
大豆GmCYP85A2基因克隆与表达特性分析
2022-04-01 11:26刘盼盼孟肖冰
西北农林科技大学学报(自然科学版) 2022年3期
刘盼盼,孟肖冰,付 娆,张 尧,常 玮
(南阳师范学院 生命科学与农业工程学院,河南 南阳 473061)
大豆(Glycinemax)是植物蛋白和油脂的主要来源之一,也是生产生物柴油的重要原料。随着人类消费和工业用料的快速增长,对大豆的需求也大幅增加,并将持续增长[1]。因此,提高产量潜力已成为大豆育种的长期目标。大豆的产量潜力可以通过创造理想株型来提高,而大豆的理想株型包括株高适宜、节间距短、节数较多、分枝数少、每节荚数适中、结荚率高、四粒荚较多、百粒质量适中、叶柄角小和叶柄长度短等特征[2]。其中叶柄角(leaf petiole angle,LPA),即叶柄和主茎之间的倾角,是决定大豆和许多其他豆科植物株型的关键性状。因此,挖掘与大豆叶柄角相关的基因并在此基础上解析其调控机制,对改良大豆株型结构,进而提高产量具有重要意义。
目前,有关大豆LPA遗传机制的研究鲜有报道。在基因定位研究方面,王存虎等[3]在大豆10条染色体上检测到14个与LPA相关的QTL,其中Chr12上存在一个包含5个位点的QTL簇;王吴彬等[4]以栽培大豆为遗传背景的野生大豆染色体片段代换系群体(SojaCSSLP1)为材料,检测到5个LPA相关野生等位变异,其中Sat_286位点最高能解释22%的表型变异。在功能基因研究方面,Gao等[5]利用辐射诱变获得1个LPA增大的突变体,并挖掘到一个编码APC8样蛋白的基因GmILPA1,该基因突变后可使叶枕发育异常、叶柄角增大;进一步研究表明,GmILPA1在不同叶夹角品种间表现出不同的表达水平,且表达水平与叶夹角的大小相关。Chen等[6]2020年研究发现,过表达GmMYB14会上调GmBEN1基因的表达,后者主要参与内源油菜素内脂(brassinolipid,BR)的降解代谢,最终导致过表达植株BR含量降低、叶柄角变小,株型紧凑;该研究不仅证实了MYB转录因子在大豆株型调控中的重要作用,同时也表明大豆LPA与内源BR含量有关。
BR是植物内源激素,对植物的生长发育起着重要的调控作用[7]。如水稻BR在植株营养生长和籽粒发育过程中均发挥重要作用,拟南芥BR则在其光形态建成过程中对幼苗弯钩打开和叶柄发育至关重要[8]。在水稻及玉米上的相关研究还表明,BR在叶片夹角调控中具有重要生理功能,而BR生物合成关键基因CYP则在该调控过程中发挥重要作用[9-10]。有关豆科模式植物蒺藜苜蓿的最新研究也表明,CYP基因具有调节其羽状复叶的小叶运动的功能[11]。但CYP基因在大豆LPA调控中的作用尚不清楚。为解决这一问题,本试验拟在前人研究基础上克隆大豆CYP基因,并分析其表达特性,以期为阐明大豆LPA调控分子机制及理想株型育种奠定理论基础。
