
牛优势卵泡膜细胞黄体化相关基因的筛选
2022-04-01 11:28孟金柱伍春亚潘春威安清明吴震洋赵园园
西北农林科技大学学报(自然科学版) 2022年3期
孟金柱,伍春亚,潘春威,安清明,吴震洋,赵园园
(1 铜仁学院 贵州省梵净山地区生物多样性保护与利用重点实验室,贵州 铜仁 554300;2 湖南农业大学 动物医学院,湖南 长沙 410128)
卵巢是雌性动物的生殖器官,其通过卵母细胞(oocytes)与颗粒细胞(granulosa cells,GCs)、膜细胞(theca cells,TCs)、基质细胞(stromal cells,SCs)之间的相互作用来调控卵泡的发育及排卵[1]。GCs中表达的细胞色素P450家族19亚家族A成员1(cytochrome P450 family 19 subfamily A member 1,CYP19A1)可将TCs分泌的雄激素转化为雌二醇(estradiol,E2),抑制下丘脑中促性腺激素释放激素(gonadotropin-releasing hormone,GnRH)的产生,从而触发促黄体素(luteinizing hormone,LH)的激增,引发排卵[2]。此时,卵泡壁上的GCs和TCs分别分化成为大黄体细胞(large luteal cells,LLCs)和小黄体细胞(small luteal cells,SLCs),LLCs直径大于25 μm,SLCs直径为12~25 μm[3]。SLCs对LH的强烈响应使其分泌孕酮(progesterone,P4)的能力得以增强,而LLCs分泌孕激素的能力是SLCs的20倍[4-5]。然而,目前关于LLCs、SLCs的起源和谱系问题还不清楚。
Hatzirodos等[6]对牛直径3~5 mm和9 mm以上卵泡的GCs和TCs进行转录组测序,发现MGARP、GLDC、CHST8、GPX3是GCs新的潜在标记物。Ticianelli等[7]对热应激下不同品种牛的卵泡卵母细胞和卵丘细胞进行转录组测序后发现,品种对卵母细胞的凋亡有显著影响,品种和温度对卵丘细胞(cumulus cells,CCs)凋亡基因的表达有影响。Christenson等[8]对牛卵泡出现LH和前列腺素(prostaglandin,PG)峰时GCs与TCs的形态变化进行了阐释,但关于卵泡到黄体的转变调控机制尚不清楚。
思南黄牛是贵州省重要的黄牛品种,身躯强健,适应性强。近几十年来,由于外来品种的不断引进及杂交导致纯种思南黄牛品种严重退化,数量急剧减少。本研究拟筛选GEO数据库GSE83524基因表达芯片中SLCs和TCs之间的差异表达基因,利用生物信息学技术对其功能进行分析,筛选出与TCs黄体化相关的关键基因,并以思南黄牛为对象加以验证,旨在为深入研究细胞特异性及TCs向SLCs分化的机制提供参考。
1 材料与方法
1.1 主要试剂
RNAiso Plus,Invitrogen公司产品;琼脂糖、TBE,均为北京索莱宝科技有限公司产品;EasyScript®One-Step gDNA Removal and cDNA Synthesis SuperMix(含Oligo(dT)primer、gDNA remover)、TransStart®Tip Green qPCR SuperMix,均为北京全式金生物技术有限公司产品。
1.2 SLCs和TCs中差异表达基因的筛选与分析
1.2.1 芯片数据的获取 本研究所用到的RNA-seq表达谱数据来源于NCBI中GEO数据库(http:www.ncbi.nlm.nih.gov/geo)。对牛卵泡发育相关的数据进行检索,获得基于GPL16500平台(Bovine Gene 1.0 ST Array)的GSE83524 数据集,其中包含了3头牛优势卵泡中的3个TCs样本和3头牛的3个 SLCs样本。
1.2.2 差异表达基因的筛选 利用R软件limma包中的DESeq2来筛选TCs和SLCs中的差异表达基因,具体筛选标准为:每1 000碱基转录每百万映射读取的片段(FPKM)≥2,|lb差异倍数|≥1,校正后P<0.01;同时使用R软件中ggplot2包绘制差异表达基因火山图。
