
不同耐低温辣椒花芽分化及形态生理特性研究
2022-03-05 01:12张彦君邓昌蓉侯智超邵登魁
青海农林科技 2022年1期
张彦君,邓昌蓉,李 江,陈 艳,侯智超,邵登魁*
(1.青海大学农林科学院园艺研究所,青海 西宁 810016;2.青海省蔬菜遗传与生理重点实验室,青海 西宁 810016)
辣椒(CapsicumAnnuumL.)属茄科(Solanaceae)辣椒属(Capsicum)一年或多年生喜温性草本植物。青海高原地区受海拔和纬度影响,绝对低温和年积温均不太适宜辣椒露地栽培,因此多数辣椒在设施中栽培,即使在设施条件下,也经常会出现冬春季冷害或冻害,造成大量落花落果[1-3]。在生产中,如苗期遭受低温,先期辣椒成花率会明显降低。明确辣椒花芽分化显微学特性,系统开展辣椒花芽分化过程及形态特性研究是开展不同耐低温性材料花芽分化遗传特性及相关信号调控等机理研究的先决条件。
前人已经在许多植物进行了花芽分化形态的研究,如苹果[4]、枣花[5]、白菜[6]、棉花[7]、日本落叶松[8]、花生[9]、杨梅[10]、鸭梨[11]、油茶[12]、西瓜[13]、扁桃[14]、李[15]、大花铁线莲[16]、大葱[17]等。对花芽分化的显微学研究大多采用石蜡切片和电镜扫描的方法,一般将花芽分化过程大致分为4-6个时期。茄科作物花芽分化的有研究报道的仅有番茄[18]、马铃薯[19]等,关于不同耐低温材料的相关研究报道较少。本试验选择不同耐低温性辣椒品种为材料,采用石蜡切片法,对辣椒花芽的形态分化进行研究,旨在为深入开展辣椒花芽分化机理及材料间差异性研究提供理论依据。
1 材料和方法
1.1 材料
试验选用2个不同耐低温性辣椒材料,强耐低温品种华美105,弱耐低温品种乐都长辣椒,种子均来源于青海省农林科学院园艺研究所。