1 材料与方法
1.1 供试材料及试剂
大豆品种为郑豆196,播种于河南阳师范学院科技示范农场。于V3期移植一些幼苗至光照培养箱中,培养条件为:光照12 h/黑暗12 h,温度(25±1) ℃,光照强度20 klx,相对湿度60%。大肠杆菌DH5α感受态细胞及克隆载体pEASY-T1均购于Transgen公司。
RNA提取试剂TranZOL Up、反转录试剂First-strand cDNA Synthesis SuperMix、2×EasyTaq PCR SuperMix (+dye)和Top Green qPCR SuperMix (+Dye Ⅰ),均购于Transgen公司;凝胶回收试剂盒,购于Axygen公司;高纯度质粒小量提试剂盒,购自Solarbio公司。
1.2 大豆CYP基因的同源克隆
使用TranZOL Up试剂从大豆叶柄中提取总RNA,测定浓度后作为模板,合成cDNA第一链。登录Phytozome基因组数据库(https://phytozome.jgi.doe.gov/),以玉米BRD1基因氨基酸序列作为查询序列(GRMZM2G103773),利用在线BLAST程序中的TBLASTN工具比对大豆基因组(Glycine max Wm82.a2.v1),获得相似性最高的候选基因。
根据候选基因编码蛋白(CDS)序列,利用Primer Premier 5.0设计PCR扩增引物,根据pCAMBIA3301载体的多克隆位点,分别在上、下游引物的5′端加上EcoR Ⅰ和Hind Ⅲ酶切位点。经优化,上游引物序列为:5′-TAGGGAATTCCAACACAAGAGACT-3′,下游引物序列为:5′-CTACAAGCTTTTACATCTCATCTCATT-3′,预期产物长度为1 525 bp。
以合成第一链cDNA为模板扩增候选基因的目的片段,扩增程序为:94 ℃ 5 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 90 s,35个循环;72 ℃ 10 min;4 ℃ 保存。扩增体系50 μL:2×EasyTaq PCR SuperMix (+dye) 25 μL,1×上、下游引物各0.5 μL,cDNA模板1 μL,其余用ddH2O补齐。产物经1.0%琼脂糖凝胶回收后与T载体连接并进行测序,将目标基因命名为GmCYP85A2。
