
水稻锌吸收、转运、累积机理研究进展
2022-06-09 08:47薛欣月于雪然刘晓刚马嘉欣田蕾李培富
生物技术通报 2022年4期
薛欣月 于雪然 刘晓刚 马嘉欣 田蕾 李培富
(宁夏优势特色作物现代分子育种重点实验室 宁夏大学农学院,银川 750021)
锌是人体必需的微量元素之一,主要分布于人体的肌肉、血液和骨骼中[1],参与人体多种新陈代谢过程、包括DNA聚合酶等多种酶的合成、细胞分解、能量和其他物质的代谢,其不足阻碍人体的生长发育[2],锌对人体健康具有十分重要的调控作用。目前,我国部分地区的儿童及青少年存在不同程度的锌营养缺乏症,但直接服用高锌含量药物不利于人体健康[2]。研究表明植物体中的锌可以更安全、高效地被人体吸收,因此锌在植物体中的转运机理受到研究者的高度重视[3]。水稻作为主要粮食作物,提高水稻籽粒锌含量,将会有效缓解人类锌缺乏的困境。研究水稻体内锌元素吸收、转运和累积机理,可以为富锌水稻的创制奠定理论基础。
1 锌在水稻体内的生理功能
1.1 生物体内多种酶的组分和活化剂
锌在生物体内主要以二价阳离子形态(Zn2+)存在,且是多种酶的组成成分。研究表明,动物、植物、微生物中至少已鉴定出300多种酶的活性与功能受到锌元素的调节[4]。其中,参与水稻生理功能的锌酶(zinc enzymes)主要分为含锌金属酶和需锌酶两大类(表1)。

表1 锌酶及其可能功能Table 1 Zinc enzymes and their possible functions
锌是水稻体内多种含锌酶的组成成分,在含锌酶中主要发挥的作用包括催化作用、结构功能、辅酶、共催化功能等。碳酸酐酶存在于水稻叶绿体外膜,参与光合作用[7],当水稻植株内锌元素缺乏程度加剧时,叶片中碳酸酐酶的活性降低,阻碍了水稻植株的光合作用,水稻产量将会大幅降低[5]。锌是合成谷氨酸过程中必不可少的元素,是谷氨酸脱氢酶(GDH)的主要成分,谷氨酸脱氨酶普遍存在于水稻线粒体内,在水稻碳氮代谢中具有重要作用[8]。锌是醛缩酶的激活剂,醛缩酶主要存在于细胞质内,参与水稻光合作用碳代谢过程。铜锌超氧化物歧化酶(Cu/Zn-SOD)是1种重要的含锌金属抗氧化酶,主要存在于水稻细胞质和叶绿体中[9],其中含有Cu2+和Zn2+两个金属辅基,铜锌超氧化物歧化酶对水稻抗氧化、抗逆、抗衰老具有重要意义,Sen等[13]研究表明在水稻中具有充足的Cu/Zn-SOD时,能显著提高水稻的抗盐、抗旱及抗寒性等。
