
多聚体化对蛋白酶生理功能的影响研究进展
2022-03-14 01:47李游山
陕西理工大学学报(自然科学版) 2022年1期
李游山, 杨 玺, 张 杰
(1.陕西理工大学 生物科学与工程学院, 陕西 汉中 723000;2.陕西理工大学 维生素D生理与应用研究所, 陕西 汉中 723000)
蛋白酶是一类能够分解蛋白质肽键的酶,广泛存在于动物、植物、微生物、病毒等生物体内,并在多种生理过程中发挥重要功能。按照催化中心结构的不同可将蛋白酶分为4类:半胱氨酸蛋白酶、丝氨酸蛋白酶、金属蛋白酶和天冬氨酸蛋白酶[1]。半胱氨酸蛋白酶以木瓜蛋白酶、组织蛋白酶C、K等为代表,其催化中心含有Cys残基。丝氨酸蛋白酶包括枯草杆菌蛋白酶、弹性蛋白酶、糜蛋白酶以及胰蛋白酶等,其活性中心含有Ser、His和Asp组成的催化三联体。金属蛋白酶的催化中心含有金属离子,包括内肽酶、氨基肽酶等。天冬氨酸蛋白酶的活性中心含有Asp残基,包括逆转录蛋白酶、胃蛋白酶、组织蛋白酶D等。
蛋白酶在食物消化、凝血、胚胎发生、免疫防御、精子成熟、组织重建、肿瘤发生等诸多生理过程中发挥着重要功能[2]。细胞在转录、翻译、翻译后的激活和降解、蛋白酶抑制剂的抑制等多个层面上对蛋白酶活性进行严格调控[3]。蛋白酶活性失调往往会引发各种疾病。越来越多的数据表明,很多蛋白酶中有多聚体化现象存在,这种多聚体化状态对蛋白酶的结构、活性及酸碱稳定性具有重要的调节作用,从而影响其蛋白水解能力、底物结合能力等生理功能的发挥。蛋白酶的多聚体化在细胞迁移、炎症发生、血管生成、人类免疫缺陷病毒(human immunodeficiency virus,HIV)复制等生理或病理过程的发生、发展中起重要调节作用。近年来,随着人们对蛋白酶多聚体化研究的不断深入,对蛋白酶的多聚体化进行系统总结显得尤为迫切。本文就蛋白酶的多聚体化对其生理功能的影响研究进展做了简要综述,以期为相关领域的研究和应用提供有益参考。
1 半胱氨酸蛋白酶的多聚体化
1.1 半胱氨酸蛋白酶(Caspase)的多聚体化
半胱氨酸蛋白酶(Caspase)是一类内切蛋白酶,在控制炎症和细胞死亡的细胞调节网络中起关键作用[4-5]。这些蛋白酶在聚集成二聚体或大分子复合物后才具有催化活性。Caspase-2是发现较早的Caspase之一,研究表明Caspase-2是细胞对基因毒性等损伤反应的重要调节因子,在凋亡途径中起作用[6]。Caspase-2是通过二聚体化来激活自身活性,并可借助泛素化修饰稳定Caspase-2的二聚体结构,促进其与细胞不溶性组分的结合,并增强其活性,导致细胞完全凋亡[7-8]。最近的研究表明,Caspase-2的激活和催化功能是有丝分裂灾难中触发有丝分裂缺陷细胞凋亡或细胞周期停滞的关键步骤。当对Caspase-2上的保守残基S384进行磷酸化处理后,Caspase-2的催化活性降低,介导的细胞凋亡功能下降,但并不影响Caspase-2的二聚体化[9]。