
干旱胁迫下新疆杨树冠不同高度叶片水分状况与非结构性碳动态
2022-04-18 08:23陈图强徐贵青刘深思米晓军
西北植物学报 2022年3期
陈图强,徐贵青*,刘深思,米晓军,李 彦
(1 中国科学院 新疆生态与地理研究所,荒漠与绿洲生态国家重点实验室,乌鲁木齐 830011;2 中国科学院 阜康荒漠生态国家野外科学观测研究站,新疆阜康 831505;3 中国科学院大学,北京 100049)
IPCC第四次评估报告预测,21世纪末全球地表平均温度将升高1.1~6.4 ℃[1]。持续升温和降水格局调整[2],预计干旱发生频率会增加,干旱化程度会加剧[3],致使全球陆地生态系统树木死亡频发[4]。三北防护林工程于1978年启动,经40多年建设,在防风固沙、改善区域气候和水土流失等方面取得显著成效[5]。杨树作为防护林区的主栽树种,占农田防护林90%以上[6],具有适应性强、生长速度快等特性,被广泛用于木材生产和生态恢复。近年来,全球变暖及地下水位下降所引发的干旱导致了不少地区的杨树出现生长衰退甚至死亡等现象[7-8],严重影响了生态防护林功能的发挥。
根据植物对外界水分环境的敏感性差异,可将植物划分为等水植物和非等水植物[9]。等水植物在外界水环境发生变化时响应迅速,在遭受干旱胁迫时,等水植物会降低气孔导度来减少水分散失[10]。但同时会影响叶片与外界的气体交换,导致蒸腾和光合速率下降。安玉艳等[11]指出,蒸腾失水与光合碳同化对干旱胁迫敏感性的差异可提高植物的水分利用效率(WUE),降低了植物碳水收支不平衡的风险。随干旱时间的延长,植物细胞忍耐水分亏缺的能力会发生变化。张中峰等[12]指出,长期干旱胁迫下,植物组织细胞的饱和膨压渗透势、膨压损失点水势会下降,抗旱能力有一定提高。此外,植物会对其形态进行调整,如叶片增厚、叶面积减小等。当水势下降超过一定阈值后,即使植物在形态或生理上进行适应性调整,木质部也将形成栓塞,阻碍水分运输[13]。
非结构性碳水化合物(NSC)是植物生长代谢过程中的重要能源物质,主要包括可溶性糖和淀粉。NSC参与植物新陈代谢、养分吸收运输、栓塞修复等生理过程[14]。植物存储的NSC在新近合成NSC不足时,可起一定的调节和缓冲作用[15]。但如果存储的NSC难以满足其消耗需求时,植物体的碳收支平衡将会被打破,长时间干旱可能会导致NSC供应不足而引发碳饥饿[16]。而干旱胁迫不仅影响碳同化过程, 也会对碳水化合物在各组织与器官间的分配格局及组分产生影响[17]。如Traversari等[18]指出,在干旱胁迫下,杨树韧皮部的淀粉会转变成可溶性糖。章异平等[15]指出,栓皮栎(Quercusvariabilis)枝条的可溶性糖与淀粉之间存在动态转化。总之,水力性状及NSC在植物抗旱中的作用不容忽视,对于认知干旱胁迫下植物生存前景和死亡发生风险至关重要。