锌是水稻体内过氧化氢酶(catalase)、核酮糖-1,5-二磷酸羧化酶(rubisco)、过氧化物酶(peroxidase)等多种需锌酶的活化因子。过氧化氢酶分布于水稻叶绿体中,参与光呼吸、共生性氮固定,当锌营养缺乏时,CAT活性降低,容易导致生物膜的过氧化损伤,引起有机物和无机物从根细胞渗漏[7]。核酮糖-1,5-二磷酸羧化酶是光合作用碳反应中重要的羧化酶,当水稻中锌营养适当时,有利于光合色素的形成,Rubisco活性增强,保证光合作用正常进行,促进水稻种子萌发、根系发育[10]。过氧化物酶可促进水稻光合作用、呼吸作用,研究表明,不同浓度Zn2+对水稻体内的POD活性影响不同[12]。
1.2 参与生长素的合成
锌元素能促进水稻体内吲哚和丝氨酸合成色氨酸,而色氨酸是生长素合成的必要条件,因此锌间接影响生长素的形成。锌缺乏会导致水稻中生长素含量降低,从而使水稻植株矮小[14]。水稻生长旺盛的部位都含大量的锌和生长素[15-17],缺锌会使植株中生长素含量降低,影响植株生长发育速度,蛋白质合成受阻,生物量明显下降,结实率下降、空秕率上升,从而影响产量和品质[18]。
1.3 参与光合作用
Rubisco是水稻体内重要的需锌酶,存在于水稻叶绿体中,是水稻光呼吸以及光合作用中决定碳同化速率的关键酶[21]。植株体内锌含量过低时Rubisco活性降低,导致ATP中的化学能转化为葡萄糖的过程受到抑制[10]。1,6-二磷酸果糖酶主要存在于叶绿体中,参与光合产物的转运,1,6-二磷酸果糖酶是叶绿体中裂解六碳糖的关键酶,在植物缺锌时该酶活性显著降低,导致光合产物向细胞质中的转运受到抑制[7]。
1.4 锌与水稻病害
锌的抗病性可能与锌在维持膜的结构和功能方面的重要作用有关,对根系生物膜的保护作用尤为突出[22-24]。当水稻缺锌时,植株对病原体的抗性减弱,体内的天门冬酰胺和谷氨酰胺含量增加,加速了病原体的侵染和生长,导致水稻病害加重。此外,缺锌水稻根际碳水化合物和游离氨基酸渗出量增多,致使病原菌快速繁殖[25]。
水稻缺锌引发水稻缩苗病,往往在土壤有效锌含量低或氨锌比例失调时发病。轻度缺锌时,叶片出现褐色斑点,叶片失绿变白;缺锌严重时,植株生长受到抑制,新叶变小、植株矮小,致使水稻不能抽穗,明显矮缩[26-27]。
2 水稻锌的生物强化
水稻是世界上最重要的农作物之一,在全球大部分地区都有种植,是人类重要的主食之一。据估计,人类消耗的热量中有19%来自大米[28]。然而,稻米中的锌等矿物质含量低,因此锌是人类饮食中最常缺乏的营养元素[29]。在谷物中,大米的矿物质含量和遗传变异性都是最低的[30],考虑到大米的广泛消费和低营养品质,水稻的生物强化被认为是向人类提供锌含量增加谷物的最佳解决方案之一。生物强化是指使植物在其可食部分(如籽粒、叶子和根)积累更高浓度的可利用营养元素[30-31]。人们可通过增加稻米中锌的绝对含量来改善人体锌缺乏[32]。但稻米中存在植酸[33]、纤维素等因素,导致膳食中锌的生物有效性低[34]。研究表明在水稻中,植酸与二价离子(如 Zn2+、Ca2+、Mg2+、Fe2+)形成络合物,矿物的生物有效性显著降低[35-36],所以要通过提高水稻籽粒有效锌以及稻米中锌的绝对含量来进行生物强化。