1.3 大豆GmCYP85A2基因的生物信息学分析
在获得目的基因序列的基础上,采用在线工具分析候选基因编码蛋白的一级结构、分子质量及等电点(pI)等理化性质(ProtParam,https://web.expasy.org/protparam/),预测大豆候选基因编码蛋白的二级结构(SPOMA,https://npsa-prabi.ibcp.fr/cgi-bin/)、功能位点(Prosite,http://prosite. expasy.org/)、三级结构(SWISS-MODEL,https://www.swissmodel. expasy.org/)、跨膜结构(TMHMM2.0,http://www.cbs.dtu.dk/services/TMHMM-2.0/)和亚细胞定位(BaCelLo,http://gpcr2.biocomp.unibo.it/bacello/pred.htm)。以候选基因氨基酸序列作为查询序列,在NCBI上通过Blastp搜索其他物种的CYP蛋白序列并下载,利用MEGA v5.0软件中的NJ算法进行系统发育分析。利用在线软件PlantCARE (http://bioinformatics.psb.ugent.be/webtools/ plantcare/html/)对转录起始位置ATG上游2 kb的启动子序列进行分析,对候选基因的表达特性及调控模式进行初步预测。
1.4 大豆GmCYP85A2基因的表达模式分析
于R3期分别提取大豆植株的根、茎、叶片、叶柄、花、花芽和长5 mm的豆荚,提取总RNA,逆转录获得cDNA后进行实时荧光定量PCR(qRT-PCR)分析,以叶柄中的表达量为标准研究GmCYP85A2基因的组织表达情况。将V3期大豆植株移植至光照培养箱中,主茎与水平面平行放置,用量角器测量发现,靠近光源一侧的叶片会因向光运动导致叶柄角增大;分别在0(对照),4,8,12,16,20,24 h后提取叶柄RNA,逆转录后作为PCR模板,研究GmCYP85A2基因响应光照的表达特性。