Caspase-3前体为非活性的同源二聚体结构,在变构激活剂的作用下发生构象变化,转化为活性状态的二聚体结构,随后在颗粒酶B的作用下形成成熟体,活性的前体和成熟体都可促进细胞凋亡(图1[10])。Caspase-3前体的二聚体界面在其折叠和组装中具有重要意义,向该界面区域引入V266H突变会使Caspase-3成熟体活性完全丧失[11]。向二聚体界面的L4环中引入C242A或E246A突变则会降低Caspase-3二聚体的酸碱稳定性[12]。二聚体界面是Caspase-3前体的变构调节区,对酶的活性调节至关重要,因此促进癌细胞中procaspase-3的激活可作为一种治疗策略[10-11]。与Caspase-3前体不同的是,未切割的Caspase-8前体的二聚体也具有足够的活性来启动细胞凋亡,但激活的过程同样取决于Caspase-8前体的二聚体化。向二聚体界面引入P466I或F468S/V突变可促进Caspase-8的二聚体化[13]。凋亡引发因子Caspase-9的激活是诱导细胞凋亡的关键步骤,在正常的生理浓度下Caspase 9主要以无活性的酶原单体形式存在,高浓度的Caspase-9倾向于二聚体化,从而激活Caspase-9活性[14]。这些研究结果提示半胱氨酸蛋白酶Caspase-2、3、8、9的二聚体化可显著增强酶的活性。
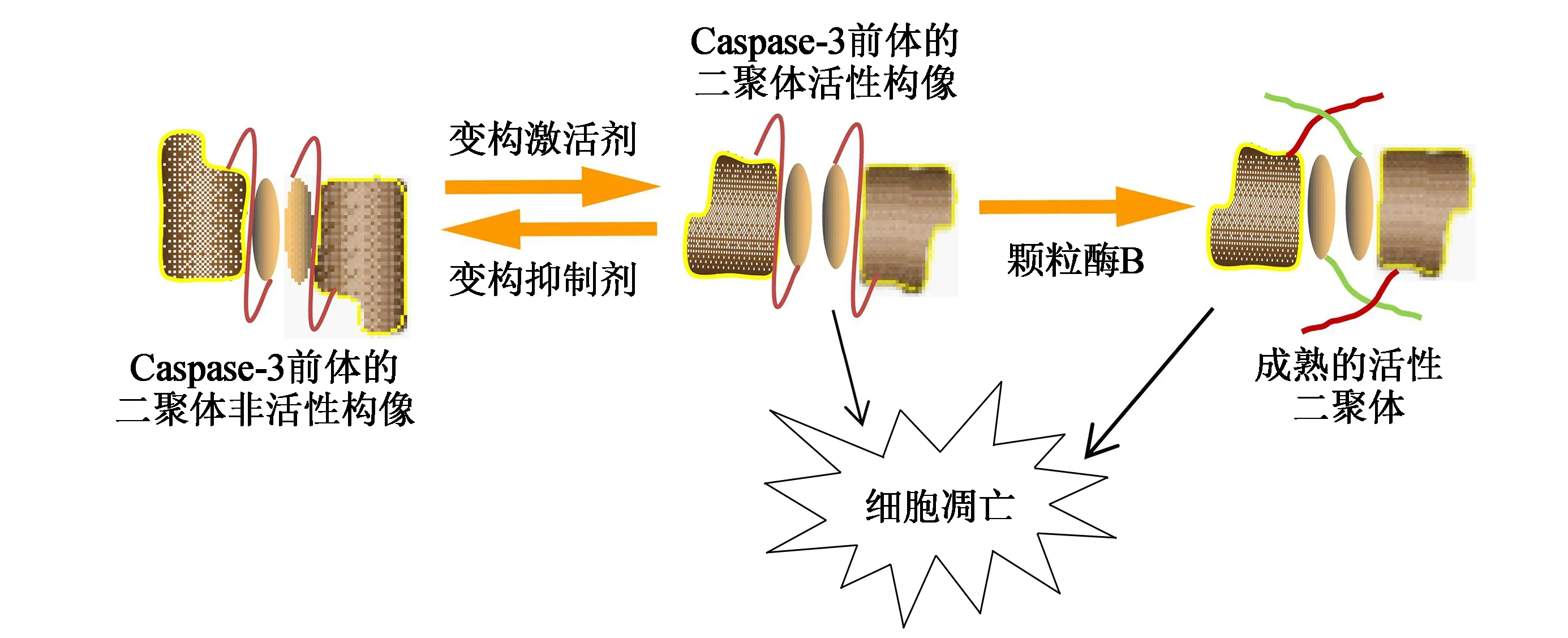
图1 Caspase-3介导细胞凋亡
1.2 组织蛋白酶的多聚体化
组织蛋白酶C(Cathepsin C)属于半胱氨酸蛋白酶,参与细胞内蛋白质的非特异性降解过程。组织蛋白酶C还可与TNF-α/p38MAPK信号通路相互作用,促进肝细胞癌的增殖和转移[15]。成熟的牛组织蛋白酶C分子由4个相同的单体组成。在组织蛋白酶C酶原四聚体的形成过程中,其活性催化位点Cys331暴露,进而被表面疏水基团包围,形成了二聚体-二聚体相互作用界面。研究显示,组织蛋白酶C酶原四聚体的形成使每个单体单元的内肽酶活性位点被包裹而不具备活性,但在其翻译后激活阶段又解离成二聚体发挥作用[16]。组织蛋白酶K(Cathepsin K)是破骨细胞在骨吸收过程中降解胶原蛋白所需的半胱氨酸蛋白酶,其活性位点抑制剂的研究受到广泛关注。Aguda等(2014)[17]解析了组织蛋白酶K(Cathepsin K)的蛋白二聚体结构,发现1个组织蛋白酶K分子可以和胶原蛋白结合的糖胺聚糖结合,并招募第二个蛋白酶分子,进而揭示了组织蛋白酶K水解胶原蛋白的分子机制。组织蛋白酶K二聚体和糖胺聚糖的结合位点是一个新的靶向位点,基于这类药物靶点开发第二代抑制剂来抑制组织蛋白酶K活性,可有效减少骨吸收,比传统的抗骨质疏松药物更有优势[17-19]。
1.3 其他