水力性状及NSC含量在不同树冠高度间存在差异。Ryan等[19]所提出的水力限制假说阐述了树冠不同高度的水分生理差异,指出随着水分运输路径的增加,运输阻力会加剧,进而降低木质部水力导度。而Fang等[20]指出,小青杨(Populuspseudo-simonii)顶梢枯死是由于水力限制所引起。此外,还有不少国内外学者对树冠不同高度的非结构性碳水化合物、光合速率、气孔导度、水分利用效率、叶绿素含量和胡伯尔值等进行了研究[21-25],揭示了树冠垂直方向性状差异。然而干旱胁迫下,树冠不同高度水力结构调整、碳收支权衡与树木局部死亡的作用关系仍需验证。新疆杨(PopulusbolleanaLauche.)是生态防护林和城市绿化的主要树种,在防风固沙和保卫绿洲中履行着重要的生态功能。探求干旱胁迫对新疆杨树冠不同高度叶片水力性状与碳分配的影响,可以促进对树木局部死亡机制的理解,为干旱区防护林的管理及未来新疆杨在防护林营造工程中的合理布局提供指导性的建议。
1 材料和方法
1.1 研究区概况
研究区位于新疆维吾尔自治区阜康市境内新垦绿洲222团,试验依托中国科学院阜康荒漠生态系统观测站进行(后简称阜康站)。该地区属于温带荒漠气候,冬冷夏热,昼夜温差大。多年平均气温6.6 ℃,一年中最高温度和最低温度分别在7月和1月,分别为25.6 ℃和-17 ℃。年日照时数约2 143 h,年积温3 000 ℃以上。多年平均降雨量164 mm,年蒸发量2 000 mm以上, 降水主要集中在5~9月。土壤类型以风沙土、灰漠土为主[26]。除新疆杨(PopulusbolleanaLauche.)外,该地区农田防护林的树种还有沙枣树(Bryoniadioica)、毛白杨(Populustomentosa)及白榆(Ulmuspumila)等[27]。
1.2 试验设计
以阜康站东侧农田防护林带中的新疆杨为研究对象,其树龄25 a,树高约13.5 m。在防护林南北两侧各标记8棵株高、胸径基本一致的植株,于2019年5~9月和2020年7~8月进行试验。干旱处理组在试验开始后即停止浇水,对照组正常浇水。与对照组相比,干旱处理组树冠因叶片凋落而明显稀疏于对照组。在2019年及2020年生长季,分别测定黎明叶水势、正午叶水势、枝条含水量和叶片含水量;在2019年7月测定胡伯尔值和净光合速率;于2020年7~8月测定饱和膨压渗透势、膨压损失点水势、叶绿素SPAD值、比叶面积、淀粉含量和可溶性糖含量并计算NSC含量。另外,通过对0~20、20~40、40~60、60~80和80~100 cm深度土壤含水量的测定,结果表明,各土层土壤含水量在对照组和干旱处理组间均存在显著性差异(表1)。
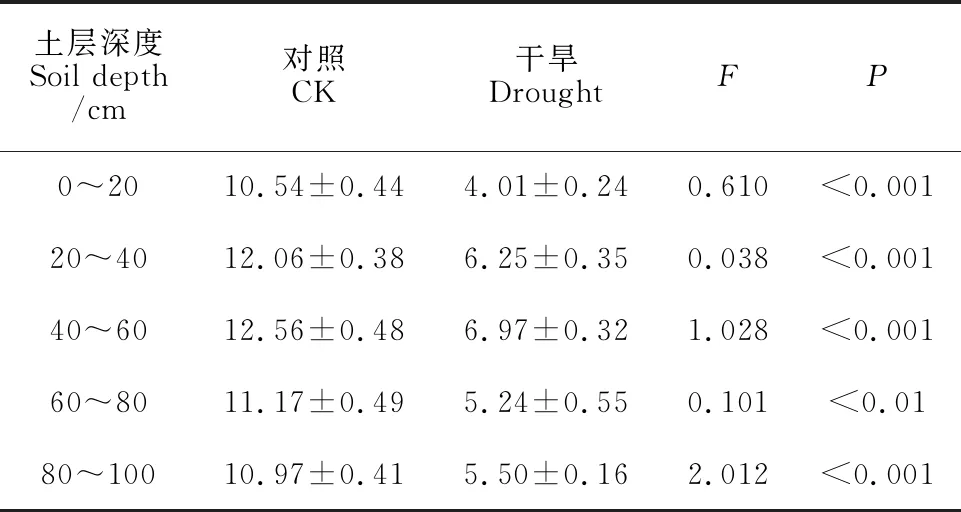
表1 不同土层深度土壤含水量(平均值±标准误,n=3~4)
1.3 试验方法
1.3.1 水分生理指标的测定(1)叶片水势。于2019年5~8月及2020年8月分别对新疆杨的叶水势进行测定。在新疆杨树冠距地面3.5 m(下部)、7 m(中部)及10.5 m(上部)处分别选择4~5条向阳且健康生长的小枝,将其装入有湿纸球的隔热箱中,待取样完成后用压力室水势仪(Model 3500, PMS Instrument Company, Albany, USA)测量枝条末端叶片的水势。选在日出前30 min测定黎明前叶水势,于12:30~14:00测定正午叶水势。