以GmCYP85A2基因和大豆β-Actin基因CDS序列为模板,设计并合成GmCYP85A2基因特异引物(上游引物序列为:5′-CACAGTGGATTCCAGGCAAG-3′;下游引物序列为:5′-CAGTCACGGCCTAGTTAGTAAGATG-3′),及β-Actin内参基因特异引物(上游引物序列为:5′-GATCTACCATGTTCCCAAGT-3′;下游引物序列为:5′-ATAGAGCCACCAATCCAGAC-3′),应用ABI 7500实时荧光定量PCR仪进行扩增。扩增反应体系为:10 μL Top Green qPCR SuperMix (+Dye Ⅰ),1 μL cDNA模板,上、下游引物各0.4 μL,8.2 μL ddH2O。PCR反应程序为:94 ℃ 30 s;94 ℃ 5s,55 ℃ 10 s,72 ℃ 15 s,共 40 个循环;融解曲线分析条件为:94 ℃ 10 s,65 ℃ 34 s,94 ℃ 15 s。组织表达特性和光照响应表达特性试验中每处理均设3个样本,每样本设3个重复。用2-ΔΔCT法分析GmCYP85A2基因的相对表达情况。采用Excel 2019进行相关分析及显著性测验。
2 结果与分析
2.1 大豆GmCYP85A2基因的克隆
以合成第一链cDNA为模板,利用基因特异引物扩增大豆候选基因,PCR产物经1.0%琼脂糖凝胶电泳检测,在1 000~2 000 bp有1条特异性条带,与预期产物长度相符(图1)。回收纯化PCR产物后,经测序产物长度为1 524 bp,将该基因命名为GmCYP85A2。
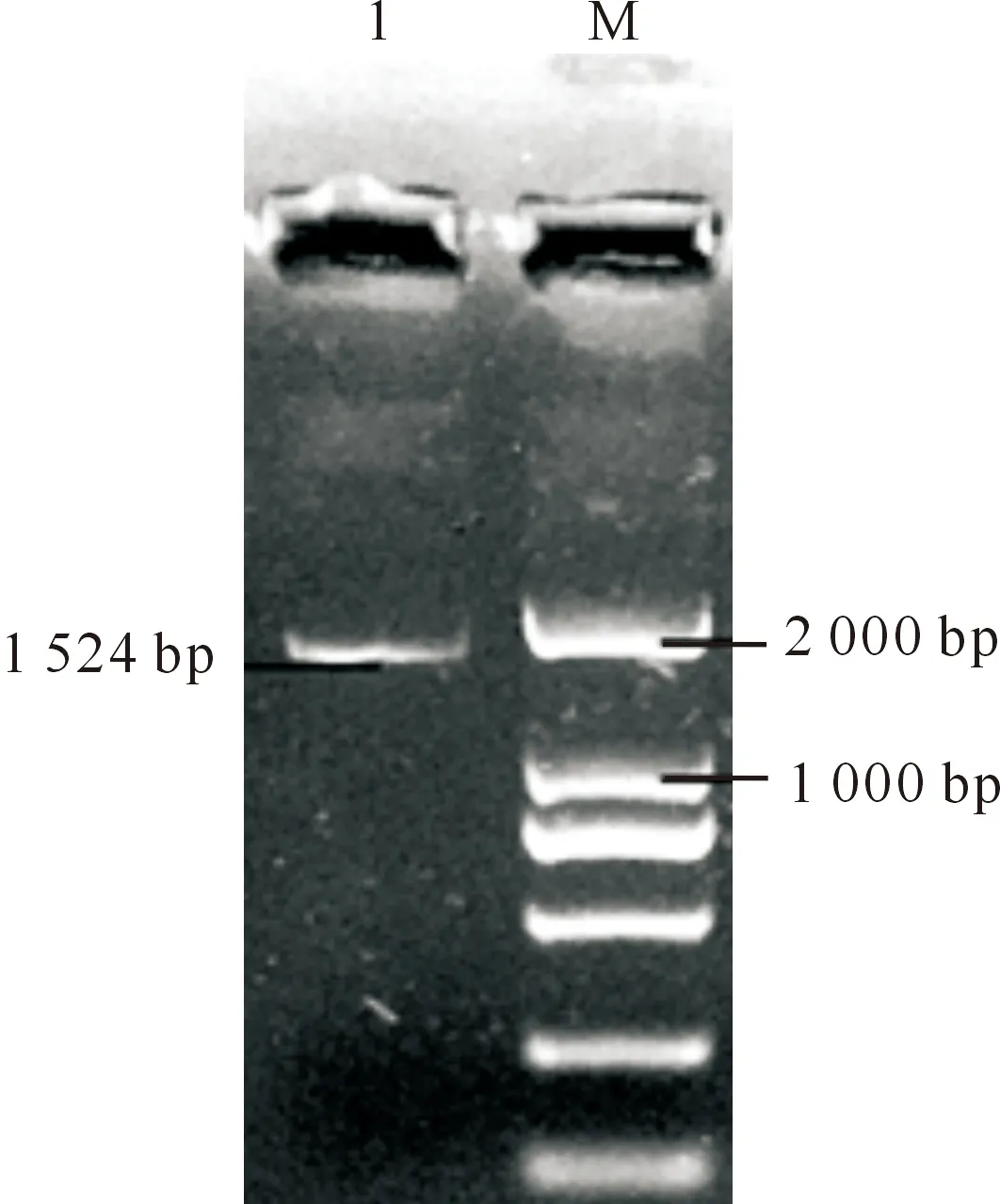
1.GmCYP85A2基因PCR产物;M.DL2000 Marker1.Production of GmCYP85A2;M.DL2000 Marker图1 大豆GmCYP85A2基因的PCR扩增结果Fig.1 PCR of GmCYP85A2 gene in soybean
2.2 大豆GmCYP85A2基因的生物信息学分析
2.2.1 测序结果 序列分析结果表明,GmCYP85A2基因开放阅读框长1 401 bp,可编码466个氨基酸。与参考基因Glyma.19G033900.1的CDS序列相比,GmCYP85A2基因在1 393 bp位置(参考基因终止密码子上游3 bp)存在1个核苷酸的缺失,从而导致该位点发生移码。与参考基因相比,GmCYP85A2基因编码蛋白序列的第465个氨基酸由酪氨酸(UAC)变为苏氨酸(ACU),且多出1个天冬酰胺(AAC)(图2)。

