
希腊牛至辐射诱变突变群体的构建和筛选
2022-03-26 02:15郭贺峰殷庭超刘信宝
草业科学 2022年3期
郭贺峰,何 悦,殷庭超,蔡 冬,刘信宝,张 敬,徐 彬
(1.南京农业大学草业学院, 江苏 南京 210095;2.福建傲农生物科技集团股份有限公司, 福建 漳州363000;3.福建省生猪营养与饲料重点实验室,福建 漳州 363000)
牛至(Origanum vulgare)是唇形科牛至属多年生草本或半灌木植物,也被称为止痢草、牛膝草、马郁兰、小叶薄荷等,是典型的多年生芳香植物。其分布范围广泛,原产于欧亚大陆及部分非洲地区,我国大部分省区皆有野生牛至分布[1]。牛至又可被分为6 个亚种,其中希腊牛至(O.vulgaressp.hirtum)香味浓郁、精油含量高、抑菌效果好,被广为栽培应用[2]。牛至用途多样,全草皆可入药,是一味传统的中草药,对急性肠胃炎、腹泻等症具有很好的疗效[3];牛至香辛味浓郁,能够改良食物风味,因此在西班牙、墨西哥、希腊等国被当作香料长期使用[4];牛至花期长、花型优美、花色艳丽,既可用于观赏,还是一种很好的蜜源植物;此外,牛至花、叶可提取芳香精油,具有较强的抑菌和抗氧化作用[5-7],因此还被广泛应用于动物饲料添加剂,能够保护动物肠道健康,改善肉产品品质,提高生长性能,降低死亡率,减少疾病,提高饲料利用率等[8-15]。近年来,牛至干草和牛至精油的需求量逐年增加,现有野生牛至资源已不能满足生产需求;另一方面,牛至干草价格较高,限制了其在饲料添加剂中的生产应用。培育高产、优质牛至新品种是解决牛至供需矛盾与应用瓶颈的关键环节之一。
辐射诱变是一种快速培育新品种的方法,具有突变频率高、变异范围广、后代性状稳定快、育种年限短等优点[16]。牛至种子极小,千粒重仅0.1 g,可能对辐射诱变敏感,是诱变育种的理想材料。另外,牛至可以通过扦插或组培等方式扩繁[17],因此牛至突变体单株能够通过营养体扩繁保留具有特殊变异性状。
本研究以南京农业大学草业学院前期筛得到的希腊牛至为材料,利用60Co-γ 射线作为辐射源处理希腊牛至种子,旨在找出适宜牛至种子的辐照剂量,获得牛至突变体种质资源库,筛选出具有优异变异表型的突变体,为牛至诱变系品种改良和选育提供基础。
1 材料与方法
1.1 试验地点
本研究在南京航空航天大学辐照中心进行辐照处理,辐射源为60Co-γ 射线源,装源量为200 万Ci。在全自动化玻璃温室内育苗,温室内的昼夜温度28 ℃/23 ℃,实际温度为设定温度的 ± 5 ℃,空气湿度保持在25%~42%。幼苗移栽定植到南京农业大学白马教学科研基地的试验田 (119°18′71.96″ E,31°62′0.06″ N)。试验田土壤质地为黏壤土,容重为1.27 g·cm-3,比重为2.52 g·cm-3,土壤总孔隙度为49.53%,pH 7.45,EC 值为53.21 μS·cm-1,有机质含量为0.82%。
1.2 试验材料
母本种子材料采自南京农业大学草业学院筛选的希腊牛至株系,母本植株的叶片精油含量4.2%,花的精油含量5.3%,全株(包括茎、叶和花)精油含量3.2%。