1.2 方法
2021年2月25日将2个辣椒品种播种于青海省农林科学院1号试验园育苗棚,4月18日筛选大小一致的壮苗,分苗于50穴的穴盘中进行后续的试验检测,田间正常管理。花芽采样时期为4月23日-6月4日。材料每隔2d,于每日9时-10时进行顶部门椒花芽部位的采集,采集后用FAA固定液固定并保存,常规石蜡法制片。并对同时期辣椒幼苗株高、完全功能叶、叶绿素含量和可溶性糖含量进行测定。
2 结果与分析
2.1 不同耐低温辣椒花芽分化过程比较
通过生长锥形态变化特点(图1,图2),将2个辣椒材料花芽分化分为5个时期。 花芽分化初期:辣椒叶芽处生长锥变圆凸起明显,长度大于宽度(图1a)。萼片分化期:生长锥顶部变宽,在顶部两边出现2个向内弯曲的小突起(萼片原基)将生长锥包在中间(图1b)。花瓣分化期:萼片分化后期,生长锥顶部凹陷,凹陷两边凸起为花瓣原基,此时萼片原基向内弯曲外侧连在一起,形成两个三角锥突起(图1c)。雄蕊分化期:随着花芽不断分化,雄蕊原基分化完成,在花瓣原基内侧突起(图1d)。雌蕊分化期:同雄蕊分化期一样,雌蕊原基在雄蕊原基基部形成突起(图1e)。雌蕊原基发育生长后呈锥形(图1f)。之后随着胚珠的产生,最终形成完整花器官(图1g)。
对2个辣椒材料花芽分化时期进行对比发现,从乐都长辣椒花瓣分化期(图1c)和华美105花瓣分化期(图2c)可以发现,乐都长辣椒萼片比华美105凹陷更加明显;通过比较图1c和图2c可以看出,乐都长辣椒花柄附着点较细,而华美105花柄附着点较粗;从图1f和图2f可以看出,乐都长辣椒心皮分化完成后空腔更小,且较华美105心皮分化完成后空腔较大,形成的三角锥形更钝。
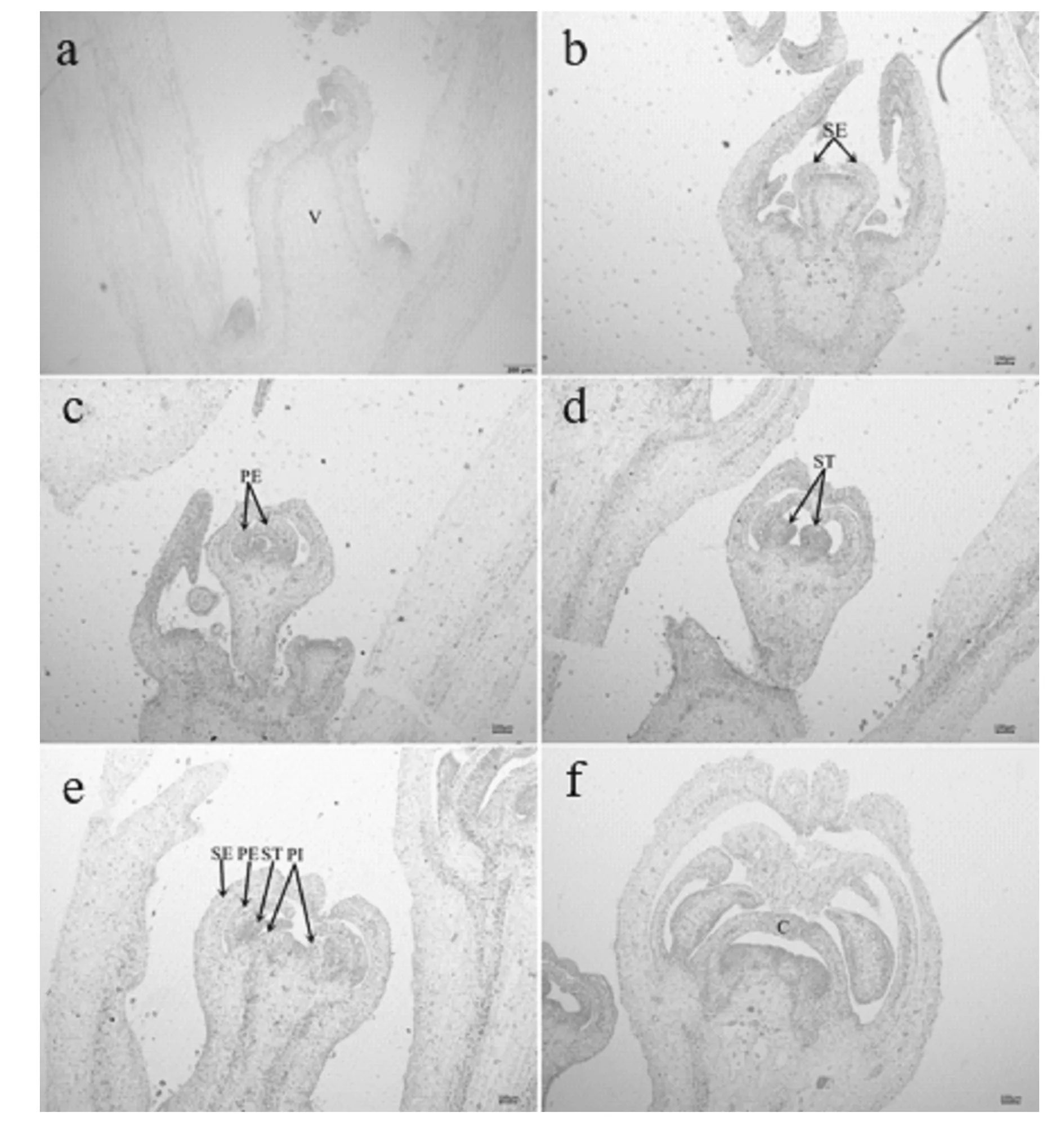