3 锌与水稻产量及品质的关系
锌在水稻的生长发育中具有不可替代性。锌是水稻多种酶的构成元素,也参与光合作用、激素合成等多种重要的生理生化反应,在水稻的不同生育时期发挥着重要作用,从而影响水稻的产量和品质。
根据现有工艺流程,将硝化工艺的循环泵出口管线分成两路,第一路继续将好氧池底部的活性污泥循环至好氧池顶部,不影响硝化工艺的正常运行;第二路将好氧池底部的活性污泥循环至厌氧池,使其进行反硝化反应。并可根据需要调节第一、二两路的阀门开度,控制进入好氧池、厌氧池的循环污泥比例,使中水系统处于最佳的工作状态。
3.1 锌与水稻产量
祁明等[37]研究发现,在石灰性土壤中水稻易发生缺锌现象,水稻分蘖期缺锌非蛋白氮含量显著增加,蛋白氮总量减少。水稻内双氨基和多氨基的氨基酸含量明显降低,导致蛋白质的合成受阻。王孟兰[38]研究表明,在缺锌的土壤上施用锌肥,可有效促进氮素吸收,通过增加单位面积穗数和穗粒数增加水稻产量,但施锌量过高会抑制水稻对锌的吸收,并导致减产,水稻产量随施锌量的增加而先递增后减少。麦日桂[39]研究表明,在水稻生长期间,增施锌肥能有效改善水稻的农艺性状,以达到增产提质目标。
3.2 锌与水稻品质
施用锌肥可改善稻米品质,如提升稻米的食味值和香味,锌可增强水稻植株的光合作用,促进糖类代谢,改善稻米口感[40]。张丽等[41]研究发现,基施锌肥或基施+喷施锌肥可有效增加稻米的蛋白质含量,合理喷施锌肥可有效增加水稻籽粒中锌含量的积累,改善水稻籽粒营养品质,如降低稻米碳水化合物含量等。袁少文等[42]研究表明,稻米中锌含量与喷施锌肥浓度呈正相关,喷施浓度为2.5%时,稻米中锌含量最高。
4 水稻不同营养器官锌吸收途径
水稻植株对锌元素的吸收主要通过两种方式进行。一方面通过水稻根系通过土壤中有效锌的吸收及锌肥施用进行吸收,另一方面通过锌的叶面喷施锌肥的方式进行锌吸收。
4.1 水稻根系锌吸收途径
水稻根系锌吸收途径主要有两种,分别是质外体和共质体途径,共质体途径为主要方式[43]。质外体途径为自由扩散,无能量消耗,共质体途径为主动运输,消耗能量。Zn2+通过被动和主动运输由细胞膜进入根系细胞,在根中Zn2+从木质部薄壁细胞装载到木质部[44]。
4.2 水稻叶片锌吸收途径
水稻叶片锌吸收途径主要是叶面喷施锌肥。叶片对锌的吸收主要通过气孔或表皮细胞进入叶片表皮。但是由于叶片表面张力作用,纯水很难自由进入气孔,所以要借助外部力量,在叶面锌肥中加入表面活性剂可以降低叶片表面的张力,增加水稻叶片对锌肥的吸收量[45-46]。但有些表面活性剂会对水稻植株产生毒害作用,影响植株光合作用,所以要注意表面活性剂的用量。Zn2+最后大部分积累在叶、茎和籽粒中,而不是根部[47]。