1.3 试验方法
1.3.1 辐照处理
共设置6 个60Co-γ 射线辐照剂量梯度,分别为0、100、200、400、800 和1 600 Gy,剂量率为1.61 Gy·min-1,每个剂量处理置于塑料自封袋中的2 g 种子(约2 万粒),以未经辐照的种子作为对照(CK)。
1.3.2 发芽试验
将辐照后的种子与未处理对照种子均匀摆放在铺有两层滤纸的培养皿中,每皿100 粒种子,每个剂量4 个重复,每天定时定量洒水以保持发芽所需湿度,每天记录发芽数,并根据发芽数计算发芽率、发芽势和发芽指数。
发芽率 = 发芽数/每皿粒数×100%;
发芽势 = 5 d 内发芽种子粒数/供试种子数×100%;

式中:Gt为发芽试验终期内每日发芽数,Dt为发芽日数。
1.3.3 育苗移栽
种子用10 mg·L-1赤霉素溶液处理后[18],在温室育苗,育苗基质采用丹麦品式托普泥炭土(pH 5.5~6.0,粒径 < 6 mm),将泥炭土打湿后分装于72 孔穴盘中;依据发芽率计算所用穴盘数与用种量,将种子与细蛭石混合,均匀播于穴盘内,每天喷水保持基质湿润。种子发芽10 d 后间苗,保持每穴1 株小苗。于2021 年3 月中旬,待幼苗生长至8 cm 高时移栽大田,移栽前试验田提前挖好排水沟确保排水通畅并喷洒封闭型除草剂隔绝杂草,株行距均设置为30 cm。
1.3.4 突变体表型性状测定
2021 年7 月 - 8 月,通过对田间突变表型的观察,筛选出变异明显的株系,测定对照株系和突变体的表观性状,包括株高、叶长、叶宽、茎节长度和分枝数。使用电子游标卡尺测量植株顶端向下第3 节间已舒展叶片最宽和最长处即叶长、叶宽实际值。茎节长度的测定使用电子游标卡尺测量植株顶端向下第3 节节间长度。除株高与分枝数测量外,其余各指标均测量4 个分枝并取其均值。
1.4 数据处理
采用Excel 2016 进行数据统计与制图,SPSS 13.0 软件进行单因素方差分析比较数据。
2 结果与分析
2.1 辐射对牛至种子发芽和成苗的影响
100 Gy 辐照剂量的60Co-γ 射线对牛至种子发芽率、发芽势和发芽指数均没有显著抑制作用(P>0.05) (表1),但更高的辐射剂量显著抑制了牛至种子萌发(P< 0.05),且随辐射剂量的增大,抑制作用显著增加,牛至种子发芽率、发芽势和发芽指数呈显著降低的趋势(P< 0.05)。辐照处理还推迟了牛至种子的发芽时间(图1),对照和100~200 Gy 辐射剂量处理的种子在第4~5 天时发芽最多,400~800 Gy 剂量处理的种子在第5~6 天时发芽最多,而1 600 Gy 剂量处理导致第7 天时才达到发芽高峰期。根据种子发芽情况,以发芽率为因变量(y),辐射剂量为自变量(x),得到发芽率与辐射剂量的线性回归方程y= -0.000 6x+ 0.649 9,并将对照发芽率的一半(即37.6%)带入方程,得到半致死辐射剂量LD50为539 Gy。