(2)叶片及枝条含水量。于2019年生长季5~9月及2020年8月采用烘干称重法测定叶片及枝条含水量。取样后立即带回实验室用万分之一天平称其鲜重(Wf),然后放入75 ℃烘箱烘干至恒重(Wd),通过以下公式计算含水量(θ):
θ=(Wf-Wd)/Wd×100%
(3)叶压力-容积曲线。在日出前30 min采取向阳且健康生长的枝条,随后立即放入蒸馏水中,并在水中剪去一小段,带回实验室将其置于黑暗环境下饱和吸水3 h以上,期间室温与外界环境基本一致。取复水完枝条上面的叶片称其饱和重,随后立即用压力室水势仪测定叶水势,所有测定叶片复水完的水势均大于-0.3 MPa。然后将其放在试验台上自然干燥一段时间后,再称其重量并测定水势(干燥初期隔1~3 min测定1次,后期每隔30~60 min测定1次),重复该过程直到叶片严重萎蔫。最后将叶片在75 ℃烘箱中烘干至恒重。用压力-容积曲线拟合程序[28]计算饱和膨压渗透势及膨压损失点水势。
1.3.2 光合生理指标的测定(1)净光合速率和水分利用效率。在晴朗无风的天气下测定新疆杨光合参数,选择3个树冠高度下健康成熟的叶片,用Li-6400便携式光合仪测定系统(Li-Cor, Lincoln, NE, USA)于11:30~13:00测定净光合速率(Pn)和蒸腾速率(Tr),使用2×3 cm2红蓝光源叶室,光强设置为1 500 μmol·m-2·s-1,二氧化碳浓度为400 μmol·mol-1,叶室温度为33 ℃。依据测定值计算瞬时水分利用效率(WUE):WUE=Pn/Tr
(2)SPAD值。测定净光合速率的同时,使用SPAD-502叶绿素含量测定仪(Konica Minolta, Japan)测定不同高度和水分处理下的8片叶相对叶绿素含量SPAD值。
1.3.3 可溶性糖和淀粉含量的测定选取对照组和干旱处理组不同树冠高度的叶、枝条、韧皮部和地表以下5~10 cm土壤深度的细根,带回实验室放入75 ℃烘箱烘干至恒重,随后用球磨仪(MM400, Retsch, Düsseldorf, Germany)将样品粉碎装入密封袋内保存待测。采用改进苯酚硫酸比色法测定可溶性糖和淀粉含量[29],NSC含量为可溶性糖和淀粉含量之和。
1.3.4 比叶面积、胡伯尔值的测定使用测定完含水量的叶和枝条,扫描叶片得叶图像(Epson Perfection 2400 Photo, Seiko Epson Corp., Japan),使用Image J(US National Institutes of Health, Bethesda, MD, USA)计算叶面积,同时测定枝条的边材面积。叶面积与叶干重之比即为比叶面积,胡伯尔值为边材面积与叶面积之比。
1.4 数据处理