“*”表示一致的氨基酸,方框里面指终止密码子“*”means induced amino acid,box refers to termination codon 图2 大豆GmCYP85A2基因序列与参考序列Glyma.19G033900.1的比对Fig.2 Comparison of GmCYP85A2 gene sequence in soybean and reference sequence Glyma.19G033900.1
2.2.2 生物信息学特征 生物信息学分析结果表明,GmCYP85A2基因编码蛋白分子质量为53.4 ku,理论等电点(pI)为9.00。蛋白质二级结构α螺旋所占比例最大,为51.07%;其次为无规卷曲,约占39.91%;β折叠所占比最小,约为9.01%。利用Prosite分析GmCYP85A2蛋白的功能位点发现,该蛋白的408~417氨基酸位点具有属于P450家族的半胱氨酸血红素铁配体结构,保守序列为FGGGTRQCPG。此外,蛋白三级结构预测也表明该蛋白属于P450蛋白家族(图3)。
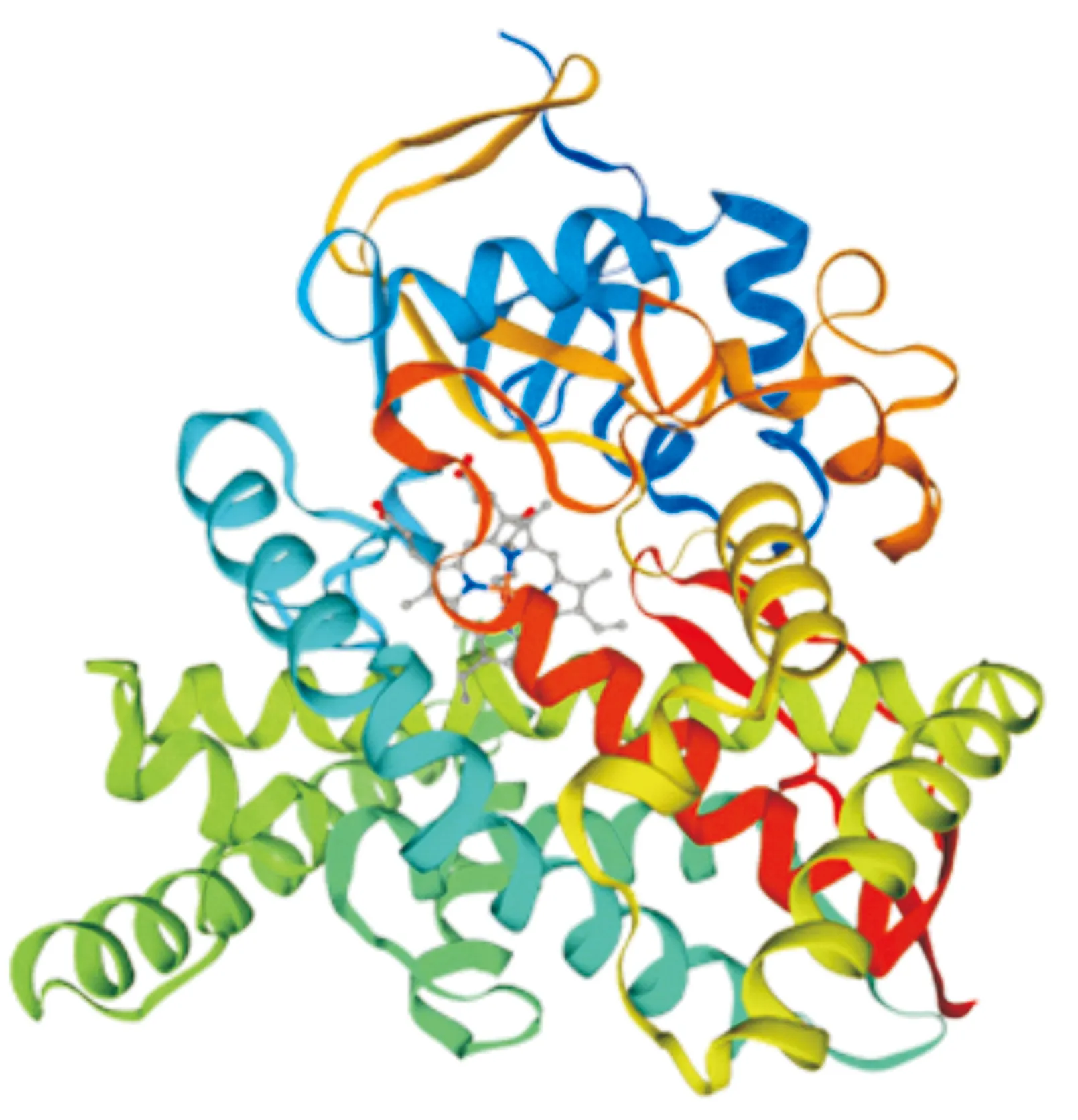
图3 大豆GmCYP85A2蛋白三级结构Fig.3 Tertiary structure of GmCYP85A2 protein in soybean
对GmCYP85A2基因编码蛋白二级结构进一步分析,该蛋白无信号肽,包含1个跨膜结构,位于该蛋白N端的3~25氨基酸位点(FLMAIVVVGVVLVLCFCSALLRW),共包含18个疏水氨基酸,且功能位点位于膜外,亚细胞定位结果显示该蛋白为分泌型蛋白(图4)。
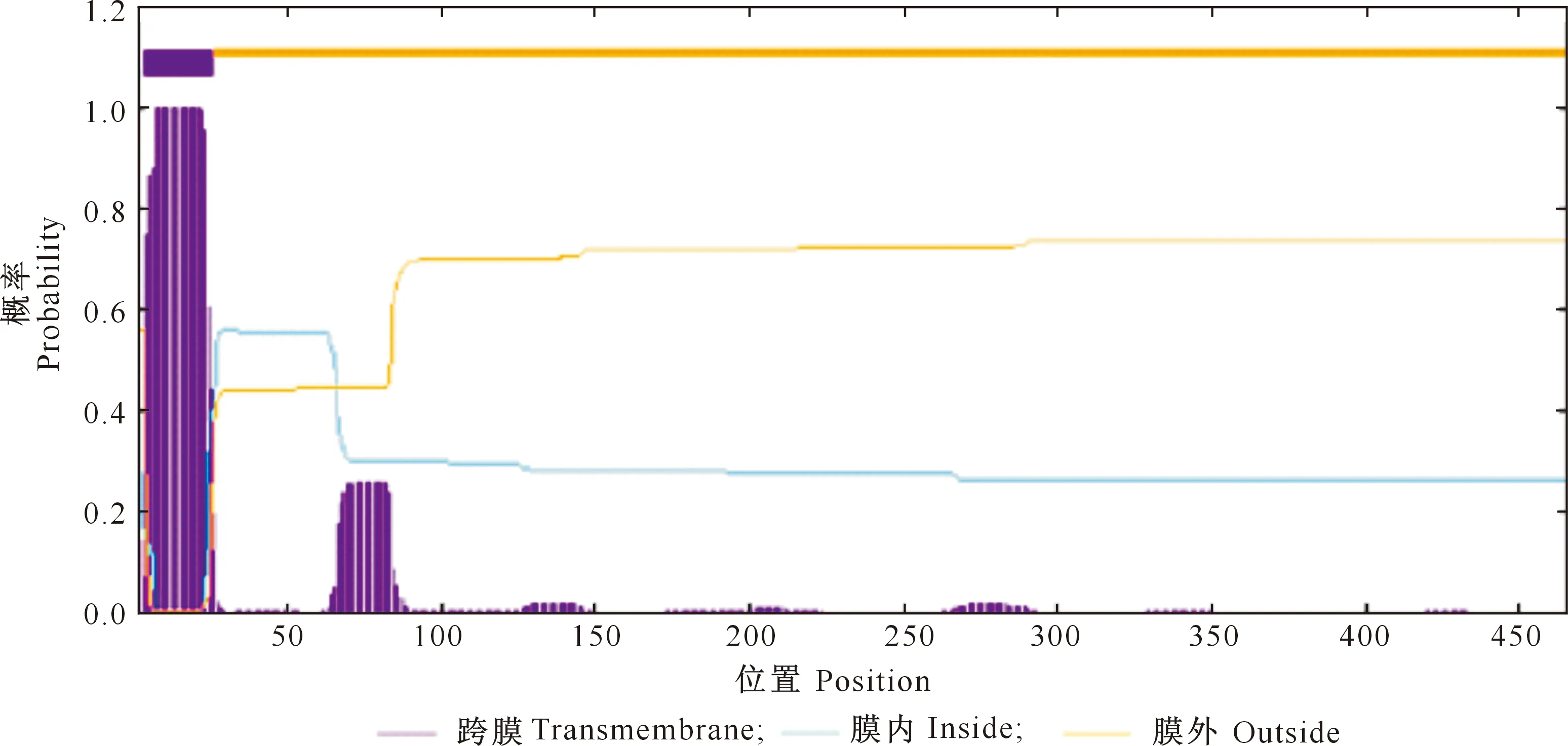
图4 大豆GmCYP85A2蛋白跨膜结构的预测Fig.4 Transmembrane structure prediction of GmCYP85A2 protein in soybean
序列比对结果显示,GmCYP85A2与野生大豆(Glycinesoja)CYP85A同源性最高,为97%。选择与GmCYP85A2相似度最高的24个物种构建系统进化分析树,结果(图5)表明,25种植物可分为4个分支,第1个分支较大,包含栽培大豆、野生大豆、木豆、鸡血藤、黧豆、绿豆、赤豆、非洲相思子、蒺藜苜蓿、鹰嘴豆和白羽扇豆等豆科植物;第2个分支包括蔷薇科的樱桃、欧洲甜樱桃、扁桃,壳斗科的板栗,大叶栎,栓皮栎,荨麻目的异色山黄麻和大麻,以及杨梅和核桃;豆科植物蔓花生、落花生、豇豆和菜豆则属于第3个分支;豆科植物角豆树则单独在一个分支。
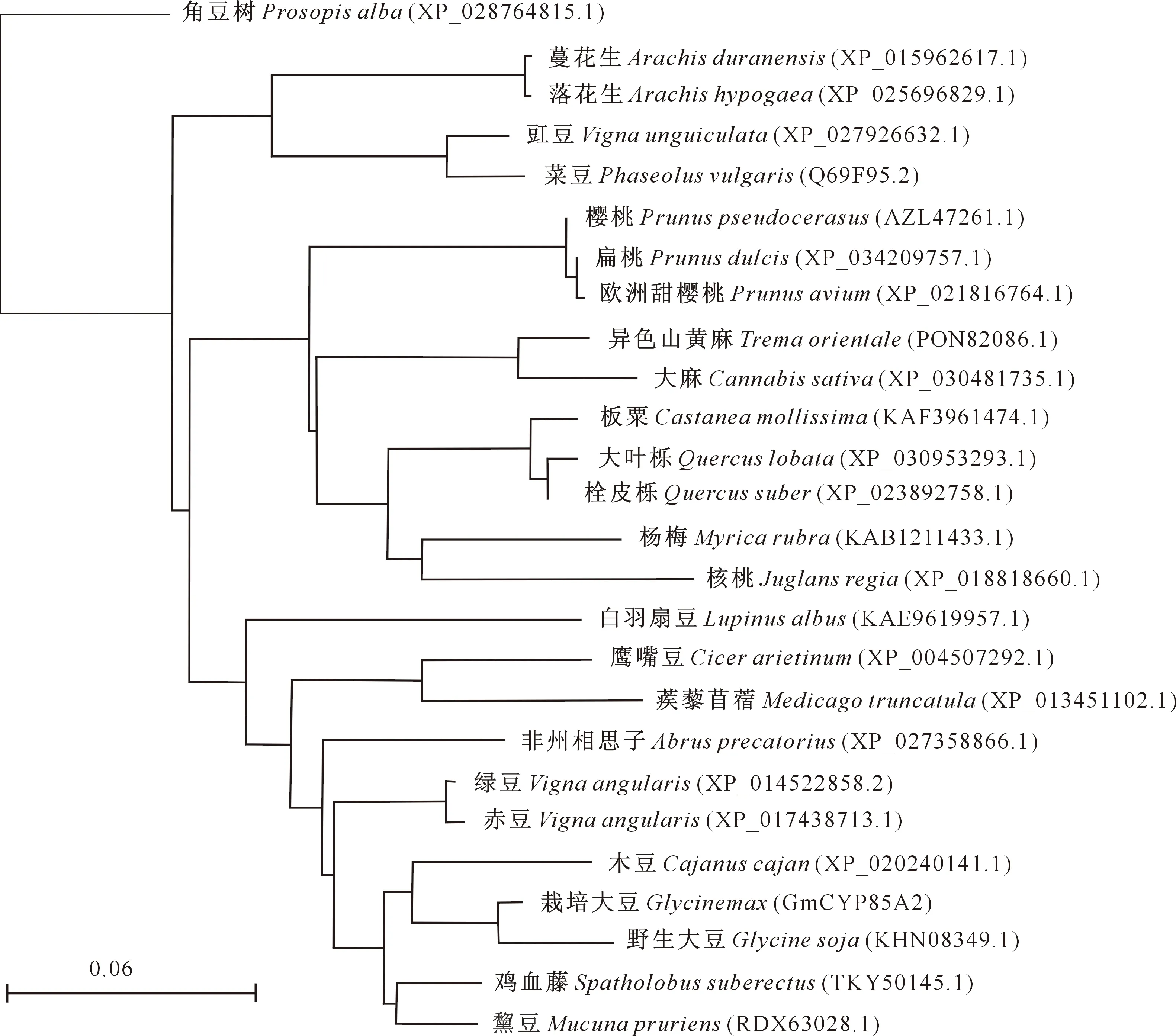
