
碳四植物光合生化机理模型的叶片含水量修正
2022-05-23 09:51冯晓钰周广胜
应用气象学报 2022年3期
冯晓钰 周广胜
1)(浙江省金华市气象局, 金华 321000)2)(中国气象科学研究院, 北京 100081)
引 言
光合作用是绿色植物利用太阳能同化大气中的二氧化碳和水汽,制造有机物并释放氧气的过程,是整个生态系统的能量来源和物质基础,也是地球碳氧循环的重要过程。植物光合作用模拟是陆地碳循环模拟的重要组成部分,在陆面过程和全球大气-土壤-植被相互作用模拟中起重要作用[1]。随着控制试验水平的提高和试验设备、观测技术的改善,植物叶片光合作用对各种环境因子(光照强度、二氧化碳浓度、温度、水分)的响应模拟得到较大发展,建立了多种光响应曲线模型、二氧化碳响应曲线模型和水分影响函数等[2]。有关光合作用的研究从现象描述到机理阐述[3],逐步发展到光合生化机理模型研究,其中Farquhar等[4]提出的碳三植物光合生化机理模型被广泛应用。该模型基于光合作用,涉及核酮糖-1,5-二磷酸羧化酶受限阶段、1,5-二磷核酮糖再生受限阶段、磷酸丙糖利用率受限阶段3个生物化学过程,考虑了与光合作用密切相关的叶绿素影响,建立光合作用和光合有效辐射、胞间二氧化碳浓度、叶片温度和叶片氮含量等因素的关系,为准确表述叶片光合作用奠定基础。
Farquhar模型最初研究无水分胁迫状态下探讨植物光合参数对温度和光强的响应,后经Harley等[5-6]修正,完善了二氧化碳浓度增加对植物光合作用的影响,但未考虑植物光合参数对水分变化的响应。Foley等[7]在建立整合生物圈模型时利用的叶片光合作用模型考虑了碳三、碳四植物对光照强度、二氧化碳浓度以及叶片温度的响应,但并未包含水分影响。水分是光合作用的重要原料之一,直接影响叶片气孔导度、蒸腾速率、光合速率等[8]。
近年关于水分变化影响光合作用的研究得到重视,逐渐从定性分析[9]转向定量模拟,经过大量试验,从经验模型[10-12]发展至生理生态模拟[13-15]。
尽管已有学者探讨水分变化对植物光合生物化学特性的影响[16],将最大羧化速率和最大电子传递速率引入土壤含水量影响函数[17-18],以表示水分变化对叶片光合作用的影响,但关于植物叶片光合作用与水分关系的定量研究较少,且多采用土壤含水量表示,而非起直接影响作用的叶片含水量,不利于光合作用的准确模拟。
叶片光合作用模型是冠层、生态系统、区域以及全球尺度下光合作用模拟的基础,为准确模拟叶片含水量对植物光合作用的影响,本研究基于von Caemmerer[19]修正后的碳四植物光合生化机理模型,结合玉米干旱影响模拟试验数据,以叶片含水量变化为切入点,研究碳四植物叶片光合生化机理模型的叶片含水量修正函数。
1 试验及模型简介
1.1 水分试验设计及影响变量控制
本研究通过控制变量试验,将影响叶片光合作用的光照、二氧化碳、温度等因素尽量控制一致,同时控制取样叶片的一致性,使其具有光合性能代表性,进行水分控制试验,以叶片净光合速率为指标,研究其与叶片含水量的关系,经归一化得到碳四植物光合作用生化模型的水分修正函数。
2014年在中国气象科学研究院固城生态与农业气象试验站开展玉米不同水分梯度持续干旱影响模拟试验,该区属于温带大陆性季风气候,多年平均气温为12.2℃,年平均降水量为528 mm,年平均日照时数为2264 h。试验区土壤为沙壤土,0~50 cm 深度平均土壤容重为1.2 g·cm-3,平均田间持水量为22.1%,全氮为0.98 g·kg-1,全磷为1.02 g·kg-1,全钾为1.02 g·kg-1,pH值为8.19。
试验共分6个处理,每个处理3个重复,共18个试验小区。播种前进行底墒控制,出苗后(三叶期)按照保定地区1981—2010年7月平均降水量的7%(W1)、20%(W2)、40%(W3)、60%(W4)、80%(W5)和100%(W6)6个水分梯度的灌溉量进行一次性灌水处理后,不再灌溉,分别于7月11日、7月18日、7月31日、8月7日和8月20日开展观测,试验设计、试验材料、测定内容与方法等详见文献[20]。叶片净光合速率利用美国LI-COR公司生产的LI-6400型便捷式光合系统分析仪测定。
6个水分处理的叶片含水量和土壤含水量观测值如表1和表2所示,玉米叶片含水量受土壤含水量影响显著,二者呈非直角双曲线关系[20]。经计算,当土壤相对湿度为100%时,玉米叶片饱和含水量为85.14%。

