据分析
使用SPSS 22.0软件对不同类型斑块的植物功能群重要值、植物功能群盖度、地下生物量、土壤含水量、剪切强度进行单因素方差分析, 并用Duncan法对各测定数据进行多重比较; 使用Origin 2018软件制图。计算公式如下:
(1)重要值=(相对高度+相对盖度+相对频度)/3
2 结果与分析
2.1 不同类型斑块植物功能群特征变化
如图2所示, 活动斑块的多年生杂类草重要值显著高于其他类型斑块(<0.05)。非活动斑块与恢复斑块的莎草科植物重要值差异显著(<0.01)。研究发现一、二年生杂类草植物仅生长在非活动斑块内, 且其重要值显著高于其他功能群(<0.05)。恢复斑块和高寒草甸的多年生杂类草重要值之间没有显著差异, 莎草科植物重要值也没有显著差异, 这表明恢复斑块已经基本具备了正常高寒草甸的植物群落结构特征。除了非活动斑块具有少量禾本科植物外, 活动斑块不存在禾本科植物, 说明自然生态恢复下, 禾本科植物的生长是需要稳定的表层土壤理化条件。
如图3所示, 活动斑块上仅有极少数先锋植物生长。尽管非活动斑块和恢复斑块的植物总盖度相似, 但是非活动斑块上仍然以一、二年生杂类草植物为主, 而恢复斑块中莎草科植物已经占主导地位。图4是不同斑块类型的地下生物量变化。高寒草甸地下生物量显著高于其他类型斑块(<0.01)。活动斑块上几乎没有植物生存, 其地下生物量与非活动斑块和恢复斑块的没有显著差异。活动斑块的地下生物量主要由干扰前的植物组成。
2.2 不同类型斑块的根-土复合体特征变化
如图5所示, 恢复斑块、非活动斑块和对照高寒草甸的表层土壤含水量没有显著差异, 活动斑块显著低于其他类型斑块(<0.05)。因此, 水分不是导致恢复斑块与非活动斑块的植物群落出现差异的主要原因, 莎草科植物不会因为土壤含水量低而无法在这些次生裸地上生长。

注: 不同小写字母表示同类斑块内不同功能群间重要值差异显著(P<0.05), 不同大写字母表示不同斑块间相同植物功能群重要值差异显著(P<0.05)。
Figure 2 Important values of plant functional groups of different types of patch

图3 不同类型斑块的植物功能群盖度
Figure 3 Coverage of plant functional groups of different types of patch

图4 不同类型斑块的地下生物量
Figure 4 Underground biomass of different types of patch

图5 不同类型斑块土壤含水量
Figure 5 Water content of soil of different types of patch
图6显示了不同类型斑块的土壤剪切力强度。恢复斑块的原状土剪切力强度大于其他干扰斑块。高寒草甸的原状土剪切力强度达到60 kPa, 说明放牧活动等干扰很难对高寒草甸造成干扰影响。如图6所示, 恢复斑块和非活动斑块的重塑土剪切强度与活动斑块的原状土剪切强度没有差异, 它们显著低于原生草甸重塑土剪切强度(<0.05)。
2.3 高寒草甸不同类型斑块分布特征变化