5 水稻植株锌吸收、转运的影响因素
水稻植株在进行锌的吸收转运过程中会受到内在因素和外在因素的影响,制约水稻植株对锌的吸收转运。内在因素包括水稻品种多样性、水稻生育时期的影响和水稻籽粒锌含量相关QTLs的影响。外在因素包括土壤理化性质和农艺措施的影响[32]。
5.1 内在因素
5.1.1 水稻品种多样性 不同水稻品种对锌的吸收积累量存在显著差异。Yang等[48]对285份水稻品种的籽粒矿质元素含量进行测定,发现籽粒中锌含量籼稻>粳稻,红米>白米。研究表明云南稻核心种质的锌含量,地方品种明显高于改良品种,地方稻核心种质的锌含量在籼粳、光壳与否及有无芒之间差异不显著,但水陆、软米与普通米、紫米与红米间的锌含量差异极显著,以陆稻锌含量最高,软米则最低[49]。Gregorio等[50]对 1 138 份不同来源的水稻品种籽粒锌含量进行测定发现,锌元素在不同种质间的变异范围为15.3-58.4 mg/kg。
5.1.2 水稻生育时期 张标金[51]研究表明,不同水稻品种在分蘖盛期对锌的吸收能力均为最强,因为营养器官发育最关键的时期为水稻分蘖期,植株需大量的营养供生殖生长所需,此时根系活力增强,大量的矿质元素储存在根部。张济龙等[52]发现植株灌浆期锌含量低于成熟期,尤其是根和茎中的锌含量,这可能是由于锌在水稻根系中的积累浓度已达到峰值,而锌在水稻根中的积累主要在水稻灌浆期进行。但当锌浓度低于0.1 mg/L时,植株在灌浆期对锌的吸收量与成熟期相近。
5.1.3 水稻籽粒锌含量相关QTLs 锌在水稻籽粒中的积累受多基因控制,不同水稻品种籽粒锌含量的差异也受不同基因控制,国内外学者在水稻每条染色体上都发现了控制水稻籽粒锌含量的QTL(表2)。

表2 水稻籽粒锌含量相关QTL简表Table 2 A brief list of QTLs related to zinc content in rice grains
虽然近年在有关水稻籽粒锌含量的QTL定位的研究取得了一定进展,但由于水稻控制锌含量的遗传机制较为复杂,所以对相关基因的精细定位和克隆研究一直进展较慢。这些研究不仅为众多学者在水稻籽粒锌含量相关基因研究方面提供了经验,也为进一步阐明控制水稻籽粒锌含量基因的功能分析和相关目标基因的克隆奠定了基础,使通过分子标记辅助选择指导富锌水稻品种成为了可能。
5.2 外在因素
5.2.1 土壤理化性质 土壤中的有效锌可以反映土壤的供锌能力[68],有效锌受成土母质、土壤pH值等众多因素的影响。水稻植株对锌的吸收量因土壤质地的不同而变化,水稻土和红壤等类型的土壤有效锌含量均较高,草甸风沙土、石灰性紫色土和灌耕土等的有效锌含量较低[69]。植株根系对锌的吸收速率在砂土中明显高于壤土及黏土。除此以外,水稻中的锌元素来源于土壤有效锌和土壤速效锌,水稻植株可以直接吸收和利用土壤中的锌元素。速效锌可以衡量土壤的供锌能力,当其含量偏低时,水稻缺锌症状加重[70]。
