
PCV2感染对猪淋巴细胞NF-κB信号通路及炎性细胞因子表达的动态影响
2022-07-15 00:49王艳萍徐倩倩苗立中王金良沈志强山东省滨州畜牧兽医研究院山东滨州256600山东绿都生物科技有限公司山东滨州256600
中国兽医学报 2022年6期
董 林,王艳萍,唐 娜,徐倩倩,苗立中,王金良,沈志强* (.山东省滨州畜牧兽医研究院,山东 滨州 256600;2.山东绿都生物科技有限公司,山东 滨州 256600)
猪圆环病毒2型(porcine circovirus 2,PCV2)是已知最小的单股DNA病毒,病毒结构为闭合环状,无囊膜,是断奶仔猪多系统衰歇综合征(post weaning multi-systemic syndrone,PMWS)的主要病原[1]。PCV2主要浸染免疫系统,损伤免疫细胞,导致机体免疫抑制的发生,感染仔猪的淋巴结、脾脏、扁桃体、胸腺等免疫器官呈现淋巴细胞的缺失、减少和单核巨噬细胞的浸润为主要特征的病理变化[2]。现阶段对相关因子分泌的相互影响和动态变化规律研究鲜见报道,特别是调控细胞因子分泌的细胞间信号通路还未有明确报道[3]。
核因子κB(nuclear factor κB,NF-κB)是一类参与促进多种基因转录的多向性核转录因子,属于Rel蛋白家族,主要包括P50、P52、c-Rel、RelA(P65)、RelB共5个家族成员,广泛存在于真核细胞中,是体内介导炎症反应的重要通路[4]。NF-κB含有300个氨基酸组成的Rel同源结构域(Rel homology domain,RHD),RHD通过其DNA结合区、二聚体化区域及核定位序列与含有κB序列的DNA结合,与同源或异源亚单位形成二聚体或者与IκB抑制蛋白相互作用调节核定位[5]。当细胞受到刺激时,可诱导IκB磷酸化启动降解途径,使得NF-κB易位进入细胞核内与DNA结合,实现转录调节[6-7]。
本试验选取PCV2感染的体外培养仔猪淋巴细胞,研究PCV2作用时间与NF-κB激活途径和炎症细胞因子表达之间关系,探索NF-κB的激活机制及其对细胞因子的影响,以期为PCV2感染防制提供理论支撑。
1 材料与方法
1.1 病毒PCV2 LVDU株(含病毒105.5TCID50/mL),由山东绿都生物科技有限公司提供。
1.2 实验动物28日龄PCV2抗原、抗体双阴性健康易感仔猪10头,由山东滨州禾川林丰畜牧科技有限公司养殖场提供。
1.3 主要试剂与仪器RRPMI-1640培养基(R8758-500,Gibco公司);小牛血清(20200145,天津康源生物);核蛋白提取试剂盒、BCA蛋白检测试剂盒购自北京天根生物;NF-κB EMSA试剂盒,化学发光液为美国Millpore公司产品;兔抗NF-κB/P65单克隆抗体、鼠抗猪p-IκBα单克隆抗体购自美国Cell Signaling Technology公司;TRITC-羊抗兔酶标抗体购自中杉金桥有限公司。恒温恒湿CO2培养箱(Heracell 150i,Thermo公司);高速冷冻离心机(ST16R,Thermo公司);蛋白电泳仪为(Mini-Protean Tetra,BioRad公司);化学发光成像系统(Fusion Solo,法国Vilber公司);激光共聚焦显微镜(A1si+A1Rsi+,日本Nikon公司)。
1.4 体外脾淋巴细胞培养及PCV2接毒10头经检测PCV2抗原、抗体双阴性的28日龄仔猪,无菌采集、分离脾脏细胞。用RPMI-1640培养液重悬细胞并计数,调整单层细胞浓度到6×106个/mL,在容积为250 mL细胞培养瓶中加入25 mL 制备的细胞悬液,分为对照组和PCV2感染组。感染组每毫升细胞悬液加PCV2病毒悬液200 μL;对照组加等量培养液。放入CO2培养箱中于37℃条件下培养,培养0,6,12,24,48 h收取悬浮的淋巴细胞和上清液,每个时间点各取淋巴细胞2 mL用于激光共聚焦检测,其余液氮速冻后置-70℃ 保存备用。细胞上清液用ELISA试剂盒检测分泌的细胞因子IL-1β、TNF-α、IL-6、IL-10、IL-17和TGF-β1的含量。
1.5 激光共聚焦检测NF-κB核移位取不同时间培养的细胞,用0.01 mol/L PBS(pH7.2)洗涤3次后重悬,取100 μL加入经对聚赖氨酸处理凹玻片上,室温静置15 min,浸入4%的多聚甲醛固定10 min,用0.5%TritonX-10+PBS室温通透;再用PBS洗5次,5%小牛血清封闭30 min,加入NF-κB/P65一抗37℃孵育45 min;洗涤后加入荧光二抗37℃孵育30 min,洗涤后加入核染液Hoechst,室温孵育5 min,洗涤5次后加入淬火液,于激光共聚焦显微镜观察结果(波长560 nm)。
1.6 Western blot检测NF-κB/P65、p-IκB蛋白分别提取体外培养仔猪淋巴细胞核蛋白和胞质蛋白,操作按Pierce核蛋白提取试剂盒操作说明书进行。提取蛋白用BCA法测定蛋白浓度,置于-70℃冰箱保存备用。
1.7 NF-κB与核蛋白中DNA结合活性检测依据EMSA试剂盒说明书进行操作,提取核蛋白进行非变性SDS-PAGE电泳,转移NC膜上,紫外灯下交联10 min,化学发光法检测生物素标记的DNA,LNS-4000成像系统进行测量和分析。
1.8 ELISA检测细胞培养液中细胞因子分泌蛋白含量取不同时间细胞上清液,进行IL-1β、TNF-α、IL-6、IL-10、IL-17和TGF-β1含量检测。具体操作按照对应ELSIA试剂盒说明书进行。制作标准曲线,样品检测后于450 nm酶标仪读取D值,依据标准曲线,计算得出各D值对应的样品蛋白含量。