在嗜热菌(Pyrococcushorikoshii)OT3中也发现了一种蛋白酶PH1704,属于半胱氨酸蛋白酶,这种酶具有变性剂抗性和高度的热稳定性。詹冬玲等(2014)[20]利用大肠杆菌(Escherichiacoli)BLP-CodonPlus高效表达PH1704蛋白,发现该酶主要以十二聚体的形式存在,并具有SDS抗性。十二聚体以上的高聚合物经过SDS处理后变成以六聚体为主的混合物,蛋白酶活力染色表明,其六聚体以上的高阶聚合物有蛋白酶活力,而单体不具备蛋白酶活力[21]。许多多亚基蛋白/酶都具有别构潜力,都是别构蛋白/酶,其中PH1704就是别构酶,这种别构效应可通过影响亚基聚合来影响酶的亲核催化[20]。因此我们猜测多聚体化可能会影响蛋白酶的别构,这一点还有待进一步研究。这类嗜极微生物的酶在高温及极端pH下稳定,对较高的离子强度和溶剂具有较好的耐受性,非常适合用作食品、饲料、制药、纺织、洗涤剂等行业的生物催化剂。目前也可通过蛋白质工程等现代技术对这类嗜极微生物的酶进行改造,以获得最佳性能。
2 丝氨酸蛋白酶的多聚体化
2.1 类胰蛋白酶的多聚体化
人的类胰蛋白酶(tryptase)是一种胰蛋白酶样丝氨酸蛋白酶,主要存在于肥大细胞的分泌颗粒中,可能在炎症和宿主防御中发挥关键作用,是哮喘过敏性炎症反应的重要介质[22]。人类的β-tryptase以非共价连接的四聚体形式来表现酶活,这种四聚体形式主要依赖肝素稳定,对大多数蛋白质类抑制剂都具有抗性(图2)[23-24]。四聚体的活性不受蛋白质类抑制剂的影响,而是可以通过构象变化来调节自身活性。在缺乏肝素或低盐条件下,活性四聚体会转变成非活性不稳定的四聚体,该四聚体在稀释后可逆地解离为非活性单体(图2)[25]。四聚体中每条β-tryptase分子都有两个不同的作用,既是蛋白酶,也是其相邻蛋白酶分子的辅助因子,可以变构调节酶活性,以稳定性四聚体结构提高蛋白水解活性[22]。通过创建肝素缺乏的环境,对小鼠肥大细胞特异性表达的胰蛋白酶进行抑制,可以在类风湿性关节炎的治疗中发挥作用[25],也可以通过合成抗β-tryptase的二聚体药物来调节胞内炎症反应[24,26]。

图2 β-tryptase的活性形式及结构变化
2.2 其他丝氨酸蛋白酶的多聚体化
Swamy等(1983)[27]从大肠杆菌(E.coli)中鉴定到6种丝氨酸蛋白酶,可以水解14C-球蛋白和3H-酪蛋白,分别命名为蛋白酶Do、Re、Mi、Fa、So、La。其中,蛋白酶Do(Protease Do)是一种胞质丝氨酸蛋白酶,该酶以十聚体或六聚体或五聚体的形式存在并水解酪蛋白。Huesgen等(2007)[28]对来自集胞藻(Synechocystissp.)PCC 6803菌株的HhoA蛋白酶进行了表征,这是一种的六聚体丝氨酸蛋白酶,是细胞周质中与质膜相关的可溶性蛋白,该酶需形成具有蛋白水解活性的同源六聚体复合物后方能降解底物。HhoA的蛋白水解活性随温度和pH值的增加而增加,并受Mg2+或Ca2+的刺激,其上的PDZ结构域在调节该酶活性和六聚体复合体的组装中起着至关重要的作用。