表1 辐照剂量对牛至种子发芽指标的影响Table 1 Effects of irradiation dose on germination index of oregano seeds

图1 60Co-γ 射线对牛至每天发芽数的影响和发芽率-辐射剂量回归方程Figure 1 Effect of 60Co-γ rays on number of germinating oregano seeds and regression equation between germination rate and radiation dose
此外,辐照处理后牛至种子在萌发后胚根生长受抑制,根系发育缓慢,导致萌发后幼苗在育苗基质内成活率低,幼苗早期生长缓慢。与对照的田间成苗率(58.20%)相比,100 Gy 辐照剂量导致田间成苗率下降(39.00%),200 Gy 剂量导致田间成苗率降至1.85%,更高剂量辐照处理的种子萌发后无法在育苗基质中成活。
2.2 牛至突变体筛选
将辐射诱变后的牛至种子苗移栽至大田,共获得8 170 株M1群体。将同一时期萌发、未经辐射处理的1 000 株牛至作为对照,观察、记录对照群体和M1单株的表型特征,并扦插扩繁M1群体内具有明显表型变异特征的单株。
对照株系的株高32~46 cm,分枝数18~34,叶片翠绿至深绿色,第3 节间长8.9~10.4 mm,叶面积152~165 mm2(图2),伞房状圆锥花序,多花密集,花白色。参考对照植株的表型特征,将具有明显色泽变化或超过可量化指标极值10%的株系作为M1代突变体,包括高杆和矮杆突变体型、多分枝和少分枝突变体、叶型(叶片大小和形状)、叶色及花色突变体等(表2)。这些突变体在扦插扩繁后仍能保持相应变异特性,说明变异特征与立地土壤或水肥条件无关,均为遗传变异导致的突变性状。
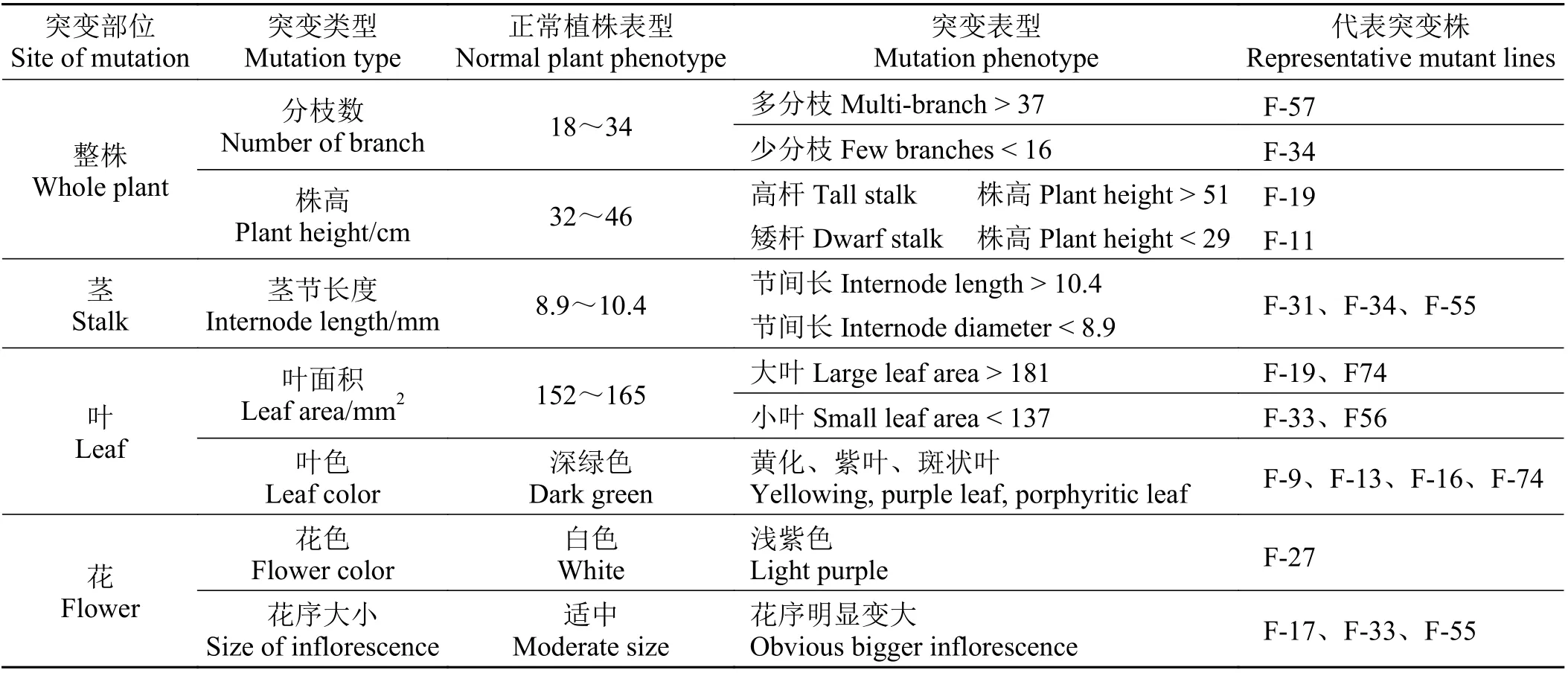
表2 辐射突变群体的突变类型及评判标准Table 2 Mutant types and trait evaluation standard in the radiation mutagenized population
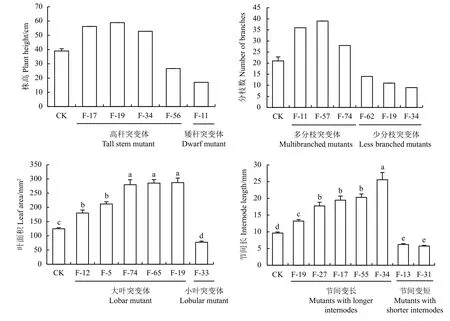
图2 部分希腊牛至突变体的株高、分枝、叶面积和节间长度Figure 2 Greek oregano mutants with typical variations of plant height, branching, leaf area, and internode length
株高变异体:根据1 000 株对照牛至的株高生长范围(32~46 cm),将株高 > 51 cm 或 < 29 cm 的株系定为高杆或矮杆突变体。例如, F-19 株高达到59 cm,而F-11 株高仅17 cm,与对照株系株高均值(39 cm)差异明显(图2)。
茎节长度突变体:短茎节突变体F-31 的平均茎节长较对照缩短了40.3%,而长茎节突变体F-34 较对照增长了166.0% (图2)。