从无人机影像上获得的斑块分布特征如表1所示。样地内非活动斑块面积远大于活动斑块和恢复斑块面积。选取相对较小面积的非活动斑块, 研究非活动斑块与恢复斑块的差异。非活动斑块和活动斑块内有较多的高原鼠兔洞口, 而恢复斑块上没有鼠兔洞口或仅有一个鼠兔洞口。
干扰斑块在高寒草甸的分布如图7所示, 图中被标记的部分是本文测定的斑块。面积较大的干扰斑块内分布着活动斑块和非活动斑块。活动斑块在图像上呈白色的原因是活动斑块上没有植被覆盖,且土壤湿度较低。活动斑块上可以看到高原鼠兔活动的痕迹, 植物无法正常生长。非活动斑块由于植物颜色较深而呈现褐色。非活动斑块土壤表层结构开始趋于稳定, 没有发现高原鼠兔活动痕迹, 并伴有直根系杂类草的生长。恢复斑块颜色呈浅绿色, 其位置独立于其他斑块, 其分布与活动斑块和非活动斑块没有关系。根据上述各类干扰斑块的颜色解译无人机图像, 获得各类斑块占草地面积的比例, 结果表明样地中干扰斑块占草地总面积的30%, 其中活动斑块占总面积的10%, 非活动斑块占总面积的15%, 恢复斑块占总面积的5%。活动斑块与非活动斑块呈相互镶嵌的结构, 活动斑块占干扰斑块40%的面积。

表1 不同种类斑块的无人机影像特征

图6 不同类型斑块的剪切强度
Figure 6 Shear strength of different types of patch

图7 不同类型干扰斑块在高寒草甸上的分布
Figure 7 Distribution of different types of patch in alpine meadow
3 讨论
3.1 不同类型斑块植物功能群重要值的差异
非活动斑块中植物功能群重要值最高的是一、二年生植物, 这说明一、二年生植物种群能忍受较频繁的扰动干扰。恢复斑块中植物功能群重要值最高的是莎草科植物, 这说明莎草科植物在植物群落竞争中具有较强优势。江小雷等(2004)和刘伟等(2012)通过对高原鼢鼠和高原鼠兔土丘的植物群落分析研究, 认为一、二年生植物逐渐会被莎草科植物替代, 非活动斑块会演替为恢复斑块[12,14]。非活动斑块的植物群落结构与退化斑块和黑土滩相似[2]。干扰斑块在演替过程中受到频繁的扰动, 会导致高寒草甸的恢复演替无法进行。
3.2 不同类型斑块的根-土复合体特征与高原鼠兔干扰
高寒草甸土壤质地以重砂壤土为主, 土壤抗剪强度很低。根-土复合体抗剪强度的提高主要依靠根系的作用[15–16]。在相同的根系结构组成的条件下, 根-土复合体的力学特征相同[17]。恢复斑块和非活动斑块的重塑土剪切强度与干扰斑块的原状土剪切强度没有差异, 且这三种斑块的地下生物量没有差异, 这表明被破坏的恢复斑块和非活动斑块的根系结构与活动斑块的相同。因此, 很多非活动斑块由于未开始恢复演替而再次受到高原鼠兔活动的干扰, 重新演替为活动斑块。这种现象主要与高原鼠兔的高强度活动有关, 因此, 适当控制鼠兔种群密度可以抑制高寒草甸的进一步退化。
3.3 不同类型斑块的演替特征
根据不同类型斑块的植物功能群和根-土复合体特征可以总结出如图8的干扰斑块演替过程。一方面, 图6所示的土壤剪切强度研究表明, 活动斑块是由于非活动斑块受外力翻动土壤表层而形成的。活动斑块地下生物量与非活动斑块的没有显著差异也能证明这一点。从无人机遥感图像上可看出, 较大面积的干扰斑块由活动斑块和非活动斑块两部分组成。以上结果说明这两种斑块的群落可以互相转化, 从而形成了两类斑块之间的演替过程[18]。随着高原鼠兔种群密度的增加, 洞穴分布由随机分布转变为聚集分布也证明了这种干扰模式的存在[19]。活动斑块作为干扰斑块恢复演替过程的负反馈机制使群落处于稳定的低生产力水平状态, 无法恢复到高生产力水平的高寒草甸[20]。