表1 2014年不同水分处理的叶片含水量(单位:%)Table 1 Leaf water content at different watering treatments in 2014(unit:%)

表2 2014年不同水分处理的土壤含水量(单位:%)Table 2 Relative soil water content at different watering treatments in 2014(unit:%)
研究表明,碳四植物为低二氧化碳补偿点植物,大气二氧化碳浓度不变条件下不同处理的叶片净光合速率差异不大;夏玉米是喜温的碳四植物,光合有效辐射和土壤水分是影响其生理指标最主要的生态因子[21]。为控制光合有效辐射的影响,7月10日(首次观测7月11日前1 d)对处于三叶期-七叶期的W1~W6处理进行光响应曲线测定,发现叶片净光合速率在光合有效辐射为1500 μmol·m-2·s-1时达到饱和(图1)。因此,选择气象条件稳定的晴天,在气温为31℃~33℃、二氧化碳浓度约为4×10-4、大气相对湿度约为50%条件下,采用荧光叶室,设定光合有效辐射为1500 μmol·m-2·s-1,流速为300 μmol·s-1,在09:00—12:00(北京时,下同)进行观测。每个小区选取1株玉米标准植株测定其顶部第1片完全展开叶中上部位置的净光合速率An(单位:μmol·m-2·s-1),以保证取样叶片对植株光合性能的代表性。

图1 不同水分梯度持续干旱处理下的光响应Fig.1 Light response under consecutive drought treatments with different water gradients
1.2 碳四植物光合生化机理模型
von Caemmerer[19]将Farquhar等[4]提出的碳三植物光合生化机理模型应用于碳四植物,其中叶片光合作用速率An受核酮糖-1,5-二磷酸羧化酶活性限制:
An=Vc·A-Rd,
(1)
(2)
式(1)和式(2)中,An是叶片净光合速率,为观测值;Vc为最大羧化速率(单位:μmol·m-2·s-1);A为参数;Rd为暗呼吸速率(单位:μmol·m-2·s-1),为0.01Vc;C是叶绿体碳分压(单位:μbar),为观测值;Kc为核酮糖-1,5-二磷酸羧化酶的二氧化碳米氏常数(单位:μbar),取650;O为叶绿体氧分压(单位:mbar),取210;Ko为核酮糖-1,5-二磷酸羧化酶的氧气米氏常数(单位:mbar),取450;γ*为核酮糖-1,5-二磷酸羧化酶特异性倒数的一半(单位:bar·bar-1),取0.000193。以上参数见文献[4,22]。
Massad等[22]将该碳四植物光合生化机理模型应用于玉米,指出在饱和光强或接近饱和光强时,在核酮糖-1,5-二磷酸羧化酶限制的光合作用对温度响应上,碳四植物比碳三植物更敏感;碳四植物的温度对Vc敏感,但对Kc和Ko敏感性较弱。因此,在Arrhenius方程的基础上考虑高温降低酶活性,得到最大羧化速率Vc的温度修正函数:
(3)
(4)
式(3)和式(4)中,f(Tk)为温度修正函数,Tk为绝对叶温(单位:K),为观测值;k为25℃时的参数,Ea为活化能(单位:KJ·mol-1),取58.52;R为摩尔气体常数(单位:J·K-1·mol-1),取8.31;B为方程中的参数,ΔS为熵(单位:J·K-1·mol-1),取710;Hd为失活能(单位:KJ·mol-1),取220。
由式(1)~(4),得
(5)
式(5)中,V*为25℃时最大羧化速率,单位:μmol·m-2·s-1。
2 结果与分析
2.1 试验变量控制
本试验较好地实现了环境变量和试验变量的控制,图2和图3分别为2014年7月11日首次观测不同水分梯度持续干旱处理下的环境变量和土壤相对湿度变化。图中主要包含平均值(柱状)、误差值(误差棒)和显著性差异结果(字母),其中显著性差异结果以显著性差异字母标记法标注。经平均值排序后,最大项标记a,并与以下各项比较,差异不显著各项均标记a,直至某一项与之差异显著,标记b;再以标记b为准,与以下相比,差异不显著各项均标记b,直至某一项与之差异显著,标记c,以此类推。标记为同一字母表示差异不显著,标记为不同字母表示差异显著。