分枝突变体:根据对照牛至的分枝数范围(18~34 个),将分枝数 > 37 或 < 16 的株系定为多分枝或少分枝突变体。例如,多分枝突变体F-57 的分枝数达到39 个,而少分枝突变体F-34 仅有9 个分枝,与对照株系分枝(21 个)差异明显(图2)。
叶色变异体:在M1突变群体中,还发现具有叶色黄化变异株系3 株,红色变异株系1 株(图3)。

图3 典型叶色变异突变体Figure 3 Typical leaf color mutants
大叶及小叶突变体:在突变体群体中,还筛选到具有明显叶片大小变化的突变体(图4)。以自上而下第3 节间基部舒展叶为例,对照株系的叶面积为124.66 mm2,叶长在10~23 mm,平均值为14.34 mm,叶宽在9~19 mm,平均值为11.07 mm;而大叶突变体F-19 的叶面积达到287.02 mm2,叶长达到22.63 mm,叶宽达到16.13 mm;小叶突变体F-33 的叶面积仅77.24 mm2,叶长仅10.10 mm,叶宽仅9.10 mm,与对照株系差异显著(P< 0.05) (表3)。
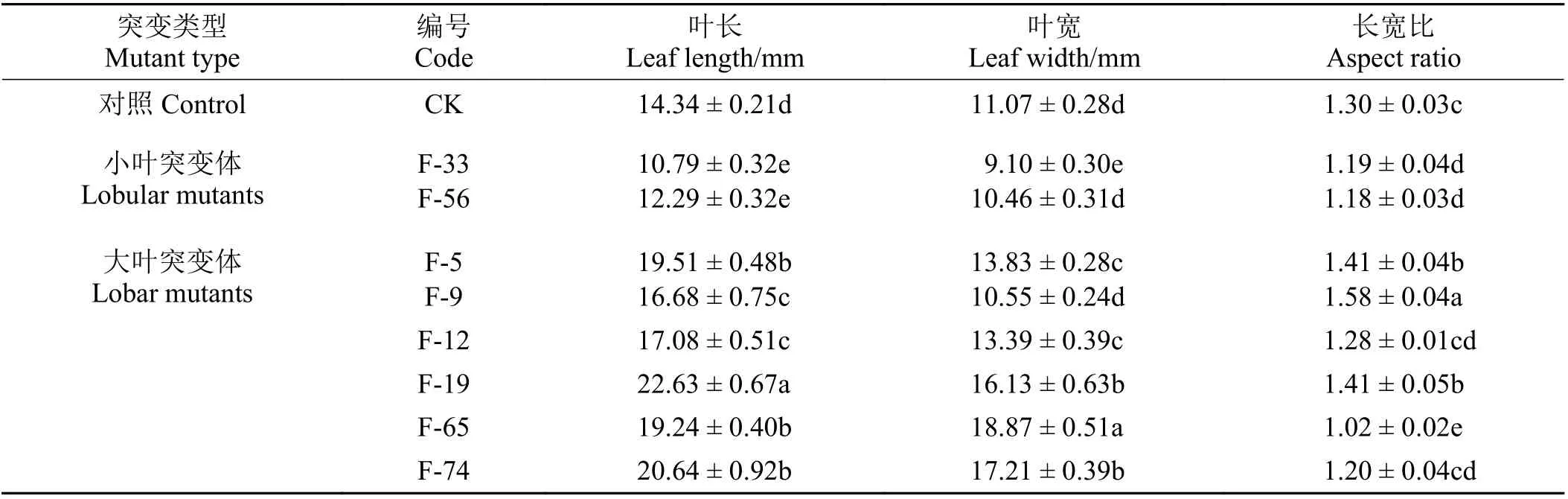
表3 辐照处理导致牛至叶长、叶宽、叶面积发生变化Table 3 Irradiation treatment causes changes in oregano leaf length, leaf width, and leaf surface area

图4 典型叶片大小突变体Figure 4 Typical leaf size of the mutants
3 讨论
60Co-γ 射线诱变是最安全高效的诱变技术之一,被广泛应用于植物的诱变育种中。例如,对水稻(Oryza sativa)品种‘圣稻15’进行诱变并通过筛选和鉴定获得93 份表型突变较为明显的突变体[19];利用60Co-γ 射线辐照花生(Arachis hypogaea)的干种子,获得高油突变体和高蛋白突变体[20];利用60Co-γ射线对扁穗牛鞭草(Hemarthria compressa)处理发现,辐照后的大部分牛鞭草叶片趋于变小、株高趋于变矮、茎趋于变细[21]。另外,辐射诱变营养体或种子的再生植株易出现突变嵌合体[22-24]。本研究在牛至M1代突变体中的观察发现,各突变体的变异表型在同一株的不同分枝较为一致,说明存在嵌合体的可能性小,这可能与牛至种子(胚)极小且选用较高的辐射剂量(100~200 Gy)有关,另外,本研究通过枝条扦插的方式扩繁变异株,进一步避免了辐射诱变可能导致的嵌合体问题。