图5 大豆CYP85A2基因编码蛋白与其他植物CYP85A蛋白的系统发育树Fig.5 Phylogenetic tree of the deduced amino acid sequences of GmCYP85A2 in soybean and CYP85A of plants
2.2.3GmCYP85A2基因启动子分析 利用Pla-ntCARE对GmCYP85A2转录起始位置ATG上游2 kb的启动子序列进行分析发现,GmCYP85A2基因启动子含有多个响应光照的顺式作用元件,如GT1、ACE、G-box、GA、Box 4、ATCT、I-box等。另外还含有与生长发育和逆境胁迫相关的顺式作用元件,如GARE响应赤霉素、TCA响应水杨酸信号(可能参与植物抗病反应),而LTR则响应低温胁迫。
2.3 大豆GmCYP85A2基因的表达特性分析
2.3.1 组织表达特异性 qRT-PCR分析结果(图6)表明,GmCYP85A2基因在不同组织中表达差异较大,且以花中的相对表达量最高,约为叶柄中表达量的6.5倍;其次是花芽和豆荚,分别为叶柄表达量的2.3和1.2倍;其他组织中的表达量均低于叶柄,且以叶片中的表达量最低。表明GmCYP85A2基因可能参与大豆花及幼胚发育过程;叶片与叶柄中表达量的差异则说明,大豆发育至V期时的各项生理活动所需CYP85A2蛋白可能来自于叶柄。
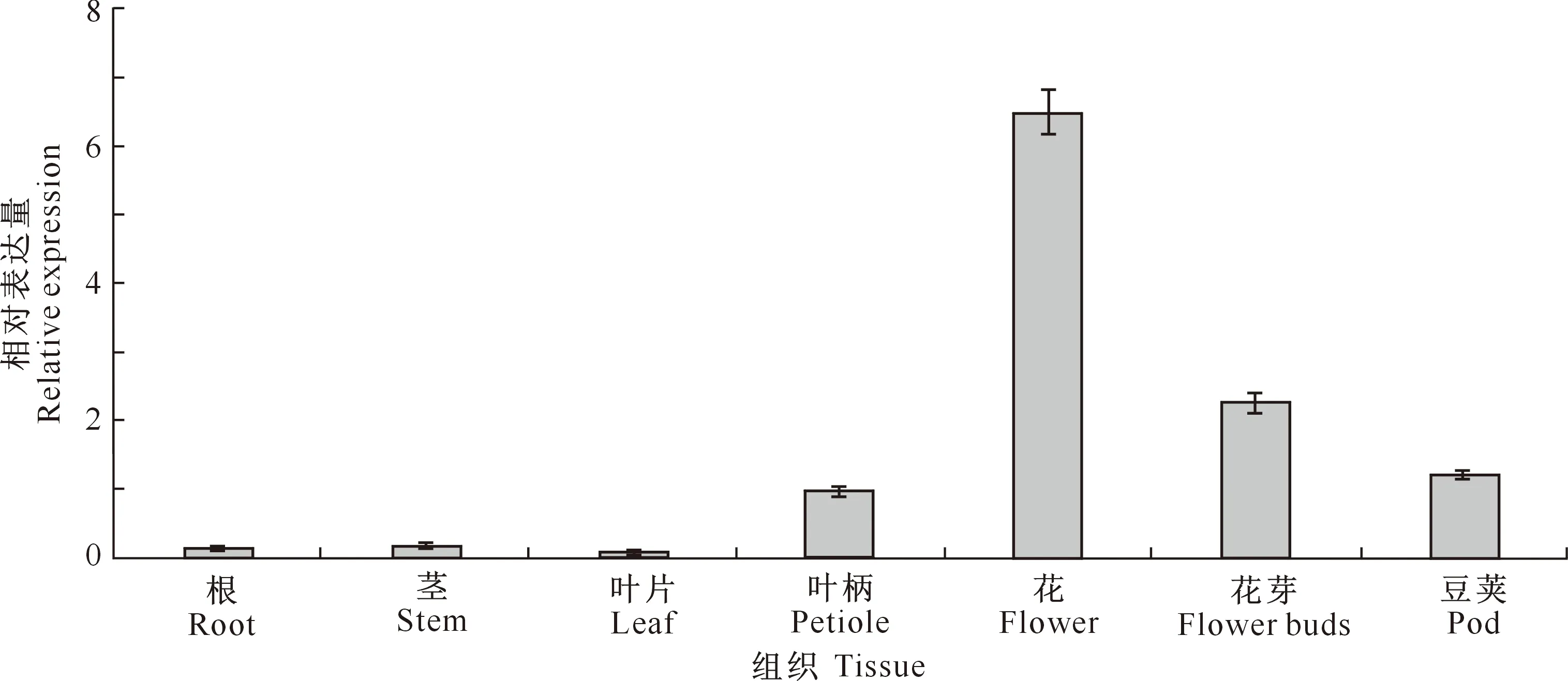
图6 大豆GmCYP85A2基因的组织表达特异性Fig.6 Tissue expression characteristics of GMCYP85A2 gene in soybean
2.3.2 光照响应特征 图7显示,将大豆主茎水平放置后,随着时间延长,靠近光源一侧的叶柄角逐渐变大,叶柄中GmCYP85A2的表达量在0~12 h呈上调趋势,到12 h时约为对照的14倍;之后开始缓慢回落,但仍然维持较高的水平;至24 h时约为对照(0 h)的9.2倍。此结果表明,光照既能诱导大豆叶柄GmCYP85A2基因的表达,也能诱导叶柄角发生改变,且某时间段(0~12 h)叶柄角与该时段起点GmCYP85A2的表达量存在正相关关系(r=0.77,0.05 图7 大豆叶柄中GmCYP85A2基因的光照响应特征Fig.7 Light response characteristics of GMCYP85A2 gene in soybean petiole 本研究得到的GmCYP85A2蛋白408~417氨基酸位点具有P450家族半胱氨酸血红素铁配体保守结构,与Xu等[12]研究结果相似。