1.2.3 GO、KEGG信号通路和PPI分析 使用R软件中的goseq包对TCs和SLCs中的差异表达基因进行GO功能富集分析,同时使用kobas软件对其进行KEGG信号通路分析;最后使用String软件构建差异表达蛋白之间的相互作用网络(PPI),并将互作数据导入Cytoscape软件,利用其自带Cyto-Hubba插件的MCC算法筛选出连通度最大的10个关键(hub)基因,进一步筛选出可能在TCs增殖分化形成SLCs过程中发挥重要作用的基因。
1.3 差异表达基因转录水平的验证
1.3.1 引物设计与合成 以筛选出的差异表达的6个基因(促卵泡素受体FSHR、磷脂酶A2-IVA基因PLA2G4A、细胞色素P450家族19亚家族A成员1基因CYP19A1、细胞色素P450家族17亚家族A成员1基因CYP17A1、苏氨酸激酶B基因BUB1B、驱动蛋白家族成员20A基因KIF20A)作为候选基因,以核糖体蛋白侧柄亚单位P0基因RPLP0为内参基因,根据GenBank中登录的牛属(Bostaurus)各基因序列,用在线NCBI软件分别设计上述候选基因和内参基因的荧光定量PCR引物(表1)。引物由生工生物工程(上海)股份有限公司合成。
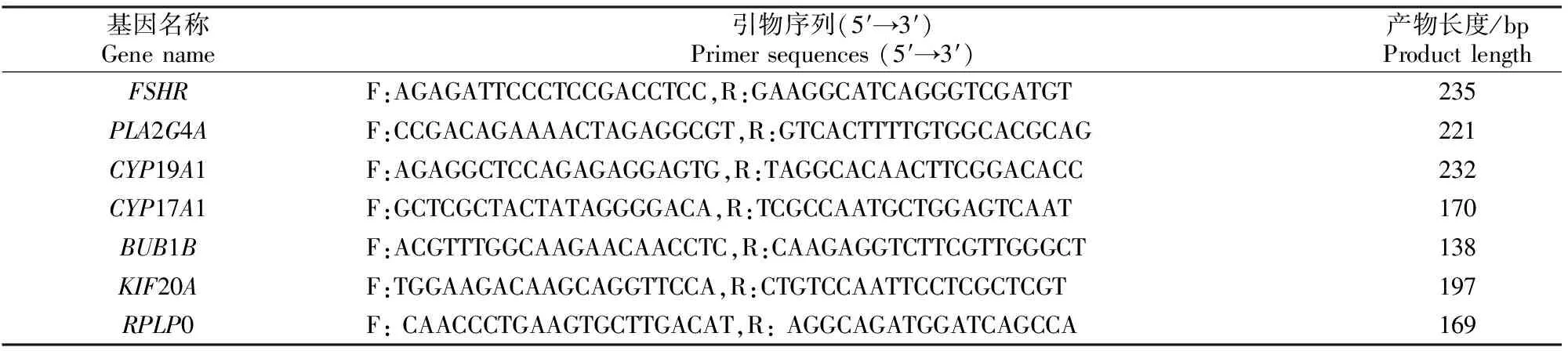
表1 试验所用引物及序列Table 1 Sequences of primers used in this study
1.3.2 候选基因在TCs和SLCs中转录水平的验证 在贵州省铜仁市万山区肉牛屠宰场,选择1岁龄健康青年思南黄牛母牛5头,屠宰后采集双侧卵巢上的最大卵泡(直径810 mm),参照本课题组前期的研究方法[9]筛选优势卵泡。将筛选出的优势卵泡置于生理盐水中,使用眼科剪刀剪开并刮去位于卵泡内膜上的GCs后,收集TCs;使用淘洗法[10]从卵巢黄体中分离出黄体细胞并经孔径约30 μm(500目)细胞筛过滤,筛选出SLCs,-80 ℃冰箱保存备用。
从-80 ℃ 冰箱中取出3个样本的GCs和TCs置于冰盒上解冻,用Trizol法分别提取TCs和SLCs的总RNA,琼脂糖凝胶电泳检测总RNA的完整性并经NanoDrop 2000测量浓度合格后,通过EasyScript®One-Step gDNA Removal and cDNA Synthesis SuperMix将TCs和SLCs的总RNA分别反转录为cDNA,同时利用gDNA Remover 去除其中的gDNA。
分别以TCs和SLCs的cDNA为模板,利用表1中的引物,经TransStart®Tip Green qPCR SuperMix通过LightCycler 480平台进行荧光定量PCR。试验设样本重复(n=3)和技术重复(n=5),建立20 μL PCR反应体系:模板cDNA 4 μL,上、下游引物各0.5 μL, SYBR MIX 10 μL,ddH2O 5 μL;PCR反应程序设定为: 95 ℃预变性30 s;95 ℃ 变性15 s,60 ℃ 60 s,72 ℃ 15 s,40个循环。
1.3.3 统计分析 各基因的相对表达量采用2-ΔΔCt来计算,并使用SPSS 17.0软件进行差异显著性分析,结果用“均值±标准误”表示。
2 结果与分析
2.1 差异表达基因的筛选
DESeq2分析后,共获得237个差异表达基因,其中上调表达基因115个,下调表达基因122个;差异倍数较高的前25个基因见表2。

表2 牛SLCs与TCs差异表达倍数较高的25个基因及其功能分析Table 2 Functional analysis of the top 25 differentially expressed genes between SLCs and TCS of cattle