图2 不同水分梯度持续干旱处理下的环境变量Fig.2 Environmental variables under consecutive drought treatments with different water gradients
由图2可知,6个不同水分梯度处理间的环境条件如温度平均值约为31.4℃,二氧化碳浓度为4.00×10-4~4.05×10-4,空气相对湿度约为50%,光合有效辐射约为1500 μmol·m-2·s-1,均未达到0.05显著性水平。由图3可知,W6处理下土壤相对湿度最大,平均为92.4%,标记a,与W5处理下土壤相对湿度(88.5%)差异未达到0.05显著性水平,而与W4处理下土壤相对湿度(81.8%)差异达到0.05显著性水平,同时W5与W4处理下土壤相对湿度差异也未达到0.05显著性水平,则W5处理下土壤相对湿度标记ab,W4处理下土壤相对湿度标记b,以此类推,分别将W3,W2,W1处理下土壤相对湿度标记为c,d,e,即除因灌溉设置(三叶期一次性灌溉),高水分处理下均达到饱和,使得7月11日观测时W5与W4,W5与W6处理下土壤相对湿度差异未达到0.05显著性水平外,其余处理两两间的土壤相对湿度差异均达到0.05显著性水平。

图3 不同水分梯度持续干旱处理下的土壤相对湿度Fig.3 Relative soil moisture under consecutive drought treatments with different water gradients
2.2 不同水分梯度持续干旱处理下的温度修正
6—10月开展的玉米干旱模拟试验中,叶片光合生理观测时采用荧光叶室,光合有效辐射均为1500 μmol·m-2·s-1,温度、湿度和二氧化碳浓度均为环境条件,LI-6400光合仪测定相应的光合生理参数。玉米为碳四植物,在光饱和条件下,温度是影响叶片生理参数的重要因子[23]。试验期间共开展5次观测,为排除不同时次观测下的叶片温度影响[24],将叶片温度和叶片净光合速率的观测值带入,并根据式(5)进行温度修正,得到25℃时最大羧化速率V*,该V*仅受叶片含水量的影响。
2.3 最大羧化速率与叶片含水量的关系
经环境变量控制和温度修正后的观测结果显示,25℃时最大羧化速率V*与叶片含水量L呈显著的二次曲线关系(图4)(达到0.01显著性水平)。本试验的温度稳定在31℃~33℃,为玉米的最适温度范围[25]。因此,经温度修正,拟合方程的决定系数有所提高,但仍存在因不同程度水分胁迫引起的发育期差异(图5),导致个别值偏小。
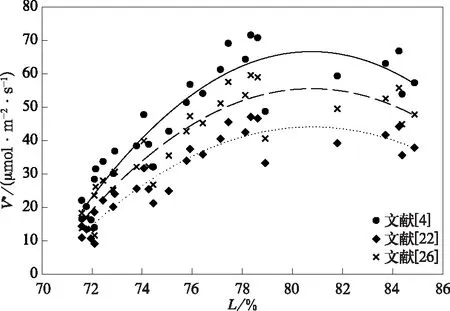
图4 V*与L的关系Fig.4 Relationship between V* and L

本研究以同样应用于玉米的参数(文献[22])为准,发现V*随着L的增加先逐渐增加,当L=80.79%时,V*达到最大(44.22 μmol·m-2·s-1),其后随L增加逐渐减小。当L<70.03%时,V*减小为零,即光合作用为零。值得注意的是,方程还有一个解为93.55%,但此时已经大于饱和叶片含水量,因涝渍影响光合特性[27]。

f(L)=-0.0086×L2+1.3966×L-55.4170。
(6)

图5 不同水分梯度持续干旱处理下的发育期Fig.5 Growth stages under consecutive drought treatments with different water gradients