使用SPSS 24.0进行数据统计分析。使用单因素方差分析(one-way ANOVA)对不同树冠高度的生理指标进行差异显著性分析,使用Duncan进行多重比较 (α=0.05)。使用独立样本t检验对不同水分处理间的指标进行显著性检验(α=0.05)。所有数据均通过正态性和方差齐性检验,对于非正态分布的数据使用非参数检验,方差不齐的指标多重比较使用Tamhane’s T2检验(α=0.05)。利用Origin 2019进行绘图。
2 结果与分析
2.1 新疆杨不同树冠高度的叶片水分状况
2.1.1 黎明叶水势2019年生长季,除第228天有过降雨外,其余时间均无降雨。从表2可知,树冠10.5 m处黎明叶水势在对照组普遍高于3.5 m处,在干旱处理组除第234天外均低于3.5 m处。除第150天外,干旱处理组10.5 m处的黎明叶水势均显著低于对照组,而其3.5 m及7 m处的黎明叶水势与对照组之间的大小关系随时间而变化。对照组生长季平均黎明叶水势在3.5 m、7 m和10.5 m处分别为-0.66、-0.68和-0.48 MPa,相应的干旱处理组分别为-0.74、-0.79和-0.86 MPa。2020年8月,干旱处理组黎明叶水势随树冠高度增加而下降,10.5 m及7 m处显著低于3.5 m处;此外,干旱处理下7 m及10.5 m处的黎明叶水势显著低于对照组(图1,Ⅰ)。
2.1.2 正午叶水势2019年生长季,对照组10.5 m处的正午叶水势除第228、234及243天外均低于3.5 m处,干旱处理组10.5 m处的正午叶水势除第195、234及243天外均显著低于3.5 m处。10.5 m处正午叶水势除第150及195天外均表现为干旱处理组小于对照组,3.5 m及7 m处的正午叶水势在对照和干旱处理组间的大小关系随时间变化。对照组生长季平均正午叶水势在3.5 m、7 m和10.5 m处分别为-1.92、-2.22和-2.07 MPa,相应的干旱处理组分别为-2.01、-1.97和-2.30 MPa。在2020年8月,对照组的正午叶水势随树高增加先降后升,而干旱处理组的正午叶水势随树冠高度的增加显著降低;干旱处理下各树冠高度的正午叶水势与相应对照组均无显著差异(图1,Ⅱ)。
2.1.3 叶和枝条含水量2019年生长季期间,除第150天外,干旱处理3.5 m处的叶含水量均显著低于同期对照组;7 m处叶含水量在第180及233天显著高于对照;10.5 m处的叶含水量在第180及215天显著低于对照组(表2)。对照组树冠10.5 m处叶含水量除第180天外均显著低于3.5 m处,而干旱处理组树冠10.5 m处叶含水量在第150、215天均显著小于3.5 m处。对照组生长季平均叶含水量在树冠3.5 m、7 m、10.5 m处分别为71.36%、66.3%和66.24%,相应的干旱处理组则分别为66.62%、68.04%和65.24%。在2020年8月,对照组各个树冠高度的叶及枝条含水量无显著差异(图1,Ⅲ、Ⅳ);干旱处理组的叶含水量随树冠高度增加而降低但差异不显著,枝条含水量随树冠高度增加而下降,且在7 m及10.5 m处显著低于3.5 m处。与对照组相比,干旱处理组10.5 m处的叶含水量显著降低,其余树冠高度则无显著性变化;各个树冠的枝条含水量在干旱处理组与对照组之间无显著差异。可见,从叶水势、叶含水量及枝条含水量来看,随着树冠高度的增加新疆杨受到的水分胁迫逐渐增大。