图1 乐都长辣椒花芽分化过程Fig.1 The courses of flower bud differentiation of ledu long pepper注:a:分化初期;b:萼片分化;c:花瓣分化;d:雄蕊分化;e:雌蕊分化;f:心皮;g:胚珠。V:生长锥;SE:萼片原基;PE:花瓣原基;ST:雄蕊原基;PI:雌蕊原基;C:心皮。(放大倍数:×40),下同。
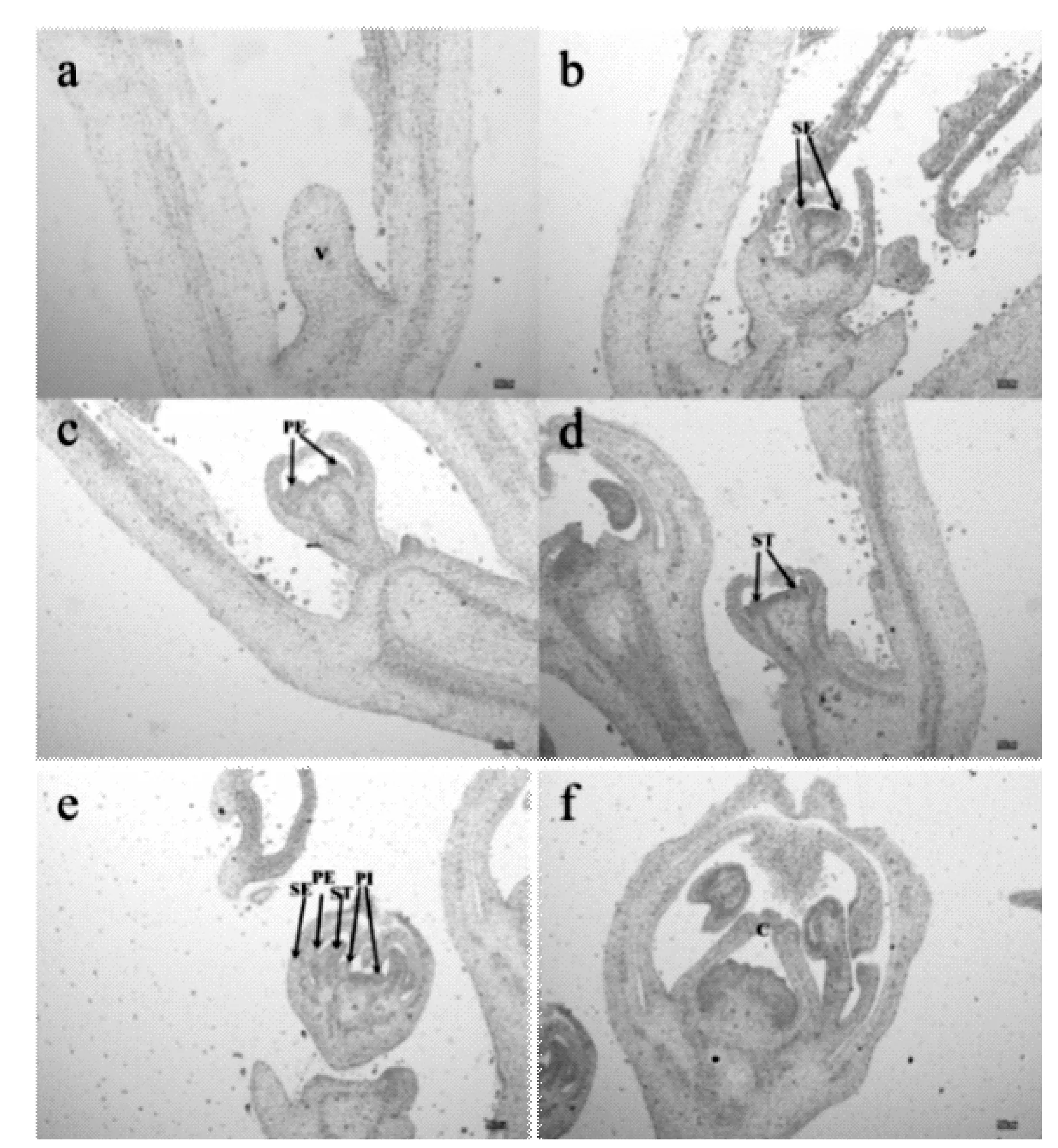
图2 华美105花芽分化过程Fig.2 The courses of flower bud differentiation of Huamei 105 pepper
2.2 不同耐低温辣椒花芽分化期株高的变化
由图3可知,乐都长辣椒在株高 8cm左右时进入门椒花芽分化初期,10-11cm进入萼片分化期,11-12cm时进入花瓣分化期,13cm时进入雄蕊分化期,14cm时进入雌蕊分化期,之后开始心皮分化,在株高达到16cm时心皮分化完成,之后随着胚珠的产生,门椒生长锥顶部花芽最终形成一个成熟的花器官。华美105则在株高9cm左右时开始门椒花芽分化;门椒花芽分化完成时,乐都长辣椒的株高在16cm左右,而华美105的株高在18cm左右。华美105株高在花芽分化各时期株高均高于乐都长辣椒。

图3 辣椒花芽分化过程中株高的变化Fig.3 Changes of plant height of pepper duringflower bud differentiation
2.3 不同耐低温辣椒花芽分化期功能叶片数变化