另一方面, 恢复斑块的植物功能群结构与高寒草甸相似, 平均面积较小, 其中的鼠兔洞口逐渐消失。恢复斑块的植物群落与形成两年的高原鼠兔土丘的植物群落相同, 说明恢复斑块中至少两年没有高原鼠兔的活动[14]。由于干扰斑块的草毡层的破坏, 高寒草甸土壤的物理和化学性质发生了变化[21–22]。草毡层隔热能力的丧失导致了冻土层退化和破坏[23]。这些因素会对非活动斑块内的植物和土壤构成干扰, 抑制非活动斑块群落的正向演替。由于土壤养分和土壤种子库的减少, 即使高原鼠兔停止干扰, 非活动斑块仍需要3至4年时间才能过渡到恢复斑块演替阶段[24]。恢复斑块上的优势物种黑褐苔草虽然是莎草科植物, 但不是高寒草甸的优势建群种, 也没有发达的根系形成草毡层, 以营养繁殖为主的矮生嵩草和高山嵩草()需要稳定的生态环境才能成为恢复斑块建群种[13]。图8所示的从活动斑块演替为恢复斑块的时间与控制高原鼠兔种群密度的周期相同[25]。这表明减轻高原鼠兔的干扰活动能促使退化高寒草甸土壤和植被的逐步恢复。
在图8中, 长时间未受干扰的斑块能从次生裸地的活动斑块恢复到顶级群落的高寒草甸。在这种干扰模式下, 高原鼠兔两次入侵同一处草地的间隔时间比干扰斑块恢复为高寒草甸的时间长, 因此将这种干扰模式称为间歇性干扰模式。图8的活动斑块与非活动斑块相互演替过程构成了干扰斑块的持续性干扰模式, 这是高原鼠兔密度增大所产生的两次入侵间隔时间小于干扰斑块恢复时间造成的, 为防止高寒草甸的进一步退化, 及时控制高原鼠兔密度是非常有必要的。持续干扰模式使活动斑块和非活动斑块无法通过恢复过程而减少, 面积逐渐持续扩大, 并连通其他斑块, 最终形成黑土滩退化草地[26]。高原鼠兔干扰由间歇性干扰模式转变为持续性干扰模式, 是导致高寒草甸退化斑块扩大连通的原因, 也是退化黑土滩草地形成的主要原因。
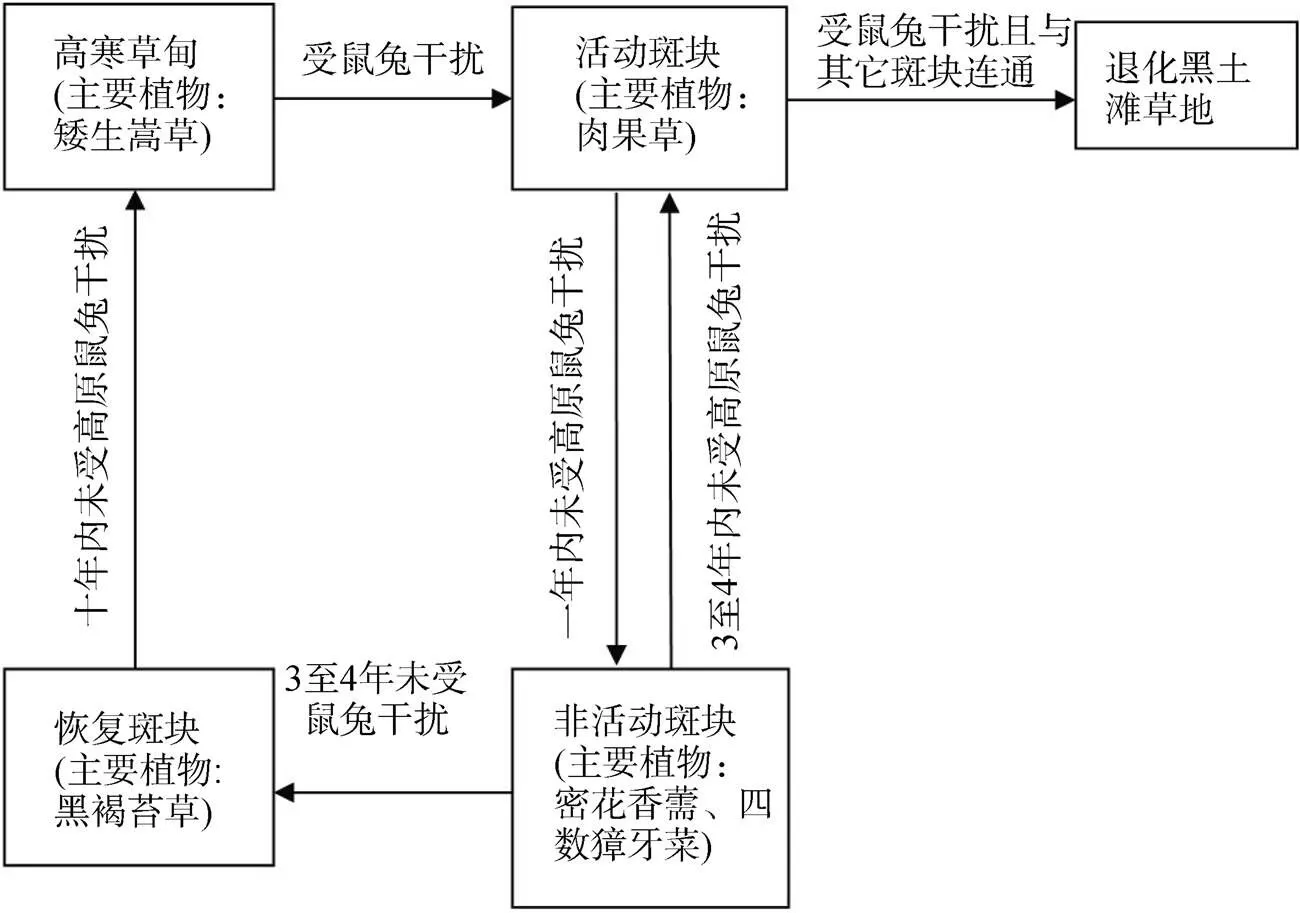
图8 干扰斑块演替过程
Figure 8 Succession process of interfering patch
4 结论
高寒草甸由于高原鼠兔的活动会形成不同类型的干扰斑块, 这些干扰斑块主要包括活动斑块、非活动斑块和恢复斑块。研究发现活动斑块上偶有植物生长, 非活动斑块以一、二年生植物为主, 恢复斑块的优势种植物是黑褐苔草。非活动斑块和恢复斑块的重塑土剪切强度与活动斑块的原状土剪切强度没有显著差异, 三种斑块的地下生物量没有显著差异。活动斑块与非活动斑块呈现镶嵌性分布, 恢复斑块的分布相对独立。由于高原鼠兔密度增加, 鼠兔干扰由间歇性干扰转变为持续性干扰模式, 是导致高寒草甸退化斑块扩大连通的原因, 也是退化草地黑土滩形成的主要原因。
[1] BRIGITTE W H. The plateau pika: A Keystone Engineer on the Tibetan plateau[D]. Phoenix: Arizona State University, 2010.