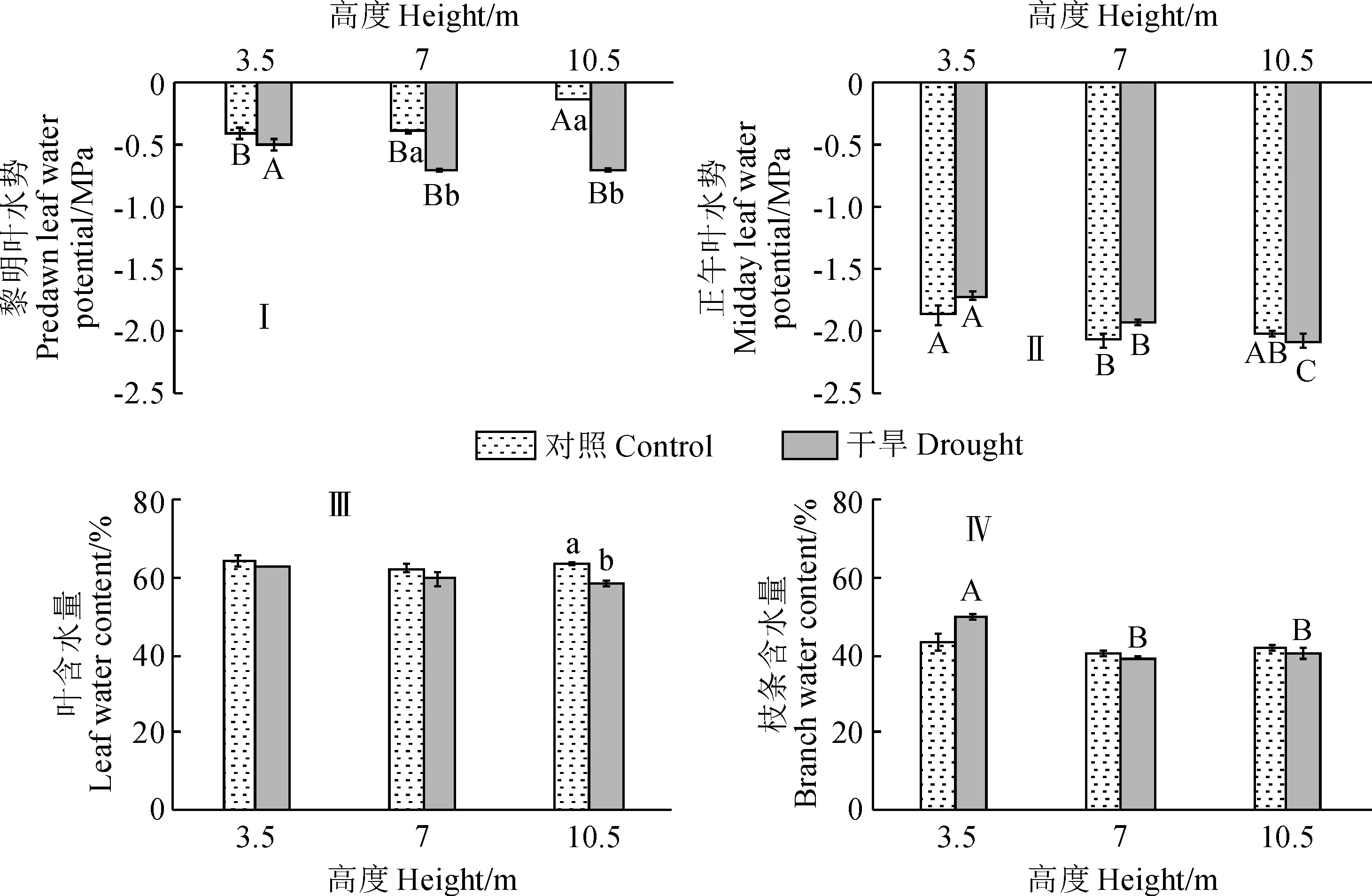
同一处理内不同大写字母表示树冠高度间存在显著性差异(P<0.05),而同一树冠高度下不同小写字母表示处理间存在显著性差异(P < 0.05)。图中数值为平均值±标准误,下同
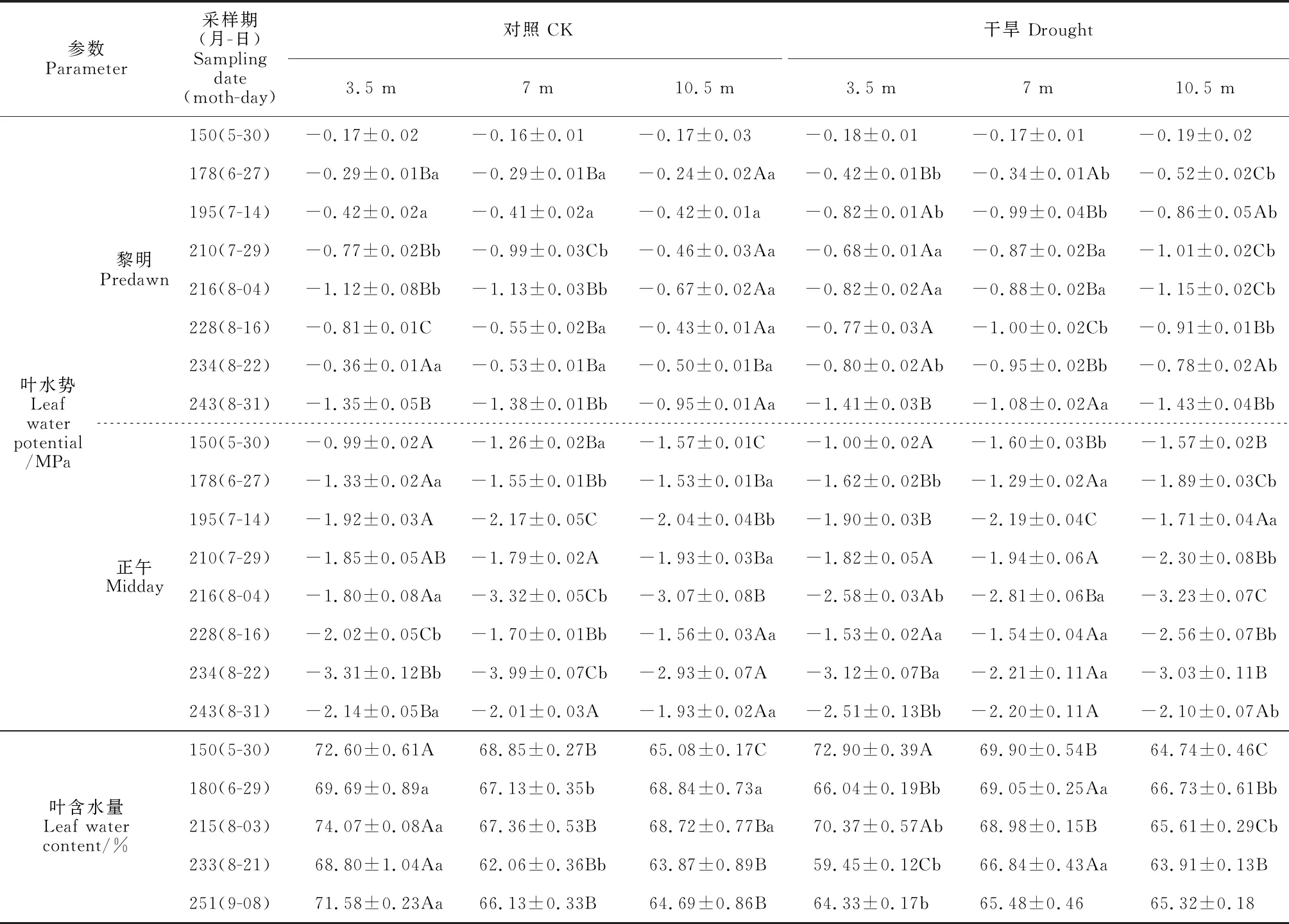
表2 2019年生长季不同处理及树冠高度下的叶水势和叶含水量
2.1.4 叶片饱和膨压渗透势和膨压损失点水势干旱处理组各树冠高度的叶片饱和膨压渗透势(图2,Ⅰ)、膨压损失点水势(图2,Ⅱ)与对照组均相比无显著差异;同时,叶片饱和膨压渗透势、膨压损失点水势在对照组及干旱处理组的各树冠高度间也均无显著性差异。
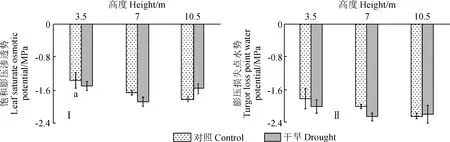
图2 两种水分处理下新疆杨不同树冠高度叶片的饱和膨压渗透势和膨压损失点水势(n=4)
2.2 新疆杨不同树冠高度叶片的光合生理特征
在不同试验处理及树冠高度下新疆杨叶片光合生理响应有所差异。其中,对照组叶片净光合速率(图3,Ⅰ)随树冠高度增加呈下降趋势,且在10.5 m处显著低于3.5 m处;水分利用效率(图3,Ⅱ)在各树冠高度间无显著差异;叶绿素SPAD值(图3,Ⅲ)在7 m及10.5 m处显著低于3.5 m处。在干旱处理下,叶片净光合速率随着树冠高度增加也呈下降趋势,且10.5 m处显著低于3.5 m及7 m处;水分利用效率表现为树冠上部高于下部,但无显著差异;叶绿素SPAD值在10.5 m处显著低于3.5 m处。与对照组相比,干旱处理组叶片的净光合速率在7 m处显著增加,各树冠高度水分利用效率均显著提高,叶绿素SPAD值在3.5 m及10.5 m处显著降低。
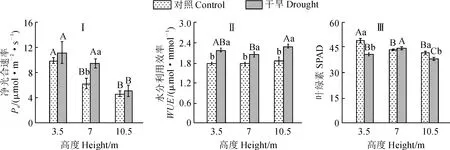
图3 两种水分处理下新疆杨不同树冠高度叶片的光合参数比较(平均值±标准误,n=5~8)
2.3 新疆杨不同树冠高度的非结构性碳水化合物含量比较
在不同水分处理及树冠高度下新疆杨叶、枝条及韧皮部的可溶性糖、淀粉及非结构性碳水化合物(NSC)含量的变化趋势有所差异(图4)。首先,对叶片而言,对照组和干旱处理组叶片的可溶性糖及NSC含量随树冠高度的增加都无太大变化,均未达到显著水平;干旱处理下从树冠下部到上部叶片的可溶性糖含量占NSC总含量的85.8%~90.2%;干旱处理组叶片的淀粉含量在7 m及10.5 m处要显著高于相应对照组,其NSC含量在7 m处要显著高于相应对照组。