在花芽分化过程中需要消耗大量的能量物质,只能依靠光合产物提供。通过测定比较长度大于2cm功能叶总数发现(如图4所示)。乐都长辣椒在5片功能叶时进入门椒花芽分化期,基本每增加2片,花芽分化进程向前一个时期,功能叶片数达到14片时门椒完成花芽分化;华美105则在7片功能叶时进入门椒花芽分化初期,花芽分化初始期功能叶片数多于乐都长辣椒,在门椒完成花芽分化时的功能叶片数(15-16片)也多于乐都长辣椒。综上所述,乐都长辣椒先于华美105 进入花芽分化,华美105在第7片功能叶长成后进入花芽分化期。
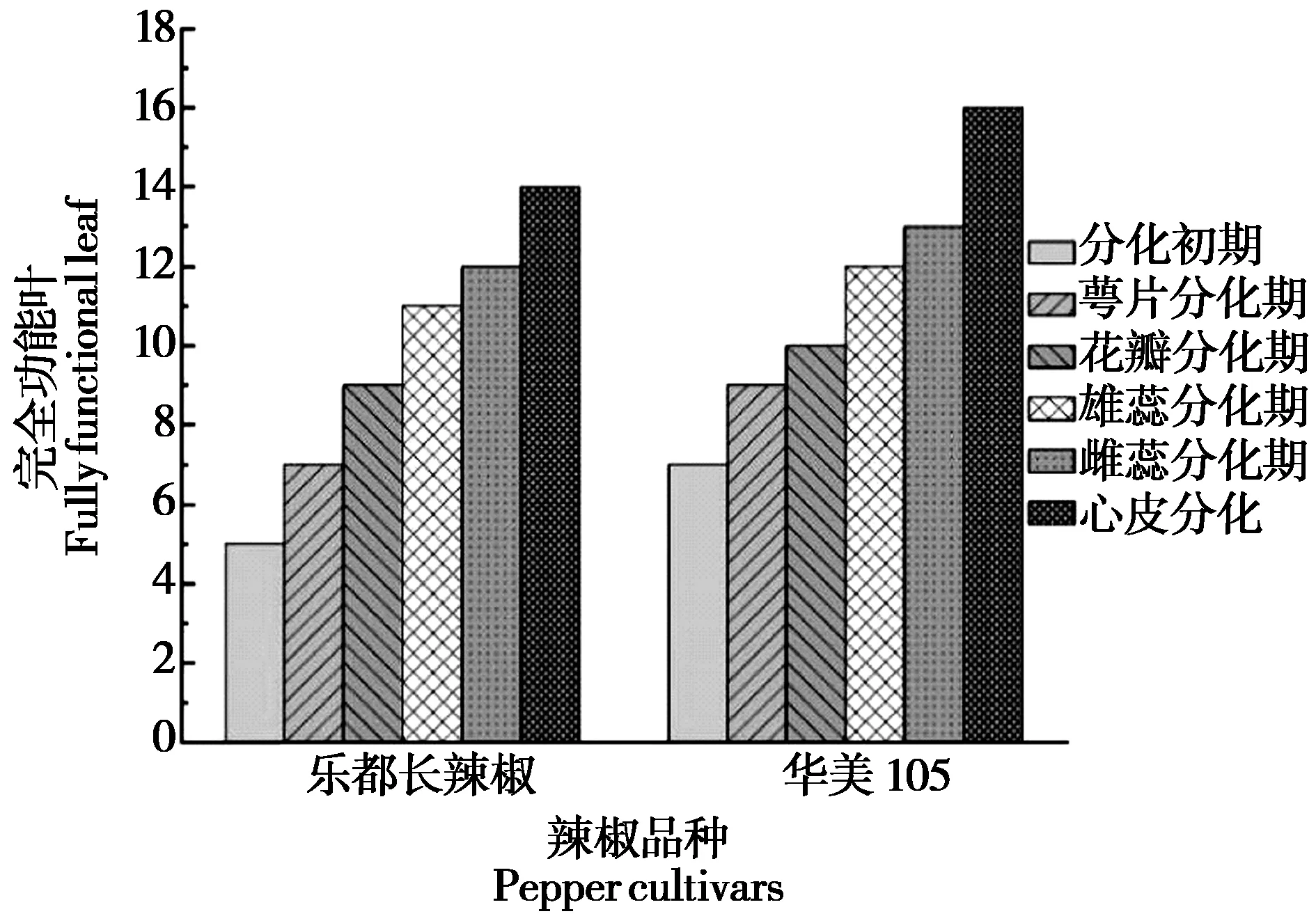
图4 辣椒花芽分化过程中完全功能叶片数的变化 Fig.4 Changes in the number of fully functional leaves during the differentiation of pepper flower buds
2.4 不同耐低温辣椒花芽分化期叶绿素及可溶性糖变化
由图5可知,在辣椒花芽分化过程中,2个辣椒材料叶绿素含量不断增加,在雄蕊分化期叶绿素含量达到同一水平;可溶性糖含量呈先升高后降低的趋势,乐都长辣椒和华美105分别在雌蕊分化期和雄蕊分化期达到峰值。综上所述,随着花芽分化期每一时期的发生,叶绿素含量逐渐上升,可溶性糖在花芽分化的前期升高,花芽分化后期,可溶性糖含量均出现下降趋势,华美105在雄蕊分化期出现拐点,乐都长辣椒拐点出现在雌蕊分化期。

图5 辣椒花芽分化过程中叶绿素和可溶性糖含量的变化Fig.5 Changes in chlorophyll and soluble sugar content during floral bud differentiation in pepper
3 讨论与结论