[2] 尚占环, 龙瑞军. 青藏高原“黑土型”退化草地成因与恢复[J]. 生态学杂志, 2005, 24(6): 652–656.
[3] ANTHONY D A, ROGER P P, JIEBU, et al. Grassland degradation on the Tibetan plateau: the role of small mammals and methods of control[M]. Canberra: Australian Centre for International Agricultural Research, 2007.
[4] LIU Yanshu, FAN Jiangwei, SHI Zhongjie, et al. Relationships between plateau pika () densities and biomass and biodiversity indices of alpine meadow steppe on the Qinghai–Tibet Plateau China[J]. Ecological Engineering, 2017, 102: 509–518.
[5] 许华磊, 王溪, 刘伟. 放牧条件下高原鼠兔扰动对高寒草甸植物群落特征的影响[J]. 兽类学报, 2019, 39(05): 537–545.
[6] PANG Xiaopan, GUO Zhenggang. Plateau pika disturbances alter plant productivity and soil nutrients in alpine meadows of the Qinghai-Tibetan Plateau, China[J]. The Rangeland Journal, 2017, 39: 133–144.
[7] 周云艳, 陈建平, 王晓梅. 植物须根固土护坡的复合材料理论研究[J]. 武汉理工大学学报, 2010, 32(18): 103–107.
[8] 宋路, 朱海丽, 李国荣, 等. 两种方法测定根-土复合体抗剪强度试验对比研究[J]. 水土保持研究, 2016, 23(04): 282–287.
[9] 施颖, 胡廷花, 高红娟, 等. 两种放牧模式下高寒草甸群落植被构成及稳定性特征[J]. 草业学报, 2019, 28(09): 1–10.
[10] ZHOU Bingrong, LI Shuai, LI Fu, et al. Plant functional groups asynchrony keep the community biomass stability along with the climate change- a 20-year experimental observation of alpine meadow in eastern Qinghai-Tibet Plateau[J]. Agriculture, Ecosystems and Environment. 2019, 282: 49–57.
[11] SUN D, ZHENG J H, MA T, et al. The analysis of burrows recognition accuracy in Xinjiang’s pasture area based on UAV visible images with different spatial resolution[C]// ISPRS TC III Mid-term Symposium “Developments, Technologies and Applications in Remote Sensing”. Beijing, 2018: 1575–1579.
[12] 江小雷, 张卫国, 杨振宇, 等. 不同演替阶段鼢鼠土丘群落植物多样性变化研究[J]. 应用生态学报, 2004, 15(5): 814–818.
[13] 李积兰, 李希来, 马福俊. 黄河上游高寒草甸矮嵩草生理特征对流域不同生境的响应[J]. 草业学报, 2018, 27(01): 62–72.
[14] 刘伟, 李里, 严红宇, 等. 高原鼠兔挖掘活动对植物种的丰富度和地上生物量的影响[J]. 兽类学报, 2012, 32(03): 216–220.
[15] 程洪, 颜传盛, 李建庆, 等. 草本植物根系网的固土机制模式与力学试验研究[J]. 水土保持研究, 2006, (01): 62–65.
[16] 格日乐, 张成福, 蒙仲举. 3种植物根-土复合体抗剪特性对比分析[J]. 水土保持学报, 2014, 28(2): 85–90.
[17] 言志信, 宋云, 江平. 植被护坡中植物根和岩土相互作用的力学分析[J]. 应用数学和力学, 2010, 31(5): 585–590.