近年的研究表明,目前大多数已知的植物P450被存在于N端的疏水肽锚定在内质网(ER)的膜表面,形成跨膜片段[13]。本研究中的GmCYP85A2蛋白N端3~25氨基酸位点包含1个由18个疏水氨基酸组成的跨膜结构,且该蛋白为分泌型蛋白,均符合植物P450的结构特征。 细胞色素P450羟化酶超基因家族催化活性广泛,无论在植物基础代谢还是次级代谢过程中都具有重要作用[14]。植物细胞色素P450羟化酶数量庞大,根据氨基酸序列相似性可分为11个独立的家族[15]。仅大豆中有功能注释的基因就多达306个,但多数为生物学功能未知的基因[13]。少数几个功能明确的基因被证实在大豆的多个生理过程中发挥重要作用,如CYP81E11[16]、CYP81E22[17]分别是大豆对盐胁迫及除草剂苯达松高度敏感的关键基因,CYP86A37和CYP86A38在体内发挥脂肪酸ω-羟化酶的作用[18];双子叶植物特有的CYP82家族基因则被证实参与了茉莉酸和乙烯信号转导,进而增强了大豆对生物和非生物胁迫的抗性[19]。 本研究的系统发育分析发现,大豆GmCYP85A2基因编码蛋白与多种植物的CYP85蛋白具有较高的同源性。在植物中P450CYP85家族具有广泛的底物特异性,催化BR生物合成中多个必要的C-6氧化反应,是BR合成的关键酶[20-21],具有调控株高、叶角和籽粒大小、开花、叶片衰老、花粉发育和雄性不育、叶绿体发育、根生长和发育、光形态建成的功能[22-28],并在植物对生物和非生物胁迫的应激反应等生理过程中发挥着重要作用[29-31]。此外,CYP85基因还是植物二萜类化合物合成[32]以及皂角(GlehniaLittoralis)中呋喃香豆素(Furanocoumarin)生物合成途径下游的关键酶基因[33]。 本研究对大豆GmCYP85A2基因的组织表达模式进行了初步探索,结果表明,该基因具有组织特异性,主要在花、花芽、豆荚及叶柄等地上部分表达,与Bancos等[20]的“拟南芥CYP85A2转录本表达特性”研究结果一致。本研究中,与其他组织相比,GmCYP85A2在花中相对表达量明显上调,表明该基因可能参与大豆花发育过程的调控。在拟南芥中,BES1转录因子受BR的调控而表达,再与ELF6和REF6等转录因子互作来调节开花时间[25]。推测大豆GmCYP85A2也可能通过调控BR的生物合成发挥开花调控功能。 