由表2可见,25个差异表达倍数较高的基因分别为细胞色素P450家族19亚家族A成员1基因CYP19A1、纤维蛋白原γ链(fibrinogen gamma chain)基因FGG、细胞色素P450家族17亚家族A成员1基因CYP17A1、乳腺癌雌激素调控蛋白1(growth regulation by estrogen in breast cancer 1)基因GREB1、烯酰ACP还原酶(enoyl-ACP reductase)基因INHA、钙粘蛋白12(cadherin-12)基因CDH12、共济失调蛋白3(ataxin-3)基因ATXN3、穿透素3(pentraxin 3)基因PTX3、合子运动缺陷蛋白(ookinete motility deficient)基因OMD、生长激素抑制素(somatostatin)基因SST、丝氨酸蛋白酶抑制剂E家族成员2(serpin family E member 2)基因SERPINE2、骨甘氨酸(osteoglycin)基因OGN、原蛋白转化酶枯草杆菌素1型(proprotein convertase subtilisin type 1)基因PCSK1、无孢蛋白(asporin)基因ASPN、未知功能基因LOC506828、糖蛋白非转移性黑色素瘤蛋白B(glycoprotein non-metastatic melanoma protein B)基因GPNMB、卵泡抑素(follistatin)基因FST、脂蛋白受体相关蛋白8(lipoprotein receptor-related protein 8)基因LRP8、碳酸酐酶(carbonic anhydrase 13)基因CA13、多能蛋白聚糖(versican)基因VCAN、Rho家族GTP酶1(Rho family GTPase 1)基因RND1、谷氨酸受体辅助蛋白3(glutamate receptor auxiliary protein)基因CNIH3、促卵泡素受体(follicle stimulating hormone receptor)基因FSHR、胶原Ⅻ型α1链(collagen type Ⅻ alpha1 chain)基因COL12A1、丝裂原活化蛋白激酶8(mitogen-activated protein kinase 8)基因MAP3K8。该结果显示,部分基因已经明确与卵巢中激素的合成、应答及释放直接相关。
2.2 差异表达基因GO功能富集分析
通过goseq软件对237个差异表达基因进行GO功能分析,结果(表3)显示,参与生物学过程的有147个基因,占差异表达基因总数的53.6%,分别富集于30个组,其中负调控RNA聚合酶Ⅱ启动子的转录、细胞增殖调节、细胞粘附以及负调控细胞增殖组富集的基因最多;参与细胞组分的有125个基因,占差异表达基因总数的21.4%,分别富集于12个组,其中细胞外外泌体、细胞外空间、细胞外基质及蛋白质的细胞外基质组富集的基因最多;参与分子功能的有98个基因,占差异表达基因总数的25.0%,分别富集于14个组,其中转录激活因子活性、钙离子结合和生长因子活性组富集的基因较多。

表3 牛SLCs与TCs中差异表达基因的GO分析Table 3 GO analysis of differentially expressed genes between SLCs and TCs of cattle

表3(续) Continued table 3
2.3 差异表达基因KEGG信号通路分析
KEGG信号通路分析结果(图1)显示,共获得17条通路,其中HTLV-I感染通路富集的基因最多。在17条通路中,可能与卵泡内膜细胞黄体化相关的通路为卵巢类固醇生成通路,其中包含FSHR和磷脂酶A2-IVA(phospholipase A2-IVA)基因PLA2G4A、CYP19A1、CYP17A1等4个基因,表明这些基因在卵巢类固醇激素的合成及释放过程中起着至关重要的作用。

图1 牛 SLCs与TCs中差异表达基因的KEGG信号通路分析Fig.1 KEGG pathway analysis of differentially expressed genes between SLCs and TCs of cattle
2.4 差异表达基因PPI网络互作分析