高剂量辐射会抑制种子发芽率、生长势和根系生长[25-28]。本研究结果表明,当辐射剂量超过200 Gy时,希腊牛至种子的发芽率、发芽势和发芽指数随辐射剂量的增大而显著降低。虽然超过400 Gy 辐射剂量的种子仍能够发芽,但萌发后的种子胚根尖部坏死,无法生成主根或产生侧根,播种至育苗基质后,苗期即全部死亡。适宜的辐射剂量选择与植物种类、种子大小、辐射部位及辐射材料的含水量关系密切。徐冠仁[29]认为致死剂量的60%~70%才是最佳辐射剂量;对桂花(Osmanthusfragrans)的研究认为,适宜的辐照剂量与品种有很大关系[30];史燕山等[31]则以半致死和半致矮剂量为标准对晚香玉(Polianthes tuberosa)的种球进行辐照处理,确定了晚香玉大小种球的适宜辐照剂量。本研究通过发芽率-辐射剂量回归方程计算得出牛至辐照的半致死剂量为539 Gy,与研究结果有较大的差异,半致死剂量不能作为牛至的适宜辐照剂量。结合本研究结果得出结论,希腊牛至辐照处理的适宜剂量应控制在致死剂量的18.6%~37.1%为宜,这与其他植物辐射诱变研究的结论有较大差异,可能是由于希腊牛至种子极小,对60Co-γ 射线更加敏感。因此,适宜辐射剂量的选择不能仅参照发芽率或发芽势,还应综合考虑辐射对幼苗生长发育的影响。
利用60Co-γ 射线辐射牛至种子的近期研究发现,在不同辐射剂量下处理40 粒种子,牛至的半致死剂量仅为16.39 Gy[32]。本研究的母本材料是自主筛选得到的希腊牛至优株,每个辐射剂量处理了约10 000 粒种子,在100~200 Gy 的辐照强度下获得大量突变体材料,并在田间条件下筛选有显著表型变异的突变体。本研究结果与闫一皓等[32]报道差异较大,可能与不同种子的基因型差异有关,也有可能与辐射处理种子数有关。
牛至生物产量的主要相关性状包括株高、分枝数、茎节间长度和叶片大小等。由于牛至茎秆的精油含量低,叶片和花中的精油含量高,因此筛选大叶、茎节短(叶片量相对更大)、植株高大、分枝多以及开花量大的突变株系是的牛至主要育种目标。通过辐射诱变获得具有上述表型的牛至突变株,为培育高生物产量和高精油含量的牛至品种奠定基础。但该研究结果仅为当年数据,虽然该数据能较准确地反映变异株的典型特征,但还需多点、多年研究进一步明确变异特征。具有优异特征的牛至突变株(如大叶突变株)可以通过营养体扦插扩繁,现有结果表明上述突变性状均能在扦插后代中稳定保持。因此,上述部分突变体不仅能够为牛至杂交选育提供母本材料,而且具有作为新品种推广应用的潜力。
4 结论
60Co-γ 射线辐照希腊牛至种子可以有效获得突变群体。牛至种子小,在较低辐射剂量下(100~200 Gy)能够产生具有优异特征的突变体。通过扦插繁殖突变体单株或单个分枝,能够快速稳定突变株的优异性状。此方法简单易行,创建的突变体库为牛至新品种培育奠定了基础。
猜你喜欢
作物学报(2022年2期)2022-11-06
中国现代医生(2022年21期)2022-08-22
种子(2022年6期)2022-08-03
作物学报(2022年8期)2022-05-29
作物学报(2022年6期)2022-04-08
农村科学实验(2022年2期)2022-03-12
上海师范大学学报·自然科学版(2020年5期)2020-12-18
江苏农业科学(2020年14期)2020-08-28
三农资讯半月报(2020年2期)2020-03-09
南方农业学报(2018年9期)2018-09-10