5.2.2 农艺措施 施用锌肥被认为是一个水稻增产优质的有效措施。增施锌肥可以促进水稻生殖生长,增加粒重及产量。研究表明叶面喷洒锌肥能提高果实及种子中锌的积累量[17]。王晨等[71]研究表明施用硫酸锌可有效提高水稻籽粒锌含量,施锌能提高水稻不同营养器官的锌浓度,也可以通过施用氨基酸锌复合物来增加锌在籽粒中积累。
吴春勇等[23]认为水稻盛花期喷洒CTK可促进锌在剑叶中的积累。研究表明增加施氮量可促进水稻对锌的吸收与累积,氮肥与锌肥同时施用对水稻生长具有协助作用[72],增施锌肥能有效增加水稻籽粒的氮浓度和粗蛋白含量,其含量增加与施锌量呈正相关[73]。
6 锌转运蛋白
锌是植物中重要蛋白结构的辅助因子和活化剂。水稻中有大量的转运蛋白参与锌的吸收、转运和再转运,对水稻产量和营养品质都有重要影响[74]。水稻体内Zn2+的转运与锌转运蛋白家族密不可分。研究学者在多种高等植物中发现了锌转运蛋白家族,如YSL蛋白家族、ZIP蛋白家族、NRAMP蛋白家族(自然抗性巨噬细胞蛋白家族)、CDF蛋白家族、P1b-ATP酶、Mg2+/H+的反向交换转运蛋白家族、三磷酸结合盒转运蛋白等[74-76]。锌转运蛋白包括锌吸收蛋白以及锌排出蛋白两大类[77]。
6.1 锌吸收蛋白种类
锌吸收蛋白包括YSL蛋白家族、ZIP蛋白家族、NRAMP蛋白家族。
6.1.1 YSL蛋白家族 目前在拟南芥、玉米、小麦、水稻、大麦中都有发现YSL蛋白家族(yellow stripelike protein,YSL)的存在。YSL蛋白家族主要参与Fe2+、Mn2+、Cu2+等重金属离子的转运。但还未在水稻基因组中发现有关Zn2+吸收转运的的蛋白基因[78]。
6.1.2 ZIP蛋白家族 金属稳态对植物的生长、发育和对环境胁迫的适应至关重要,主要由各种金属转运体控制。植物ZIP(Zn-regulated transporter,Iron-regulated transporter-like Protein)家族蛋白属于负责吸收和分配必需和非必需金属的完整膜转运蛋白。参与锌吸收的蛋白为锌调控转运体(Zinc regulated transporter,ZRT),研究学者在拟南芥、水稻、苜蓿等植物中鉴定出多个ZIP基因[79-80]。已知水稻ZIP蛋白基因家族主要有9个锌调控转运蛋白基因,分别为OsZIP1、OsZIP2、OsZIP3、OsZIP4、OsZIP5、OsZIP6、OsZIP7、OsZIP8、OsIRT1,其表达部位、表达诱导条件及功能如表3所示。在水稻中,OsZIP4、OsZIP5、OsZIP7和OsZIP8在地上部和根中对锌的分配中起着重要作用,这些转运蛋白可用于提高锌的吸收或分配,改善水稻生长。