2 结果
2.1 PCV2对体外培养淋巴细胞NF-κB/P65蛋白核移位的影响激光共聚焦显微镜观测结果(图1)显示,PCV2感染后6 h可观测到明显的核内NF-κB/P65蛋白存在,淋巴细胞中随着PCV2感染时间的增加,NF-κB/P65入核易位呈现不断增强的趋势。对照组仅在24 h才能观测到部分淋巴细胞核内出现少量NF-κB/P65蛋白存在。

A.PCV2接种后0 h; B.PCV2接种后6 h;C.PCV2接种后12 h;D.PCV2接种后24 h;E.PCV2接种后48 h。细胞核染色呈蓝色,胞质NF-κB/P65蛋白染色呈红色,易位核内染色合成呈粉红色图1 PCV2感染对体外培养淋巴细胞NF-κB核易位影响
Western blot检测结果如图2,随着PCV2作用体外培养仔猪淋巴细胞时间的延长,细胞核中NF-κB/P65蛋白含量呈现逐渐增多, PCV2作用6 h试验组细胞核中NF-κB/P65含量显著高于对照组(P<0.05),随着作用时间增加,试验组和对照组细胞核内NF-κB/P65均呈现增强趋势,在随后观测时间段内,试验组细胞核内NF-κB/P65含量均显著高于对照组,其中24和48 h极显著高于对照组(P<0.01)。说明PCV2感染,可导致体外培养淋巴细胞中NF-κB/P65发生核易位,并呈现明显的“时间-效应”累增关系。