从嗜热菌(P.horikoshii)中也发现了一种六聚体丝氨酸蛋白酶PhAAP,晶体结构解析发现它的底物筛选策略比较特殊,底物首先通过六聚体表面上的气孔,然后转向通过单体结构域界面处的更小开口,需通过这样的双门控“检查”系统来访问单体的活性位点[29]。同时,在丝氨酸蛋白酶中也存在八聚体化现象,例如拟南芥(Arabidopsis)同源基因Deg9编码的蛋白酶,位于细胞核中,可通过降解拟南芥应答调节因子4(Arabidopsis response regulator4,ARR4)来调节细胞分裂素和光介导的信号传导。对Deg9蛋白酶结构进行解析,Deg9蛋白酶呈现为八聚体结构,该结构由2个具有不同构象的四聚体环组成,一个四聚体的蛋白酶活化是通过蛋白酶结构域的整体重新定向来介导的,以打开相对应的另一个无活性的四聚体的底物结合位点[30]。上述研究表明,蛋白酶Do、HhoA、Deg9等主要以多聚体的形式存在并执行水解底物功能,但单体是否具备活性仍需进一步探讨。
3 金属蛋白酶的多聚体化
3.1 基质金属蛋白酶MMP的多聚体化
基质金属蛋白酶-9(matrix metalloproteinase-9,MMP-9)是MMP家族的一员,在正常和病理条件下与细胞外基质的降解、炎症发生有关,可作为各种临床反应的潜在生物标志物[31-32]。MMP-9能够以二硫键链接的二聚体形式存在[33]。Olson等(2000)[34]纯化出MMP-9前体的单体和二聚体形式,证明经过活化后,这两种酶形式均可切割肽底物的形成,且切割能力相同。研究表明,MMP的活性会影响肿瘤细胞的迁移[35-36]。Dufour等(2008)[37]证实MMP-9中的血红素蛋白(PEX)结构域是增强细胞迁移的先决条件。MMP-9的PEX结构域会发生二聚体化现象,这种二聚体化可以有效增强细胞迁移。通过设计一些模拟PEX结构基序的抑制肽,发现这些抑制肽可以有效地阻断MMP-9二聚体的形成[38]。佛波醇-12-肉豆蔻酸酯-13-乙酸酯(phorbol 12-myristate 13-acetate,PMA)是一种佛波酯,是蛋白激酶C和鞘氨醇激酶的激活剂。用PMA处理来源于正常人肌肉、上皮和结缔组织的细胞时,在这些细胞中都未观察到MMP-9二聚体的分泌,表明PMA会抑制MMP-9二聚体的形成[39]。基质金属蛋白酶-14(MMP-14)是一种膜锚蛋白,与肿瘤转移过程中细胞外基质重塑和细胞迁移密切相关。它的功能域结构与MMP-9十分相似,也存在一个血红素结合蛋白(hemopexin,PEX)结构域,该结构域可以引发MMP-14的同源二聚体化以及与细胞表面粘附分子CD44的异源二聚体化。MMP-14与CD44之间的相互作用导致表皮生长因子(EGF)受体的磷酸化,进而激活下游细胞迁移有关的MAPK和PI3K信号通路。同样,模拟MMP-14的PEX结构域中最外链的关键基序设计抑制肽,发现合成肽IS4和IVS4以剂量依赖性的方式抑制MMP-14介导的细胞迁移,这些肽会干扰肿瘤转移,但不影响原发肿瘤的生长。