本研究发现,大豆GmCYP85A2基因表现出一定的光照响应表达特性,光照0~12 h叶柄中GmCYP85A2的表达量随叶柄角逐渐变大而增大,表明该基因可能参与大豆叶柄角调控;12 h后表达量有一定程度下降,表明该基因转录水平受到抑制。Bancos等[20]研究表明,在拟南芥中,CYP85A2基因不仅是BR生物合成的关键酶,同时BR也会对CYP85A2基因具有反馈调节作用,从而使该基因表达量下调。据此推测大豆GmCYP85A2基因可能是BR生物合成的关键基因,并对叶柄角有一定的调控作用,同时该基因表达会受到BR的反馈调节。 本研究还发现,GmCYP85A2基因启动子含有多个响应光照的顺式作用元件,如GT1、ACE、G-box、GA、Box 4、ATCT、I-box等;同时表达分析结果也表明,0~12 h叶柄中GmCYP85A2的表达量随着光照时间推移而上调,并伴随着叶柄角的逐渐增大,表明GmCYP85A2基因的表达受光照的调控,并对叶柄角作出调节,而该基因是否通过参与大豆BR生物合成来调控叶柄角还有待进一步研究。此外,GmCYP85A2基因启动子含有能够响应赤霉素、水杨酸信号及低温胁迫信号的顺式作用元件GARE、TCA及LTR,但该基因是否参与大豆生长发育调节以及抵抗生物和非生物逆境胁迫也有待进一步研究。 本研究采用同源克隆技术克隆大豆GmCYP85A2基因,对其编码蛋白进行了生物信息学分析,同时进行基因组织表达和光照响应表达特性分析,结果表明,GmCYP85A2基因编码蛋白具有典型的P450超家族保守结构和蛋白三级结构,该基因启动子区含有多个响应光照的顺式作用元件。R3期该基因主要在大豆花、花芽、豆荚及叶柄中表达,且以花中的表达量最大;而在V3期,叶柄中GmCYP85A2的表达量受光照影响而上调,并伴随着叶柄角的变大。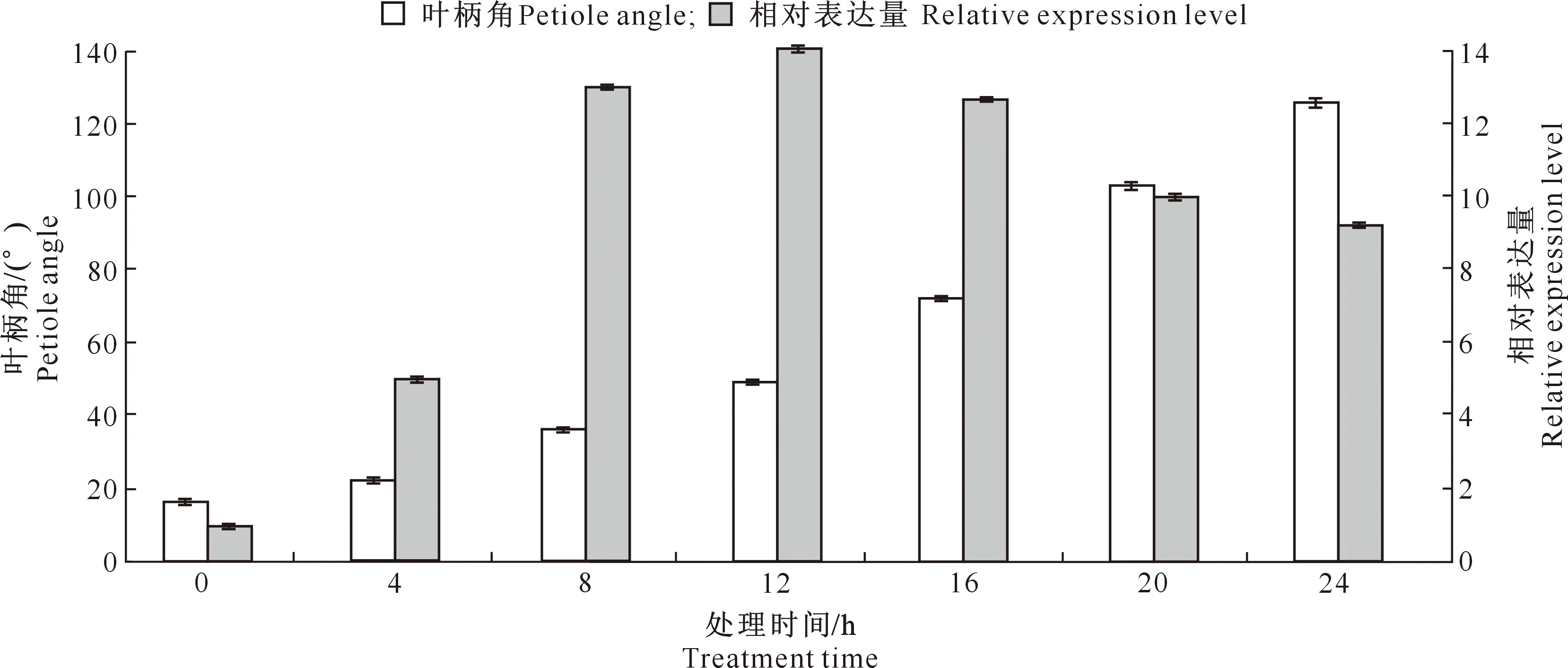
3 讨 论
4 结 论
猜你喜欢
中国动物保健(2022年10期)2022-11-04
作文·小学中高年级(2022年2期)2022-06-23
小天使·四年级语数英综合(2022年6期)2022-06-21
中国农学通报(2022年12期)2022-06-01
中国糖料(2022年2期)2022-04-06
中国种业(2021年11期)2021-11-25
作文·小学低年级(2021年4期)2021-11-02
小资CHIC!ELEGANCE(2019年40期)2019-12-10
中学生物学(2019年7期)2019-10-17
江苏农业科学(2017年3期)2017-05-02