表3 不同Kc和Ko取值下的拟合方程Table 3 Fitting equations under different values of Kc and Ko

图6 归一化后的叶片含水量影响曲线Fig.6 Normalized leaf water content influence curve
3 结论与讨论
以碳四植物玉米为研究对象,结合不同水分梯度下玉米光合作用数据和光合生化机理模型,控制光照、温度、二氧化碳浓度、相对湿度等环境影响因素,定量模拟叶片含水量变化对叶片光合作用的影响,主要结论如下:
1) 建立基于直接影响叶片光合作用的叶片含水量响应函数,完善包含叶片含水量影响的碳四植物核酮糖-1,5-磷酸羧化酶活性限制光合作用生化机理模型,为进一步提高光合作用模拟的准确性和玉米干旱监测预警提供参考。
2) 玉米的最大羧化速率与叶片含水量呈显著的二次曲线关系,其拟合方程的决定系数达0.88,参数不同时,最大羧化速率的绝对值不同,但归一化后的叶片含水量修正函数与参数无关,当叶片含水量为80%左右时,f(L)达到1.0,当叶片含水量降至70%左右时,f(L)为0。
3) 环境条件是影响玉米光合速率的因素之一,经环境变量控制后的叶片含水量修正函数决定系数达到0.88,但因水分条件差异引起的玉米发育期不同等生物因素,也在一定程度上影响光合作用模拟的准确性。
植物生理生态模型是研究全球变化对植被影响的重要手段之一,也是陆地生态系统模型研究的重要方向。叶片尺度的光合作用是陆地生态系统模型的基础,受到生态学家的广泛关注。20世纪80年代以来,Farquhar模型是叶片尺度光合作用研究的最重要进展,大量研究揭示叶片净二氧化碳同化速率随光照强度、二氧化碳浓度以及叶片温度的变化规律,并在大尺度模型中有所体现,但对于叶片含水量变化的影响研究较为欠缺。目前主要基于土壤水分变化对叶片光合生理参数的影响研究,或从叶片含水量变化影响气孔导度间接研究对光合作用的影响。叶片水分是进行光合作用的重要原料,叶片含水量状况直接影响其光合速率;同时,叶片也是作物生长对水分亏缺十分敏感的形态指标,一旦水分亏缺迅速通过叶片的变化反映在作物的长势和长相上。因此,叶片含水量是大气干燥度、土壤供水能力和作物耐旱能力综合作用的体现。特别重要的是,叶片长势还是重要的遥感探测指标[30-31]。近代科学技术的快速发展已经为研究作物叶片含水量的变化提供了强有力的支撑[32],遥感探测技术的发展也为连续动态大范围监测作物长势提供技术保障。因此,建立基于叶片含水量变化的叶片光合作用水分响应函数不仅可以提高光合作用模拟的准确性,也可以通过光合速率获取作物干旱致灾临界叶片含水量和作物干旱临界气象条件,进而结合遥感长时间、大范围干旱发生发展动态监测[33],为科学制定农业抗旱对策措施,确保农业生产安全和粮食安全提供重要的理论依据。
本研究为保证取样叶片对植株光合性能的代表性,仅观测了玉米植株顶部第1片完全展开叶(简称第1叶)中上部位置的净光合速率。研究发现不同叶位的最大叶片含水量不同,随干旱进程的推进,叶片含水量的变化速率也不同[34]。尽管第1叶的最大含水量高于其他叶位,其叶片含水量的下降速率高于其他叶位,且可进行光合产物累积的叶片水分下限(净光合速率为零时的叶片含水量)最小,其耐旱性最强,在干旱监测中最具指标意义,但是通过第1叶的叶片含水量换算得到的冠层含水量存在偏大情况。因此,在应用该模型到冠层尺度时,模拟的冠层光合作用偏大,需要考虑对整个冠层不同叶位叶片光合作用能力的差异。
本研究利用玉米不同水分梯度持续干旱模拟试验数据,在已有碳四植物生化机理模型的基础上,结合试验条件(饱和光强、当前大气分压),确定玉米主要受核酮糖-1,5-二磷酸羧化酶活性限制的光合作用,并进行温度修正以排除温度变化对叶片光合作用的影响,从而定量模拟叶片含水量变化对叶片光合作用的影响。根据拟合发现, 最大羧化速率随叶片含水量的变化呈显著的二次曲线关系,作物生长过程中叶片含水量过少或过多,均可降低叶片净光合速率,甚至减小为零。
本研究中,经温度修正后拟合方程的决定系数仅针对玉米,较多种植物类型对叶片最大羧化速率影响[15]的结果提高明显。环境因子是影响植物光合作用的因素之一,不同发育期也存在差异。随着干旱发生发展,玉米的生长发育,不同发育期的叶片净光合速率[35]不同。因此,未来需进一步区分作物不同发育期,完善叶片光合作用生化机理模型。
猜你喜欢
中原商报·科教研究(2022年1期)2022-05-13
种子(2019年8期)2019-09-11
特别健康(2018年9期)2018-09-26
中国饲料(2016年12期)2016-11-07
中国眼镜科技杂志(2016年18期)2016-11-03
Coco薇(2016年5期)2016-06-03
考试周刊(2016年6期)2016-03-11
少儿科学周刊·儿童版(2015年10期)2015-11-07
高中生学习·高三版(2014年3期)2014-04-29
中国氯碱(2014年10期)2014-02-28