A.Western blot检测p-IκBα含量结果;B.p-IκBα蛋白相对表达量。**.与对照组差异极显著(P<0.01);*.与对照组差异显著(P<0.05)。下同图2 PCV2感染对体外培养淋巴细胞核内NF-κB/P65蛋白表达影响(n=10)
2.2 PCV2对细胞核中NF-κB与DNA结合率的影响PCV2对体外培养淋巴细胞核蛋白NF-κB与DNA结合率的EMSA法检测结果见图3。体外培养淋巴细胞在PCV2感染条件下,随着作用时间的增加,NF-κB与DNA结合率呈现逐渐升高趋势,12 h 开始PCV2攻毒组结合率显著高于对照组(P<0.05),其中24和48 h极显著高于对照组(P<0.01),说明PCV2能促进易位的NF-κB与DNA结合,且呈现“时间累积效应”。

A.NF-κB与DNA结合EMSA检测结果(D.NF-κB与DNA结合带;F.无结合带);B.DNA结合活性结果图3 PCV2感染对体外培养仔猪淋巴细胞核中NF-κB与DNA结合率的影响(n=10)
2.3 PCV2对体外培养仔猪淋巴细胞质中p-IκBα蛋白含量的影响Western blot对体外培养仔猪淋巴细胞中p-IκBα含量进行检测,结果见图4。随着PCV2感染作用时间的增加,淋巴细胞质中p-IκBα蛋白呈现稳定增加,从培养6 h开始,淋巴细胞质中p-IκBα含量明显高于对照组,24 h开始极显著高于对照组,显示PCV2具有明显的促进胞质内κB磷酸化的作用。

A.Western blot检测p-IκBα含量结果;B.p-IκBα蛋白相对表达量图4 PCV2感染对体外培养仔猪淋巴细胞质中p-IκBα蛋白含量的影响(n=10)
2.4 细胞培养上清中分泌细胞因子含量检测
2.4.1细胞培养上清中分泌型IL-1β和IL-6含量检测 细胞培养上清中IL-1β和IL-6检测结果分别见图5~6。相对于对照组,PCV2感染组IL-1β在24和48 h含量均高有所升高,但与对照组差异不显著(P>0.05)。相对于对照组,PCV2感染组IL-6在24 h时增加明显,48 h感染组 IL-6含量显著高于对照组(P<0.05)。

图5 细胞培养上清中IL-1β含量检测结果

图6 细胞培养上清中IL-6含量检测结果
2.4.2细胞培养上清中分泌型IL-17和TNF-α含量检测 上清中IL-17和TNF-α含量结果见图7~8。PCV2攻毒后6 h,攻毒组细胞上清中IL-17含量明显升高,12和24 h极显著促进IL-17分泌(P<0.01),48 h时还明显高于对照组IL-17含量(P<0.05),结果显示PCV2攻毒随着时间延长可明显升高体外培养淋巴细胞分泌IL-17水平。PCV2攻毒后6 h,攻毒组TNF-α含量显著升高(P<0.05),12 h降至对照组水平,随后急剧增加,24 h达到峰值,48 h有所降低,但还极显著高于对照组(P<0.01)。

图7 细胞培养上清中IL-17含量检测结果

图8 细胞培养上清中TNF-α含量检测结果
2.4.3细胞培养上清中分泌型IL-10和TGF-β1含量检测 IL-10含量检测结果见图9,PCV2攻毒后,体外培养淋巴细胞从6 h开始分泌IL-10水平明显升高,且在随后的12,24和48 h均高于对照组水平,其中12 h达到峰值,显著高于对照组(P<0.05)。TGF-β1含量变化见图10,其变化趋势呈现先降低后升高的特征,6 h明显低于对照组,两者差异显著(P<0.05),12 h还明显低于对照组,24 h开始升高,48 h急剧增加,极显著高于对照组(P<0.01)。