其次,对于枝条而言,对照组枝条的可溶性糖含量在各树冠高度间无显著差异,而淀粉及NSC含量随着树高先增加后减少,表现为7 m>3.5 m>10.5 m,且各树冠高度间均存在显著差异;干旱处理组的枝条可溶性糖、淀粉、NSC含量随高度增加基本呈现上升趋势,且在10.5 m处显著高于3.5 m及7 m处;与对照组相比,干旱处理组可溶性糖、淀粉及NSC含量在3.5 m及7 m处均要显著降低,而在10.5 m处均显著升高。再次,对于韧皮部而言,对照组可溶性糖含量在10.5 m处显著高于其他高度,淀粉及NSC含量随高度增加呈先增后降的趋势,并均在7 m处最高,且与其他树冠高度差异显著;干旱处理组韧皮部可溶性糖、淀粉及NSC含量在各树冠高度间均无显著差异;与对照组相比,干旱处理韧皮部3.5 m处可溶性糖及NSC含量均显著升高,10.5 m处可溶性糖含量显著降低,而其淀粉含量显著升高,其余高度的各指标均无显著变化。另外,对于细根而言,干旱处理组细根的可溶性糖、淀粉及NSC含量均与相应对照组无显著差异。
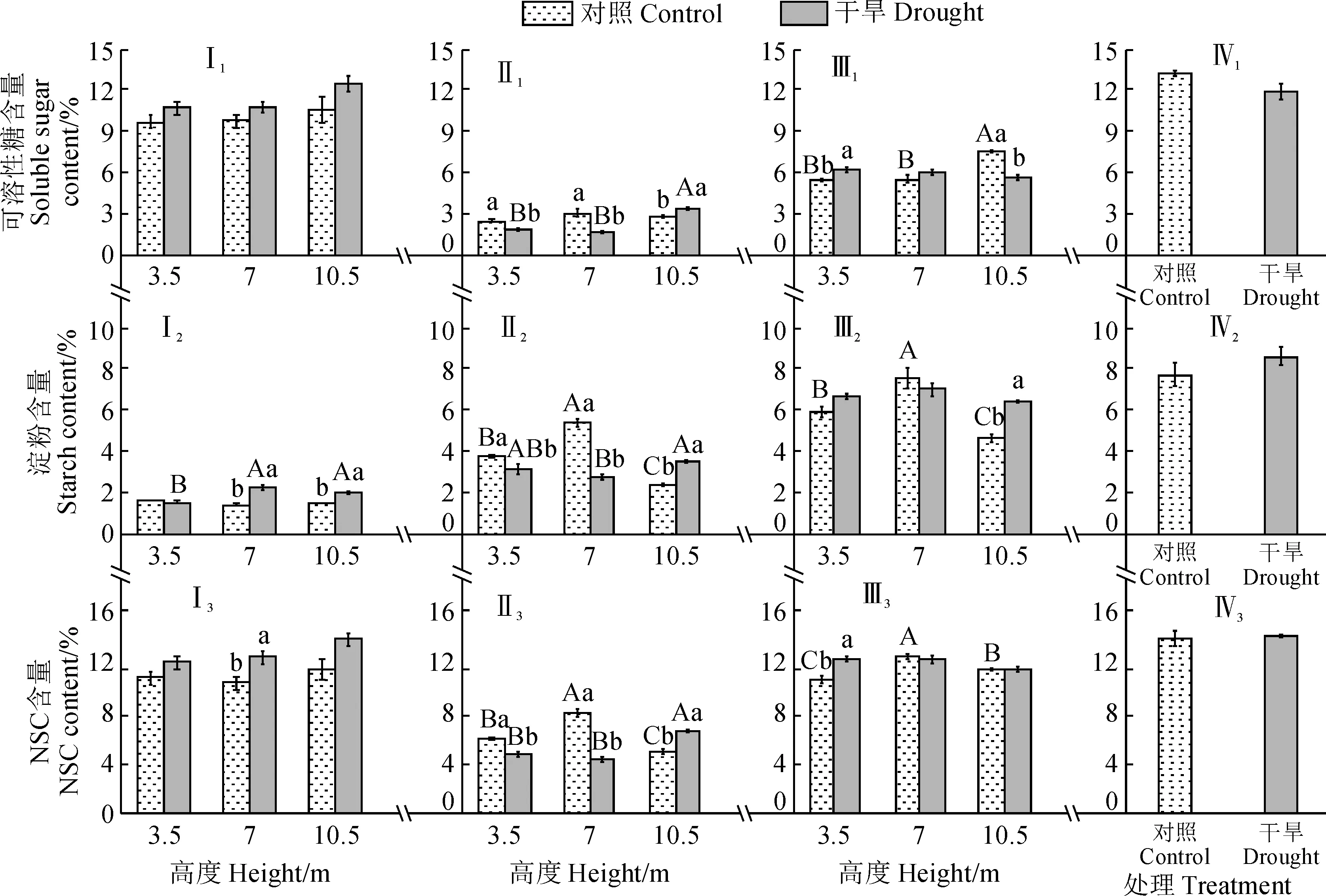
Ⅰ.叶;Ⅱ.枝条;Ⅲ.韧皮部;Ⅳ.细根;1.可溶性糖;2.淀粉;3.NSC
2.4 新疆杨不同树冠高度的比叶面积和胡伯尔值比较
新疆杨对照组和干旱处理组比叶面积在各树冠高度间均无显著差异,而干旱处理组比叶面积在各树冠高度下均显著低于相应对照组(图5,Ⅰ);对照组叶片胡伯尔值随树冠高度呈现先降后升的趋势,且3.5 m处显著高于7 m及10.5 m处,干旱处理组胡伯尔值在不同树冠高度间无显著差异,在各高度下与相应对照组之间也无显著差异(图5,Ⅱ)。
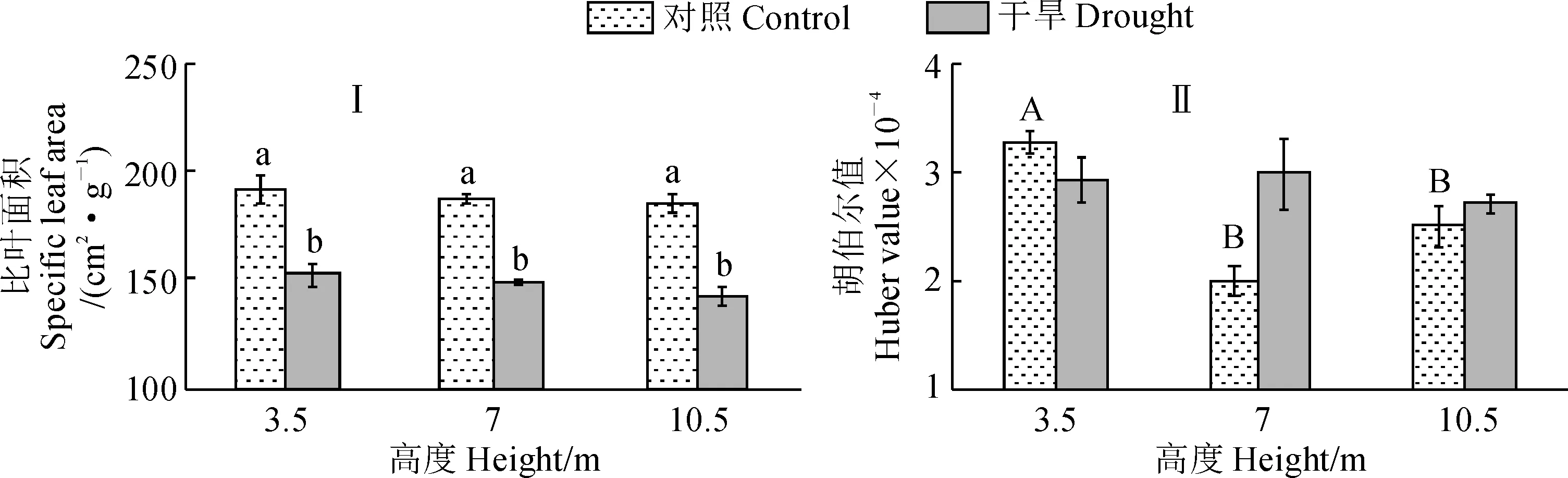
图5 两种水分处理下新疆杨不同树冠高度的比叶面积和胡伯尔值(n=3)
3 讨 论
3.1 新疆杨树冠不同高度叶片水力性状对干旱胁迫的响应
叶片水力性状取决于植物的生物学特性以及生长过程中对环境变化的适应[30]。树冠垂直层次的高度及土壤含水量的差异使新疆杨叶片水力性状发生不同程度的变化。本研究中,干旱处理使得新疆杨黎明叶水势、正午叶水势、叶含水量及枝条含水量总体随着树冠高度的增加而下降。这与Fang等[20]对小青杨(Populuspseudosimonii)的研究结果一致。树冠高度的增加使水的重力(水分的重力作用会导致木质部形成0.01 MPa·m-1的水势梯度[31])及其与导管内壁的摩檫力增加,阻碍水分的运输[32],从而减少了输送到树冠上部的水分。
等水植物对土壤含水量及饱和水汽压差的变化较为敏感,并能及时作出响应(如调节气孔导度),维持相对稳定且较高的正午叶水势[33-34]。根据杨柳等[33]对等水植物的研究,若正午叶水势变化小于1.0 MPa则可将其划分为等水植物。本研究中,除第216、234及243天3.5 m处,对照及干旱处理组新疆杨各树冠高度不同季节的正午叶水势变化普遍小于1.0 MPa,表明新疆杨属等水植物。这与Tardieu等[34]对欧美杨(Populuseuramericana)、Schultz等[35]对葡萄(VitisviniferaL.)