本试验采用石蜡切片技术和显微镜观察了强耐低温材料华美105和弱耐低温材料乐都长辣椒花芽分化的全过程,通过比较2个材料的5个花芽分化时期发现:乐都长辣椒萼片原基凹陷、花柄原基细而附着点疏松、心皮空腔小而胎座原基宽,表面附着胚珠原细胞数量多;华美105萼片原基较凸出、花柄原基粗而附着点紧致、心皮空腔大而胎座原基呈近锥形,表面附着胚珠原细胞数量较少。由这些显微结构差异可以发现,乐都长辣椒花柄原基细,有可能是其容易低温落花的原因之一,同时乐都长辣椒心皮空腔小,胚珠原细胞数量多是其生殖生长旺盛的特点,这与后期乐都长辣椒果实内种子数量多,容易造成生殖生长过盛,影响后期花芽分化的特点相一致。
结合花芽分化过程中株高、完全功能叶、叶绿素含量和可溶性糖含量的变化,发现辣椒花芽分化过程中弱耐低温的乐都长辣椒花芽分化时期要早于华美105,而且分化初期功能叶片数较华美105少,由于乐都长辣椒生殖分化旺盛,需要更多的光合作用产物支持分化过程,但功能叶少,启动的生理时期又早,这样会造成营养物质和能量代谢失衡,这也可能是乐都长辣椒容易发生低温落花的原因之一。研究还发现雄蕊分化期和雌蕊分化期辣椒细胞分裂和组织速度很快,这一时期的环境条件、营养条件是辣椒能否形成完全花器管的关键时期。
猜你喜欢
食品科学(2022年17期)2022-09-30
中国食用菌(2021年3期)2021-05-08
植物研究(2021年1期)2021-02-26
广西植物(2020年13期)2020-09-12
广西植物(2020年13期)2020-09-12
广西植物(2020年13期)2020-09-12
植物研究(2020年6期)2020-03-05
江苏农业科学(2019年2期)2019-08-13
天津农业科学(2019年1期)2019-07-23
农业科技与装备(2018年5期)2018-02-19