因此,靶向抑制MMP-14的血红蛋白结构域,可以作为一种预防癌细胞扩散的新方法[40]。膜1型基质金属蛋白酶(MT1-MMP)的同源二聚体化,是激活MMP-2前体和降解细胞表面胶原蛋白的重要步骤。对MT1-MMP的晶体结构进行解析,发现其上的PEX结构域是驱动二聚体化的关键。而当向MT1-MMP的PEX结构域中引入DTY/KAF、DTY/AAF、RWD/AAK氨基酸突变时,显著抑制PEX结构域的二聚体化作用,从而抑制细胞表面上的二聚体依赖性功能,包括MMP-2前体的激活、胶原蛋白的降解,以及对三维胶原基质的侵袭;但对非二聚体化依赖性的其它功能没有影响[41]。解整合素金属蛋白酶17(ADAM17)是一种跨膜的锌依赖性金属蛋白酶,其催化活性导致广泛的膜蛋白脱落。该酶胞外部分包括多个结构域,其胞外域的释放可引起各种生理和病理过程的转换。研究发现,该酶以二聚体的形式存在于细胞膜中,且多聚体化是由其表皮生长因子(EGF)样结构域介导的[42]。综上所述,基质金属蛋白酶MMP存在二聚体结构,其上的PEX结构域是形成二聚体的关键区域,并能影响细胞迁移、胶原蛋白降解等功能。
3.2 内肽酶Meprin的多聚体化
内肽酶Meprins是哺乳动物细胞表面的一类含锌金属蛋白酶,以膜结合或分泌形式在肠和肾的刷状缘上皮细胞、上皮肠粘膜中的白细胞,以及各种癌细胞和组织中表达。迄今为止,人们发现Meprins表现出更广泛的表达模式,在血管生成、癌症、炎症、纤维化和神经退行性疾病中发挥作用[43-44]。Meprins包含α、β两种亚基,亚基间通过二硫键连接成同型二聚体和异型二聚体,并由非共价相互作用介导了进一步的寡聚化,从而对不同的底物发挥作用[45]。α和β亚基具有高度的序列一致性,但是它们的底物特异性和寡聚化潜能是极为不同的。Meprin α亚基通常会形成多聚体,且能够分泌到细胞外。Meprin β亚基通常会形成四聚体结构,并能够以多聚体形式结合到细胞膜上(图3[45])。含有α亚基的同型或异型多聚体被命名为Meprin A(EC 3.4.24.18),而β亚基的同型多聚体被称为Meprin B(EC 3.4.24.63)。从肾脏和肠上皮细胞分泌的Meprin A是一种能够切割生长因子、细胞外基质蛋白和生物活性肽的金属蛋白酶。利用凝胶过滤、非变性凝胶电泳和交联等方法对小鼠Meprin A多聚体形式进行分析,结果表明,该酶的分泌形式是一种由α亚基组成的同型多聚体,且以十聚体为主。该多聚体是由二硫键连接的二聚体通过非共价连接而成,这种寡聚体的形成对蛋白质底物的水解至关重要[44,46]。同时Meprin A还是一种高度糖基化的蛋白酶,去除蛋白酶结构域中的糖基化位点Asn234和Asn452使其化学稳定性和热稳定性降低,而Asn152和Asn270缺失则直接导致酶活丧失[47]。此外,Meprin亚基受理化因素的调节表现出不同的生理活性。Meprin A在低盐浓度下被抑制,但随着盐浓度的增加而被激活,而增加盐浓度通常却会抑制Meprin B的活性,这对阐明Meprin和其他金属蛋白酶的催化机制具有重要意义。组织中的Meprin寡聚物的活性还受到pH和NaCl变化的影响,这些研究结果对Meprin相关的炎症和癌症的治疗具有至关重要的意义[48]。