使用String软件对差异表达基因之间的相互作用情况进行预测,同时将得到的互作数据导入Cytoscape软件进行可视化分析,筛选连通度最大的前10位关键基因。结果(图2)表明,苏氨酸激酶B(threonine kinase B)基因BUB1B、驱动蛋白家族成员20A(kinesin family member 20A)基因KIF20A的连通度最高,最具代表性;其次分别为中心体相关蛋白E(centrosome-associated protein E)基因CENPE、驱动蛋白家族成员11(kinesin family member 11)基因KIF11、核分裂周期80蛋白(nuclear division cycle 80)基因NDC80、细胞周期素A2(cyclin A2)基因CCNA2、细胞分裂周期蛋白20(cell-division cycle protein 20)基因CDC20、泛素E2连接酶C(ubiquitin E2 ligase C)基因UBE2C、MK167及爪蟾驱动蛋白2的靶蛋白(target protein for xenopus kinesin protein 2)基因TPX2等,这些关键基因间存在显著相互作用。
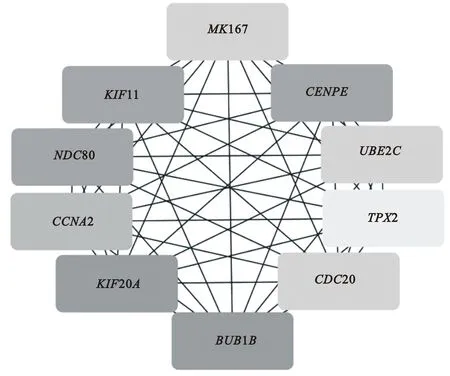
图2 牛SLCs与TCs中hub基因的PPI网络互作分析Fig.2 PPI network interaction analysis of hub genes between SLCs and TCs of cattle
2.5 差异表达基因转录水平的验证
选取卵巢类固醇生成通路中的4个差异表达基因(FSHR、PLA2G4A、CYP19A1、CYP17A1)及连通度最大的2个hub基因(BUB1B、KIF20A)进行荧光定量 PCR验证分析。结果(图3)表明,FSHR、PLA2G4A、CYP19A1、CYP17A1、BUB1B及KIF20A在SLCs和TCs中的表达趋势与GEO芯片数据结果一致,表现为FSHR、CYP19A1和CYP17A1在TCs中的表达量极显著高于SLCs (P<0.01),PLA2G4A、BUB1B及KIF20A在TCs中的表达量显著高于SLCs (P<0.05)。

图柱上标不同小写字母表示同一基因在不同细胞中的表达水平差异显著(P<0.05),标不同大写字母表示差异极显著(P<0.01)Different lowercase letters indicate significant differences in expression levels of same gene in different types of cells (P<0.05),and different capital letters indicate extremely significant difference (P<0.01)图3 差异表达基因在牛SLCs与TCs中的转录水平Fig.3 Transcriptional levels of differentially expressed genes in SLCs and TCs of cattle
3 讨 论
卵泡膜是由内膜和外膜组成的包裹在颗粒细胞周围的一层结缔组织,内膜上含有内分泌细胞,外膜是由成纤维细胞衍生而来的结缔组织层[11]。卵泡的膜层不仅能够维持卵泡结构的完整性、合成关键的内分泌调节因子,而且还可以向颗粒细胞层、卵丘细胞和卵母细胞运送营养物质[12]。在大多数哺乳动物中,卵泡TCs和GCs都参与黄体的形成,并维持不同的功能和形态[13]。灵长类动物的膜-黄体细胞形态明显,位于黄体的外边缘;而颗粒-黄体细胞则位于中央[14]。膜-黄体细胞生产雄激素,而颗粒-黄体细胞则是黄体酮和雌二醇的主要来源[15]。
研究发现,类固醇样原急性调节蛋白(steroidogenic acute regulatory protein,STAR)可以将胆固醇运输到线粒体,以调节雄激素的分泌[16]。而负责分泌雄激素的TCs所表达的CYP11A1、CYP17A1和羟基-5-甾体脱氢酶,3-和甾体-异构酶2(hydroxy-delta-5-steroid dehydrogenase,3 beta- and steroid delta-isomerase 2,HSD3B2)均为雄激素生物合成的关键酶[17]。CYP17A1是催化孕烯酮转化为17-羟基孕烯酮和孕酮转化为17-羟基孕酮的调节剂,是雄激素生物合成的限速步骤[18]。