的研究结果一致,表明等水植物的正午叶水势并不会随土壤水分含量发生明显变化。同时,本研究中2020年新疆杨除树冠高度10.5 m处的叶含水量外,干旱处理并没有使其余树冠高度叶含水量及枝条含水量与对照组之间产生显著差异。而Sade等[10]的研究指出,等水植物有维持稳定且较高组织相对含水量的能力,与本研究相似。此外,长期的干旱会促使新疆杨叶形态进行相应调整,如减小比叶面积、增加胡伯尔值。一方面,比叶面积反映出植物生长过程中的资源获取策略[36],减小了比叶面积有利于新疆杨减少水分散失[37]。另一方面,提高胡伯尔值可有效增加新疆杨枝条向叶片的供水能力[38],在一定程度上减小了干旱处理组与对照组间水分状况的差异。与Fang等[20]的研究不同的是,新疆杨叶片饱和膨压渗透势与膨压损失点水势在不同处理及树冠高度间的差异并不显著。这可能表明在长期干旱胁迫下,新疆杨并不是通过改变细胞的耐失水能力来适应干旱胁迫,而可能通过一些其他的补偿性调节来保证其生理活动的正常进行,如对叶片的形态或生理状况进行调整。
3.2 新疆杨树冠不同高度叶片光合生理参数对干旱胁迫的响应
叶片气孔导度的调节可防止叶和茎水势过度下降所引发的水分传导失效[9],但同时会阻碍水气交换降低光合及蒸腾速率。在本研究中,对照及干旱处理组新疆杨的净光合速率均随树冠高度的增加呈下降趋势,与Koch等[22]对红杉(Sequoiasempervirens)的研究结果一致。由于新疆杨属等水植物,叶片气孔对水分环境变化敏感[9],对照及干旱处理组树冠中上部的正午叶水势均有所下降,这可能促使叶片气孔关闭[10],进而导致净光合速率下降[22]。此外,干旱处理下新疆杨的水分利用效率随树冠高度的增加而提高,与何春霞等[23]对4种乔木的研究以及张永娥等[39]对侧柏(Platycladusorientalis)的研究结果一致,表明干旱胁迫下新疆杨在器官水平存在碳水收支权衡。叶绿素在叶片中起到光能吸收、传递和转换的作用, 在处于逆境时会发生不稳定降解[40]。本研究发现叶绿素SPAD值在新疆杨树冠的上部要低于下部,与姚春娟等[41]的研究结果一致,而水分胁迫是造成叶绿素相对含量下降的主要原因。
3.3 新疆杨不同器官非结构性碳水化合物含量对干旱胁迫的响应
NSC在植物体的含量反映了碳供需的平衡关系[22],其中淀粉和可溶性糖占NSC总量的90%以上[42]。可溶性糖在植物受到干旱胁迫时可作为渗透调节物质,而淀粉可作为储能物质[43]。本研究表明,干旱处理并没有使新疆杨树冠不同高度叶片的可溶性糖及NSC含量与对照组产生差异,但会使树冠中部及上部的淀粉发生累积。这与Yan等[44]对红松(Pinuskoraiensis)的研究结果一致。其可能的解释是:一是叶片净光合速率虽然随树高的增加有所下降,但水分利用效率有一定的提高,在一定程度上缓解了新疆杨上部由于水分不足致使气孔关闭带来碳同化量降低的风险[45];二是在同化物分配时,距离碳源近的器官将优先分配得到光合产物[46],且功能正常的叶所生产的光合产物通常大于自身的呼吸消耗[44],因此分配给叶片的光合产物会有一定量的累积,减小了叶片中可溶性糖及NSC含量在树冠不同高度的差异。