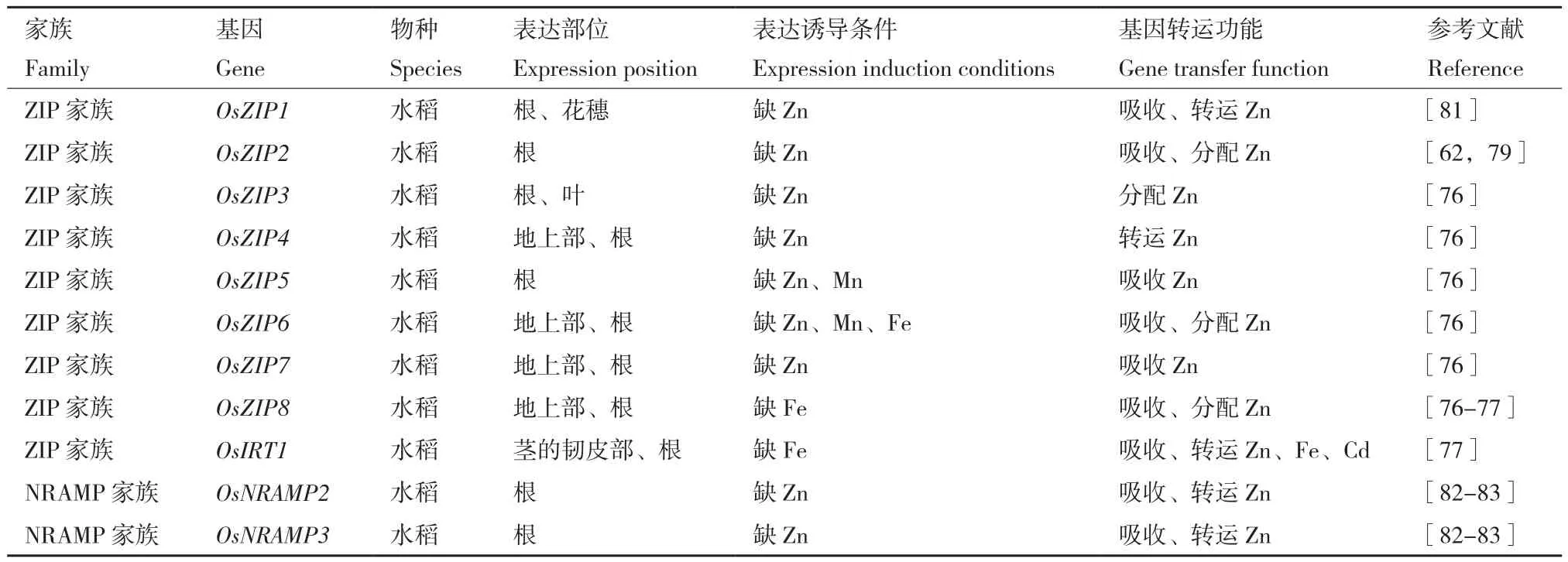
表3 锌吸收蛋白种类、功能及表达特征Table 3 Types, functions and expression characteristics of zinc absorption proteins
6.1.3 NRAMP蛋白家族 NRAMP蛋白家族为膜蛋白家族,广泛存在于各种生物体内,主要负责部分二价金属阳离子的吸收。孙丽娟等[82]研究表明,水稻中OsNRAMP2和OsNRAMP3参与锌的转运,其表达部位均位于水稻根部。图1为现阶段已发现水稻中11个锌转运蛋白成员的氨基酸序列比对,其亲缘关系如图2所示。
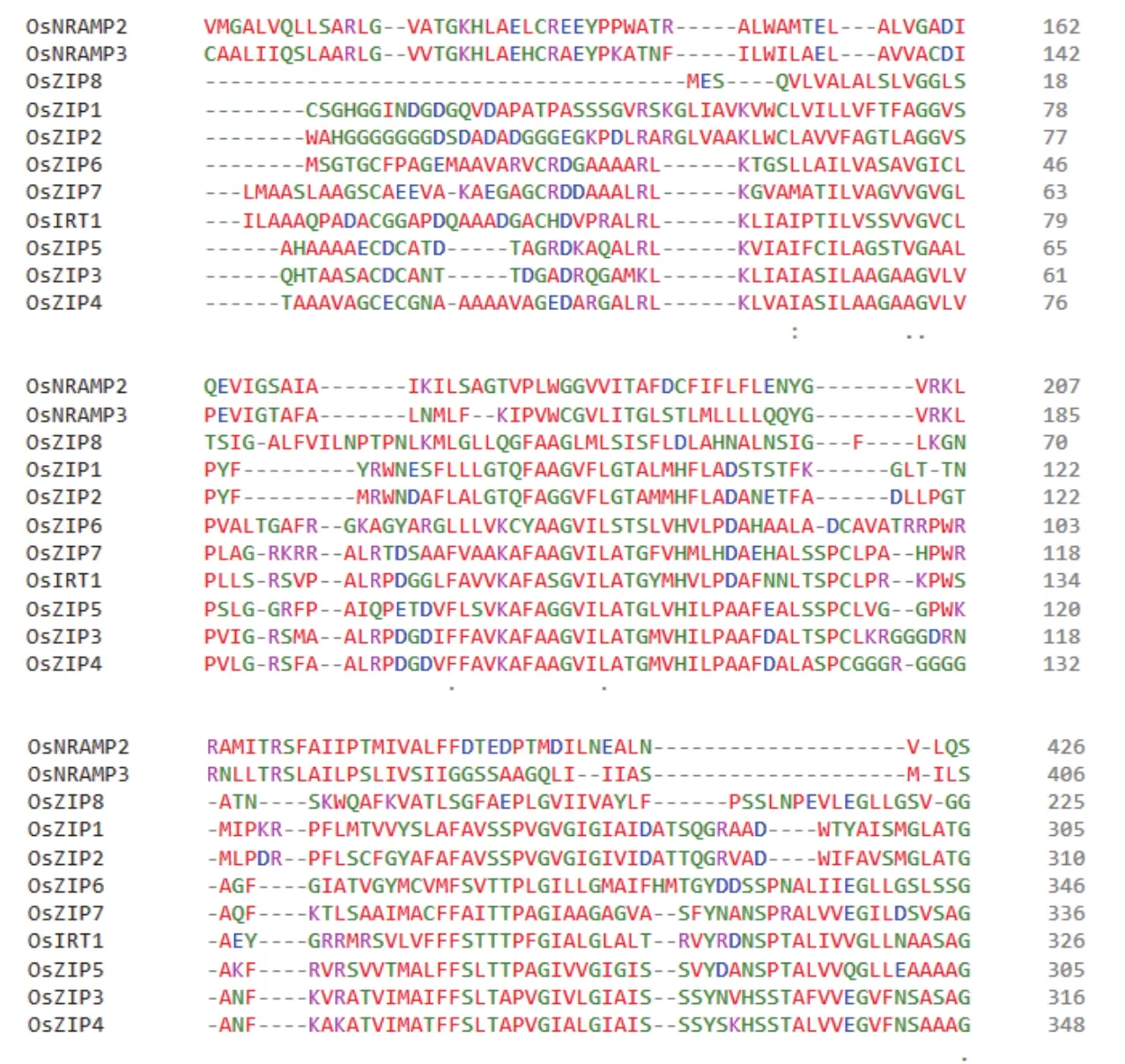
图1 锌吸收蛋白氨基酸序列比对Fig.1 Amino acid sequence alignment of zinc uptake proteins
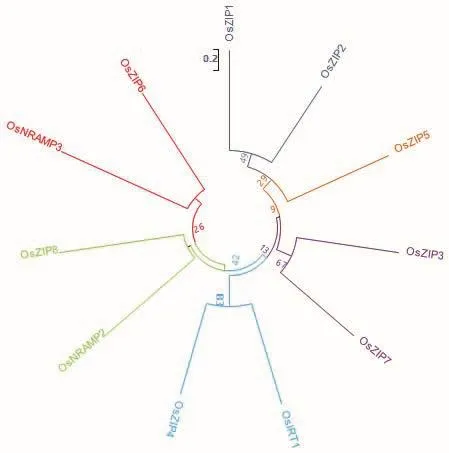
图2 锌吸收蛋白进化树Fig.2 Zinc-absorbing protein evolutionary tree
6.2 锌排出蛋白种类
当前发现的锌排出蛋白包括CDF蛋白家族、P1B型ATPases、Mg2+/H+的反向交换转运蛋白家族、三磷酸结合盒转运蛋白[76]。但目前还未在Mg2+/H+的反向交换转运蛋白家族中发现有关水稻Zn2+吸收转运的的蛋白基因。
6.2.1 CDF蛋白家族 CDF家族蛋白主要参与锌、镉和镍离子的转运,其作用是将过量的锌离子排出细胞,避免过量的锌对细胞产生毒害[83]。目前已发现有关水稻Zn2+吸收转运的的蛋白基因有OsMTP1和OZT1。OZT1在Zn、Cd胁迫下于水稻各部位处表达,参与锌和镉的转运[78]。
6.2.2 P1B型ATPases 研究学者在水稻基因组中发现9个基因(OsHMA1-9)[84],其中有3个基因为锌调控转运蛋白基因,为OsHMA1、OsHMA2、OsHMA3,分别作用于水稻地上部、根、叶,基因转运功能如表4所示。OsHMA2在营养生长期主要在根系成熟区表达,在生殖期主要在节间达,它的表达不因锌的有效性而改变。OsHMA2有助于锌和镉的转运,特别是对发育中的组织[85]。
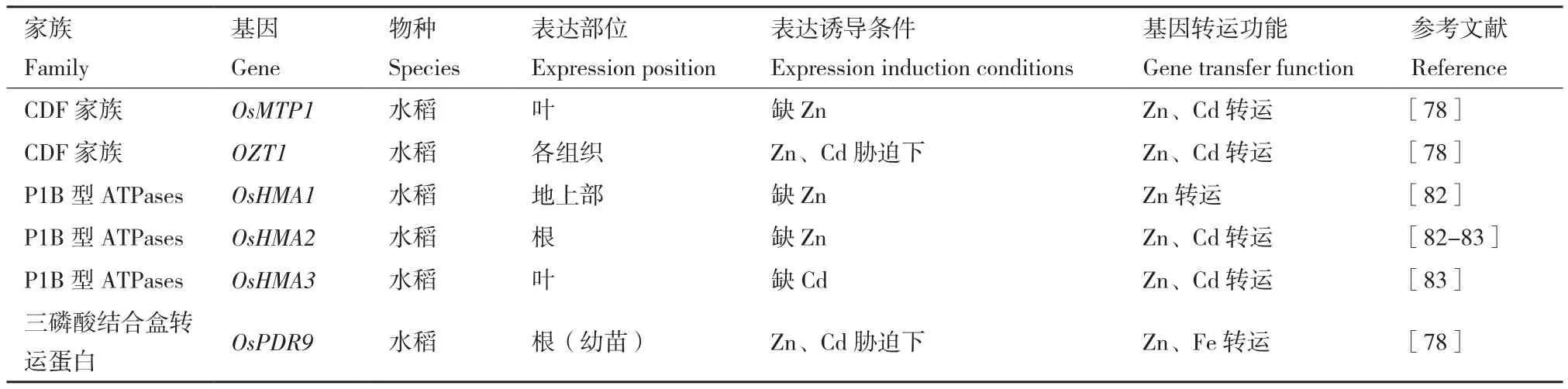
表4 锌排出蛋白种类、功能及表达特征Table 4 Types, functions and expression characteristics of zinc excretion proteins
6.2.3 三磷酸结合盒转运蛋白 OsPDR9是在水稻中发现的1种参与锌铁转运的三磷酸结合盒蛋白,在Zn和Cd诱导下,OsPDR9在水稻幼苗的根中表达[79]。图3为现阶段已发现水稻中6个锌排出蛋白成员的氨基酸序列比对,其亲缘关系如图4所示。