CYP17主要表达于肾上腺、睾丸间质细胞和卵巢膜细胞,这种酶的活性增加能够显著促进TCs中雄激素的生物合成和分泌[19]。体外研究表明,雄激素分泌的增加是由LH以剂量依赖性的方式驱动的[20-21],这是TCs的一个关键特征。在促黄体素受体(luteinizing hormone receptor,LHR)不变的情况下,胰岛素受体(insulin receptor,IR)的缺失促进了CYP17A1和CYP11A1的表达,从而导致雄激素过剩、发情周期紊乱甚至丧失生育能力[22]。本研究对GEO数据库GSE83524 基因表达芯片数据进行分析,共筛选出6个可能与TCs黄体化相关的基因。荧光定量 PCR验证分析结果显示,FSHR、PLA2G4A、CYP19A1、CYP17A1、BUB1B及KIF20A在SLCs和TCs中的表达趋势与GEO芯片数据结果一致。
类固醇激素通过调控下丘脑分泌GnRH或者直接作用于垂体前叶促性腺激素分泌细胞,从而影响LH和促卵泡素(follicle stimulating hormone,FSH)的产生[23]。FSH与其同源受体相互作用诱导CYP19A1的表达,将来自TCs的雄激素转化为E2,从而驱动卵泡发育,导致排卵[24]。此外,E2继续作用于GCs,促进其合成FSHR和LHR,使CYP19A1活性增强,进一步促进E2的分泌,使卵泡迅速生长[25],敲除小鼠CYP19A1,则会导致小鼠排卵[26]。骨形态发生蛋白15(bone morphogenetic protein 15,BMP15)在卵母细胞中特异表达并影响卵泡发育,BMP15的过表达会抑制GCs中FSHR的表达从而导致卵泡闭锁[27]。本研究通过Real time PCR研究发现,FSHR、CYP19A1和CYP17A1在TCs中的表达量极显著高于SLCs (P<0.01),表明其在TCs中发挥了重要作用,可能会抑制卵泡增殖分化形成黄体。
PLA2G4A是一个被排卵前促黄体素/人绒毛膜促性腺激素(luteinizing hormone/human chorionic gonadotrophin,LH/hCG)峰刺激诱导的潜在的候选基因,LH/hCG峰诱导牛卵泡排卵的过程要早于PLA2G4A的诱导[28]。注射hCG后,GCs中PLA2G4AmRNA的表达水平显著增加[29]。Lussier等[30]通过对优势卵泡和排卵卵泡GCs进行转录组测序发现,PLA2G4AmRNA在排卵卵泡GCs中的含量高于优势卵泡,可能其在促进排卵方面发挥着重要作用。BUB1B是催化蛋白质磷酸化过程的酶,能够在卵母细胞中被细胞质聚腺苷酸化元素结合蛋白(cytoplasmicpolyadenylation element-binding protein,CPEB)特异性结合,从而调控聚腺苷酸诱导的翻译[31]。在CPEB敲除小鼠中,由于突触复合体蛋白mRNA的翻译受到抑制,粗线处减数分裂进程被中断[32]。KIF20A可以促进癌细胞的增殖和转移[33]。KIF20A高表达的膀胱癌患者肿瘤分化较慢,预后较差,因此其可能成为影响膀胱癌患者预后的独立因素和膀胱癌的治疗靶点[34],关于其在动物繁殖中的作用尚未见报道。PPI网络互作分析表明,BUB1B和KIF20A为连通度最大的2个hub基因,表明这2个基因与牛优势卵泡TCs增殖、分化形成SLCs关系密切。综上,本研究结果为深入探讨TCs和SLCs的特异性及TCs向SLCs的分化机制提供了参考依据。
猜你喜欢
中国现代医生(2022年19期)2022-11-04
保健与生活(2022年12期)2022-06-09
昆明医科大学学报(2022年2期)2022-03-29
现代仪器与医疗(2021年6期)2022-01-18
人人健康(2021年21期)2021-12-09
现代临床医学(2021年4期)2021-07-31
云南医药(2021年3期)2021-07-21
天津医科大学学报(2021年1期)2021-01-26
保健与生活(2020年18期)2020-09-26
养生保健指南(2017年12期)2017-12-07