图9 细胞培养上清中IL-10含量检测结果
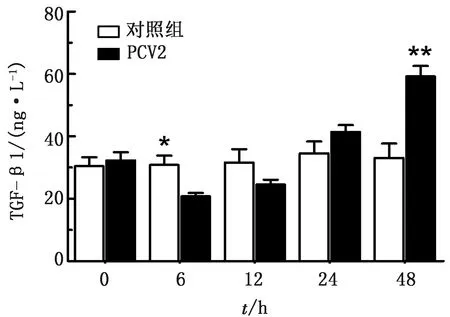
图10 细胞培养上清中TGF-β1含量检测结果
3 讨论
NF-κB作为重要的核转录调控基元,在炎性细胞因子激活与表达过程中起着重要的调控作用,并调节炎症反应最终进程。研究证实NF-κB信号活化与多种炎性细胞因子(IL-10、IL-6、TGF-β1和TNF-α)之间存在密切关联[8-10]。本研究发现,随着PCV2感染时间延长,NF-κB/P65的核易位强度不断增强,核内NF-κB/P65蛋白含量不断增强,呈现明显“时间-效应”关系。随着核内NF-κB/P65蛋白含量的增加,NF-κB与DNA的结合效率呈现明显增强。本试验证实,PCV2感染可导致体外培养淋巴细胞中NF-κB信号激活,表现明显的核易位转移和DNA结合活性增强,为调控下游相关基因的转录与表达提供条件,这一结果与吕春子等[11]的研究一致。WEI等[12]研究发现,NF-κB信号途径可以被PCV2有效激活,导致细胞核内P65蛋白含量和NF-κB与DNA结合活性增加,有利于病毒在细胞中的复制。PCV2能有效激活淋巴细胞中的NF-κB信号,易位至细胞核内与DNA调控元件结合,可有效调控细胞因子及其受体的表达,该途径可能是PCV2导致免疫抑制发生的重要途径[13]。
NF-κB激活通路存在“经典途径”和“旁路途径”2条不同途径[14]。“经典途径”通过促发胞质中IκB的磷酸化降解导致信号激活,调控基因的转录与表达,该途径主要参与机体调控炎性细胞因子、细胞凋亡等相关基因。本试验结果表明,在PCV2作用下,体外培养仔猪淋巴细胞胞质中p-IκB蛋白从6 h开始显著高于对照组(P<0.05),至24 h 极显著高于对照组(P<0.01),这种现象基本与NF-κB/P65蛋白入核和DNA结合活性同步,进一步证实PCV2是通过经典途径实现。
本试验结果表明,炎性因子IL-1β和IL-6在PCV2感染后含量变化不明显,仅IL-6在48 h显著高于对照组(P<0.05),这与陈耿等[15]的研究结果相一致。本试验结果显示,TNF-α和IL-17在PCV2作用 6 h开始明显升高,在24 h达到峰值,其中TNF-α表现出在12 h明显降低,这一结果与相关学者研究不一致,这可能与TNF-α和IL-17同时具有正调节免疫活性和产生炎症损伤的特点有关,早期的含量增加主要表现免疫调节,后期含量升高可能导致免疫损伤[16]。本试验中PCV2感染12 h出现IL-10明显的含量增加,提示PCV2可导致细胞免疫抑制,这与相关研究结果一致。在PCV2感染6,12 h时TGF-β1出现明显降低,这可能是感染PCV2后机体免疫反应增强,随后开始升高,48 h显著高于对照组,可能与机体免疫抑制发生存在一定关联。细胞因子含量检测进一步证实PCV2可以激活NF-κB信号通路,并导致炎性细胞因子表达出现相应变化。
综上,本研究从NF-κB的核易位、核蛋白NF-κB/P65含量变化,NF-κB与核内DNA结合效率以及炎性细胞因子分泌表达等4个方面评价了NF-κB细胞信号通路激活的主要特征,定量检测PCV2对体外培养仔猪淋巴细胞NF-κB信号通路激活及其对炎性细胞因子表达活性的影响,为PCV2感染、致病机理和免疫应答研究奠定了一定基础。
猜你喜欢
中国现代医生(2022年21期)2022-08-22
中国医药科学(2022年5期)2022-05-05
特别健康·下半月(2018年2期)2018-04-24
中国当代医药(2017年3期)2017-03-18
中国当代医药(2016年29期)2017-01-03
中国科技纵横(2016年20期)2016-12-28
中国实用医药(2016年24期)2016-10-17
祝您健康(1990年6期)1990-12-30
祝您健康(1981年4期)1981-12-29