图3 锌排出蛋白氨基酸序列比对Fig.3 Zinc excretion protein amino acid sequence alignment
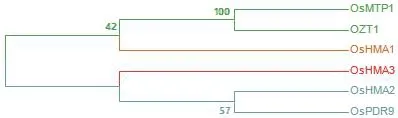
图4 锌排出蛋白进化树Fig.4 Zinc excretion protein evolutionary tree
6.3 锌转运相关蛋白基因克隆
现已有大量研究表明锌转运相关蛋白可参与锌的吸收转运及分配,深入研究其分子机制及其基因克隆,可为人们培育富锌水稻奠定理论基础。因此,研究学者对锌转运相关蛋白的基因克隆进行了深入研究。目前已克隆的锌转运相关蛋白基因有 OsZIP1、OsZIP3、OsZIP4、OsZIP5、OsZIP6、OsZIP8、OsIRT1、OsNRAMP3、OsMTP1、OsHMA2、OsHMA3、OsPDR9,基因分布如图5所示。
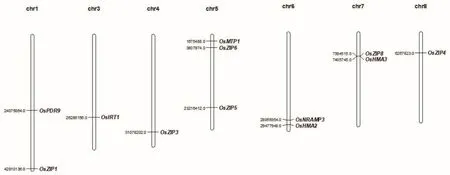
图5 已克隆锌转运蛋白基因分布图Fig.5 Distribution map of cloned zinc transporter
OsZIP1被认为是一种锌吸收转运体,也是一种限制水稻锌过量积累的金属外排转运蛋白,其表达是在缺锌的情况下诱导的[81]。OsZIP3主要在水稻根、叶处表达,在根中的表达受缺锌诱导[86]。OsZIP4、OsZIP6、OsZIP8、在水稻地上部和根中表达,其表达是在缺锌、铁的情况下诱导的[76-77]。OsIRT1缺铁时在水稻茎的韧皮部处表达[77]。OsHMA2、OsHMA3在缺铁、镉时分别在水稻根、叶处表达,都参与锌和镉的转运[82-83]。OsPDR9在锌、镉胁迫下在根及幼苗处表达[78]。
锌被水稻吸收后在根部积累,提高锌向茎叶转运效率成为关键。因此,对未克隆的相关基因(OsZIP2、OsZIP7、OsNRAMP2、OZT1、OsHMA1)进行功能分析将成为今后的研究重点。
7 展望
锌对促进水稻生长发育,保障优质高产等方面具有重要作用,水稻所需的锌主要从土壤中摄取,但由于我国水稻土资源丰富,类型复杂,不同种稻土中锌含量分布差异性较大,且不同基因型水稻对锌元素的吸收积累能力也不同,导致水稻对锌的吸收水平和籽粒富锌效果欠佳。因此今后的研究可以从以下几个方面加强:
目前,专家学者对于植物营养与肥料的研究已经实现从定性方面深入到定量方面,根据不同水稻土的含锌量和水稻的营养特点,制定测土施肥和植株诊断施肥方案,通过施肥定量的投入锌营养,调节水稻生长和发育[87]。数字模型的应用可以满足不同环境、不同生育期内水稻锌营养的需求,做到定量调控,提高锌肥利用效率,避免“奢侈吸收”和无效累积[88]。