干旱处理使得新疆杨树冠上部枝条的可溶性糖、淀粉和NSC含量高于下部,与Woodruff等[21]对花旗松的研究结果一致,该现象与新疆杨上部水分条件较差有关。在遭受干旱胁迫时,非等水植物能储存更多非结构性碳,具有更高的生产力[10]。而对于新疆杨这类等水植物而言,气孔对水分环境敏感,气孔导度下调将使得碳同化率下降,而此时NSC的储存量对植物抵御干旱胁迫尤为重要,因为 NSC是栓塞修复、渗透调节的重要物质[14]。
多数植物的光合产物以可溶性糖的形式由韧皮部从“源”运输到“汇”,以满足植物各器官对光合产物的需求[47]。有研究表明,干旱胁迫会导致韧皮部的含水量降低、汁液粘度增加,从而限制韧皮部对光合同化物的运输[16]。与干旱胁迫导致韧皮部运输受阻原理相似,王文杰等[48]对红松(Pinuskoraiensis)进行的环剥实验发现,环剥会阻碍光合产物的纵向运输,导致环剥上部韧皮部NSC积累。而在本研究中,干旱处理下新疆杨各树冠高度韧皮部的可溶性糖、淀粉、NSC含量没有显著差异,这表明干旱胁迫并没有影响韧皮部的装载功能,可保证光合产物正常运输,按各器官的需求进行碳分配。另外,细根是植物水分与养分吸收的主要器官。本研究中干旱处理并没有使新疆杨细根中的可溶性糖、淀粉及NSC含量与对照组产生显著差异。这与韧皮部运输功能的正常运作密不可分,保证了光合产物可从“源”(叶)向“汇”(根)进行运输。
4 结 论
干旱胁迫下,新疆杨植株受到的水分胁迫随树冠高度的增加而加剧,其中黎明叶水势、正午叶水势、叶片含水量、枝条含水量在树冠上部要普遍低于下部但饱和膨压渗透势及膨压损失点水势在不同树冠高度下并无显著差异,而随着干旱期的延长新疆杨树冠上部发生栓塞枯梢的风险要大于下部;净光合速率随树冠高度的增加有所下降,而水分利用效率有一定程度的提高,叶绿素SPAD值在树冠上部要低于下部;NSC在不同器官之间的分配有所差异,且其组分会随水分状况发生变化,叶片可溶性糖、NSC含量在不同高度间差异不显著,而淀粉含量在树冠的中部及上部要大于下部,枝条上部的可溶性糖、淀粉和NSC含量高于中部和下部,韧皮部的可溶性糖、淀粉和NSC在不同树冠高度间没有显著差异;细根的可溶性糖、淀粉及NSC含量在不同水分处理下差异不显著。
猜你喜欢
干旱地区农业研究(2021年6期)2021-12-08
环境卫生工程(2021年1期)2021-03-19
农村实用技术(2020年9期)2020-11-19
种子(2020年9期)2020-10-22
东方企业家(2020年5期)2020-05-29
山西文学(2019年8期)2019-11-01
湖北林业科技(2019年4期)2019-09-16
种子(2019年8期)2019-09-11
文学港(2019年5期)2019-05-24
青年文学家(2017年28期)2017-11-28