Meprin α和Meprin β均包含PRO、CAT、MAM、TRAF、EGF、T和C结构域,其中Meprin α还包含一个I域
4 天冬氨酸蛋白酶的多聚体化
天冬氨酸蛋白酶(aspartic protease)是一类重要的蛋白水解酶,参与机体的新陈代谢及生物调控,已广泛应用在食品工业等生产领域[49],它的活性中心由两个催化性Asp残基组成。天冬氨酸蛋白酶包括逆转录蛋白酶亚型、胃蛋白酶亚型、天冬氨酸膜内裂解蛋白酶亚型等。
4.1 逆转录病毒蛋白酶的多聚体化
大量前瞻性研究表明逆转录病毒蛋白酶(retroviral protease,PR)属于胃蛋白酶样天冬氨酸蛋白酶家族,而天冬氨酸蛋白酶是由一种较小的二聚体祖先蛋白进化而来的。研究显示,二聚体结构使蛋白酶更加稳定。对Rous肉瘤病毒(RSV)蛋白酶晶体结构进行解析,发现RSV蛋白酶为二聚体且具有与天冬氨酸蛋白酶相似的结构,主要切割逆转录病毒gag、pol和env基因翻译后产生的前体蛋白,以产生在病毒中发现的成熟蛋白[50]。Katoh等(1989)[51]通过生化分子质量测定及交联实验发现,逆转录病毒蛋白酶分子会自组装成二聚体。研究发现,人免疫缺陷病毒HIV-1蛋白酶(HIV-PR)也是一种二聚体结构,该酶对于HIV的复制至关重要,可以裂解HIV病毒颗粒的前体使其成熟,从而被公认为是抗病毒治疗的靶酶。通过合成基因在大肠杆菌中表达,获得了一种单链链接的HIV-PR二聚体。该单链二聚体由两个含99个氨基酸的HIV-PR亚基组成,并通过五肽连接在一起。在pH 5.5条件下,单链二聚体和天然状态下的HIV-PR二聚体的酶学性质相似。然而在pH 7.0时,单链二聚体比天然HIV-PR二聚体更稳定,这是由于天然HIV-PR二聚体在酸性条件下解离。将其活性位点Asp残基突变为Gly残基,该蛋白酶失去活性,表明二聚体形成中各亚基的Asp残基构成了HIV-PR的活性位点[52]。
4.2 其他天冬氨酸蛋白酶的多聚体化
猪的胃蛋白酶(Pepsase)自溶会裂解成没有活性的片段,而只有在二聚体化后方能显示出蛋白水解活性。二聚体形成使胃蛋白酶对周围环境更具耐受性,酸碱稳定性和抗自溶能力增强,即使是在零度以下的低温条件依然保持良好活性[53]。恶性疟原虫(Plasmodiumfalciparum)的组织天冬氨酸蛋白酶(histo-aspartic protease,HAP)由于其独特的结构而成为一种有趣的天冬氨酸蛋白酶。