2004年以来,中国作物营养强化项目(Harvest plus-China)一直将通过作物营养强化提高主要粮食作物中的微量营养元素含量(铁、锌、维生素A等)作为重要研究目标。但要实现这一目标,应从以下三个方面进行开展:一是集中人力、物力资源对现有水稻种质的锌吸收积累能力进行评价,为水稻资源的合理应用提供理论依据;二是利用现代分子生物技术选育不改变外观、口感和食味特性的锌营养高效型的水稻品种,提高水稻对锌的吸收积累,实现生物强化;三是通过提高稻米中锌的绝对含量和降低营养抑制物质(植酸)的水平,提高水稻锌的生物有效性,改善人类的锌营养[36]。
随着分子生物学的发展,越来越多参与水稻体内锌吸收和解毒作用的蛋白被相继发现,并对大多数锌转运蛋白家族成员的结构、功能、细胞定位和器官表达特异性等方面展开深入研究[77-80]。但仍存在以下问题:一是研究目标单一化,对多个锌吸收转运蛋白协同调控的研究较少。二是目前对水稻籽粒锌积累调控基因的研究过少,已有研究表明锌转运体基因的过表达,会使地上部的Zn2+向根部迁移,从而导致锌在籽粒富集减少,为了提高水稻的锌含量水平,若通过更换一个特异性的启动子,或发现一个组织特异性表达的基因来实现相关蛋白基因在籽粒中进行异位表达,达到籽粒富集锌的目的,将是今后研究锌转运蛋白的方向之一。三是发现的锌转运蛋白成员中克隆报道不多,对未克隆的相关基因进行功能分析,丰富水稻中锌元素吸收、转运和分配机制的基因调控网络,并为富锌水稻分子育种研究提供理论依据。
随着第二代测序技术的发展和鉴定技术的进步,全基因组关联分析(genome-wide association study,GWAS)因关联精度高,研究周期短必将成为发掘与水稻锌含量相关遗传变异位点的有效方法,获取与营养相关的关键位点将成为可能,也为系统全面的认知锌营养信号通路奠定了基础[89-90]。通过GWAS分析和转录组分析,充分挖掘富锌水稻种质资源,有助于了解水稻对相关性状适应性的途径和机制,也有助于设计和制定有效的育种策略,培育相关的富锌水稻品种。这些结果将为在育种中提高水稻籽粒锌含量和探索其遗传机理提供线索[91-92]。
猜你喜欢
现代农村科技(2022年10期)2022-12-06
现代畜牧科技(2021年4期)2021-12-05
小哥白尼(军事科学)(2019年9期)2019-12-21
河南农业·综合版(2019年6期)2019-09-10
河南农业(2019年6期)2019-06-05
电影(2019年3期)2019-04-04
河北农业科学(2018年2期)2018-07-26
种业导刊(2017年7期)2017-08-22
少儿科学周刊·少年版(2017年3期)2017-06-29
科学种养(2017年6期)2017-06-13