[18] MAY R M. Patterns of species abundance and diversity[C] // CODDY M L, DIAMOND J M. Ecology and Evolution of Communities. Cambridge: Harvard University Press, 1975: 81–120.
[19] 张瑞, 许华磊, 刘伟. 高原鼠兔种群恢复过程中的洞穴分布动态[J]. 兽类学报, 2018, 38(1): 46–55.
[20] 马玉寿, 张自和, 董全民, 等. 恢复生态学在“黑土型”退化草地植被改建中的应用[J]. 甘肃农业大学学报, 2007, 42(2): 91–97.
[21] 张雯娜, 金少红, 于成, 等. 高原鼠兔洞口密度对高山嵩草草甸土壤主要养分含量的影响[J]. 草业科学, 2018, 35(07): 1593–1601.
[22] 姚宝辉, 王缠, 张倩, 等. 甘南高寒草甸退化过程中土壤理化性质和微生物数量动态变化[J]. 水土保持学报, 2019, 33(3): 138–145.
[23] 胡宏昌, 王根绪, 王一博, 等. 江河源区典型多年冻土和季节冻土区水热过程对植被盖度的响应[J]. 科学通报, 2009, 54(2): 242–250.
[24]才文代吉, 张静, 谈静, 等. 高寒草甸鼠丘土壤种子库结构特征的研究[J]. 草地学报, 2019, 27(5): 1181–1187
[25] 卫万荣. 高原鼠兔和高原鼢鼠种群消长规律及其与植被关系的研究[D]. 兰州: 兰州大学, 2018.
[26] LI Xilai. The spatio-temporal dynamics of four plant-functional types (PFTs) in alpine meadow as affected by human disturbance, Sanjiangyuan region, China[D]. Auckland: The University of Auckland, 2012.
Characteristics of plant functional groups and root-soil complexes of different disturbed patches in alpine meadow
SONG Zihan1, LI Xilai1,*, LI Jiexia1, Kazhuocairang2, MA Geliang2
1. College of Agriculture and Animal Husbandry, Qinghai University, Xining 810016, China 2. Grassland Integrated professional team of Mongolian Autonomous County of Henan, Henan 811500, China
Activity of plateau pika () disturbs alpine meadows and creates disturbed patches. The type change of patches leads to the change of ecosystem resilience. Currently, it is unclear which key indicators of disturbed characteristics lead to a shift in patch type. We classified the disturbed patch into three categories (active patch, restored patch and inactive patch), and investigated the community functional groups structure, soil-root complex characteristics and patch distribution characteristics of different types of patches. The results revealed that with the disturbance of grazing and plateau pika in the grassland, the soil of active patches was strongly disturbed and it was impossible to form a stable plant community. Plant community of inactive patches was in the early stages of secondary succession. Restored patches already had conditions for the growth of sedge plants. Active patches and inactive patches were inlaid with each other, and they could be transformed into each other. Restored patches did not intersect with the above two types of patch. Our research indicates that the transform of disturbing behaviors of plateau pika from intermittent interference to continuous and interference is the main cause of type changes of disturbed patches.
alpine meadow; disturbed patch; plant functional group; root-soil complex
10.14108/j.cnki.1008-8873.2022.01.004
宋梓涵, 李希来, 李杰霞,等. 高寒草甸不同扰动斑块植物功能群和根土复合体特征变化研究[J]. 生态科学, 2022, 41(1): 31–38.
SONG Zihan, LI Xilai, LI Jiexia, et al. Characteristics of plant functional groups and root-soil complexes of different disturbed patches in alpine meadow[J]. Ecological Science, 2022, 41(1): 31–38.
S812.6
A
1008-8873(2022)01-031-08
2020-06-02;
2020-07-07
国家自然科学基金面上项目(31872999); 青海省科学技术厅项目(2018-ZJ-781); 高等学校学科创新引智计划(D18013); 青海省科技创新创业团队项目“三江源生态演变与管理创新团队”
宋梓涵(1994—), 男, 青海省西宁人, 硕士研究生, 研究方向为草地生态与环境保护, E-mail: 2521980537@qq.com
李希来(1964—), 男, 博士, 教授, 主要从事高原生态修复与环境模拟研究, E-mail: xilai-li@163.com
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
海外文摘·文学版(2021年3期)2021-05-19
矿产勘查(2020年11期)2020-12-25
航空发动机(2020年3期)2020-07-24
家教世界·创新阅读(2020年4期)2020-06-03
当代工人·精品C(2020年1期)2020-05-20
家教世界·创新阅读(2020年1期)2020-05-11
家教世界·创新阅读(2020年2期)2020-04-07
模具制造(2019年10期)2020-01-06