Xiao等(2010)[54]构建了含硫氧还蛋白标签的重组融合蛋白Trx-tHAP,发现Trx-tHAP融合蛋白在纯化过程中会发生聚集,而加入0.2%的表面活性剂CHAPS可以防止这种聚集。Trx-tHAP融合蛋白通过自激活和肠激酶或酶plasmepsin II的激活形成成熟的tHAP。凝胶过滤层析等结果显示,成熟的tHAP存在于一个动态的单体-二聚体平衡中,当CHAPS存在的时,该酶的二聚体解离速率增大。酶活数据显示,HAP的单体形式活性最强,多聚体化反而抑制了它的活性。马铃薯(Solanumtuberosum)中天冬氨酸蛋白酶StAP中的saposin样结构域是一个开放的saposin折叠二聚体,其主要功能是细胞内分选和宿主防御[55-56]。该酶二聚体形成是pH依赖性的,从非活性单体到活性二聚体需依赖酸化过程。可见,并非所有的蛋白酶寡聚化都可提高酶的活性,部分多聚体化现象反而抑制酶的活性,因此,仍需针对不同蛋白酶的多聚体化所介导的功能进行具体研究。
5 结语与展望
笔者对已报道的半胱氨酸蛋白酶、丝氨酸蛋白酶、金属蛋白酶和天冬氨酸蛋白酶的多聚体化研究进行简要汇总(表1),发现在人和小鼠中发现的半胱氨酸蛋白酶、金属蛋白酶、天冬氨酸蛋白酶均以二聚体的形式表现酶活,进而水解底物,与胞外基质降解、细胞迁移及炎症反应密切相关。而多聚体化的丝氨酸蛋白酶多数在细菌中被鉴定到,主要表现形式为六聚体及更高阶的多聚体形式。

表1 已报道蛋白酶的多聚体化
蛋白酶的多聚体化研究具有重要的研究价值。大量研究发现多聚体化可稳定蛋白酶的高级结构、理化稳定性、提高它们的体外活性,并对其介导的底物水解、细胞迁移、炎症反应等生物学功能都具重大影响。本文通过对蛋白酶的多聚体化研究进行综述,发现当前蛋白酶多聚体化的研究从结构到功能都已十分丰富,但其影响机制的研究尚不完全清楚,仍需更多的实验数据支撑。另一方面,蛋白酶的多聚体化应用在产业上的报道仍然较少。认识蛋白酶的多聚体化在机体生理和病理过程的重要作用及它们的形成机制,有助于设计抑制肽靶向抑制蛋白酶多聚体的生成,从而用作治疗肿瘤细胞迁移、骨吸收、炎症反应、病毒复制等人类疾病的新的药物作用靶点。同时对蛋白酶多聚体化的关注还可服务于食品生产、制药等工业领域,通过基因工程及蛋白质工程的手段,可以筛选出性能最佳的蛋白酶产品用作生物催化剂,从而造福于人。
猜你喜欢
沈阳医学院学报(2022年6期)2022-12-03
中国动物保健(2022年9期)2022-09-25
南昌大学学报(医学版)(2021年1期)2021-11-29
化工设计通讯(2017年10期)2017-03-02
海南医学(2016年8期)2016-06-08
中国卫生标准管理(2015年16期)2016-01-20
听力学及言语疾病杂志(2015年5期)2015-12-24
现代检验医学杂志(2015年2期)2015-02-06
现代检验医学杂志(2015年1期)2015-02-06
现代检验医学杂志(2014年5期)2014-02-02
