
PI3K-Akt-Rho A通路在猪血凝性脑脊髓炎病毒入侵N2a细胞中的调控作用
2022-07-23 07:33周倩宇吕晓玲王改丽贺文琦吉林大学动物医学学院吉林长春006山西农业大学动物医学学院山西晋中00800吉林省畜牧兽医科学研究院吉林长春006
中国兽医学报 2022年5期
周倩宇,吕晓玲,王改丽,李 姿,贺文琦* (.吉林大学 动物医学学院,吉林 长春 006;.山西农业大学 动物医学学院,山西 晋中 00800;.吉林省畜牧兽医科学研究院,吉林 长春 006)
猪血凝性脑脊髓炎病毒(PHEV)属于单链RNA病毒,是一种典型的嗜神经性冠状病毒,主要感染哺乳期仔猪,引起脑脊髓炎和/或消化系统疾病,以呕吐、衰竭及中枢神经系统障碍为特征[1-2]。该病毒攻击宿主中枢神经系统的神经细胞,但其神经损伤机制尚不清楚[3]。肌动蛋白(F-actin)是真核细胞骨架的重要组成部分,神经细胞的突触可塑性、胞质环流、神经递质传递及信号转导等均受其动态调节所驱动[4]。同时,细胞F-actin骨架重塑在防御病毒入侵、病毒胞内运输及病毒粒子释放过程中扮演重要的角色[5-7]。
磷脂酰肌醇3-激酶(PI3K)具有磷脂酰肌醇激酶活性和丝氨酸/苏氨酸激酶活性,通常为激活细胞整合素信号所必需的,参与特定病毒的入侵过程,如单纯疱疹病毒1型(HSV-1)、爱泼斯坦-巴尔病毒(EBv)和腺病毒(adenovirus)[8-11]。哺乳动物中的丝氨酸/苏氨酸激酶Akt又称蛋白激酶B,包括3种亚型Akt1、Akt2和Akt3,它们是PI3K信号转导通路中的关键分子。PI3K调节亚基可与细胞膜上的相应受体结合,对其催化亚基进行激活以催化PIP3形成,随后将Akt募集到细胞膜上而起作用[12-13]。同时,PI3K可通过下游Akt信号调控F-actin细胞骨架重塑过程[14]。尽管PHEV通过劫持整合素α5β1-黏着斑激酶(FAK)-丝切蛋白(CFL)信号调节神经细胞F-actin骨架已被证实[15],但PI3K信号通路是否在这一过程中起作用尚不明确。
PI3K-Akt通路的下游调控因子众多,其中Ras相关蛋白超家族研究较广泛。Ras蛋白与信号转导有关,Ras蛋白结合GDP时为失活态,结合GTP时为活化态。当细胞外存在信号分子时,Ras蛋白释放出自身的GDP并结合GTP,从而由失活态向活化态转变,最终实现将胞外信号向胞内传递。Ras相关蛋白超家族可分为3个亚家族Rho、Rac和Cdc42,它们可以利用鸟嘌呤核苷酸结合和水解循环发挥作用[16-18]。其中,小G蛋白Rho亚家族GTP酶成员A(Rho A)可以调节细胞骨架系统,特别是肌动蛋白CFL的功能[19],已经被广泛认可。基于此,本研究拟探究PI3K-Akt-Rho A信号通路是否参与PHEV入侵宿主细胞过程以及该通路在PHEV入侵过程中的调控机制,有助于进一步阐明PHEV诱导神经损伤的致病机制,并寻找有吸引力的治疗靶点。
1 材料与方法
1.1 细胞小鼠神经母细胞瘤细胞(N2a细胞,ATCC:CCL131)在DMEM(6%胎牛血清)中传代培养。
1.2 病毒PHEV CC14毒株(GenBank:MF083115.1)由吉林大学动物医学学院动物病理解剖实验室保存,在N2a细胞中传代繁殖。
1.3 主要试剂鼠源PHEV-N单克隆抗体,由本实验室提取并保存;Cofilin(3318)、p-cofilin(3313)、PI3K(4255)、p-PI3K(4228)、Akt(4685)、p-Akt(4060)等单克隆抗体购于CST公司;GAPDH(60004-1)、FITC-phalloidin F-actin(SA00003-2)等单克隆抗体购于Proteintech公司;PI3K抑制剂(LY 294002)、Rho A抑制剂(CCG-1423)等购于GLPBIO公司;RNAiso-plus(9109)、SYBR Green qPCR试剂盒等购于TaKaRa公司。
1.4 蛋白质免疫印迹PHEV(MOI=1)孵育N2a细胞2 h,并给予药物干预。细胞在RIPA缓冲液中裂解,12 000 r/min离心后,取上清液重悬于5×SDS-PAGE缓冲液中,并煮沸。可溶性细胞裂解产物在12% SDS-PAGE凝胶上分离,转移到0.45 μm孔径硝化纤维素膜上,用5%牛血清白蛋白封闭并孵育抗体。用ImageJ软件进行灰度值分析。
1.5 荧光实时定量PCR(RT-qPCR)利用RNAiso-plus试剂提取细胞总RNA,逆转录为cDNA,采用TaKaRa SYBR Green qPCR试剂盒进行RT-qPCR检测。特异性引物:PHEV-P1,5′-TCT GGG AAT CCT GAC GAG C -3′;PHEV-P2,5′-AGG CGC TGC AAC ACT TAC-3′;GAPDH-P1,5′-CTC AAC TAC ATG GTC TAC ATG TTC-3′;GAPDH-P2,5′-ATT TGA TGT TAG TGG GGT CTC GCT C-3′。
1.6 间接免疫荧光将N2a细胞接种于细胞爬片,用信号分子抑制剂干预细胞后,再用PHEV孵育1 h,用0.1 mol/L PBS洗涤细胞3次,去除细胞表面未结合的病毒,于培养箱培养。用4%多聚甲醛和冷甲醇固定细胞,0.5% Triton X-100透膜处理,5% BSA封闭。孵育FITC-phalloidin特异性标记F-actin或PHEV-N一抗,Hoechst染核后于激光扫描共聚焦荧光显微镜(Olympus FluoView FV1000)下观察。
1.7 统计学分析利用GraphPad Prism 5.0软件进行数据分析,2组间差异用双边非配对t-test检验,3组及以上组间差异用单因素方差分析检验,P<0.05为差异具有统计学意义。*示P<0.05,**示P<0.01,***示P<0.001。
2 结果
2.1 PHEV入侵N2a细胞激活PI3K-Akt信号通路如图1A所示,PHEV感染N2a细胞5 min时PI3K迅速发生磷酸化,随后其磷酸化水平下降,并在20 min后磷酸化作用消失。检测PI3K下游底物Akt活性发现,PHEV感染早期(5~15 min)Akt磷酸化水平较高,20 min后其活性逐渐下降(图1B)。利用灭活的PHEV感染细胞后,PI3K磷酸化水平并未见显著变化(图1C)。该结果表明,PHEV入侵N2a细胞早期可迅速激活PI3K-Akt信号通路。
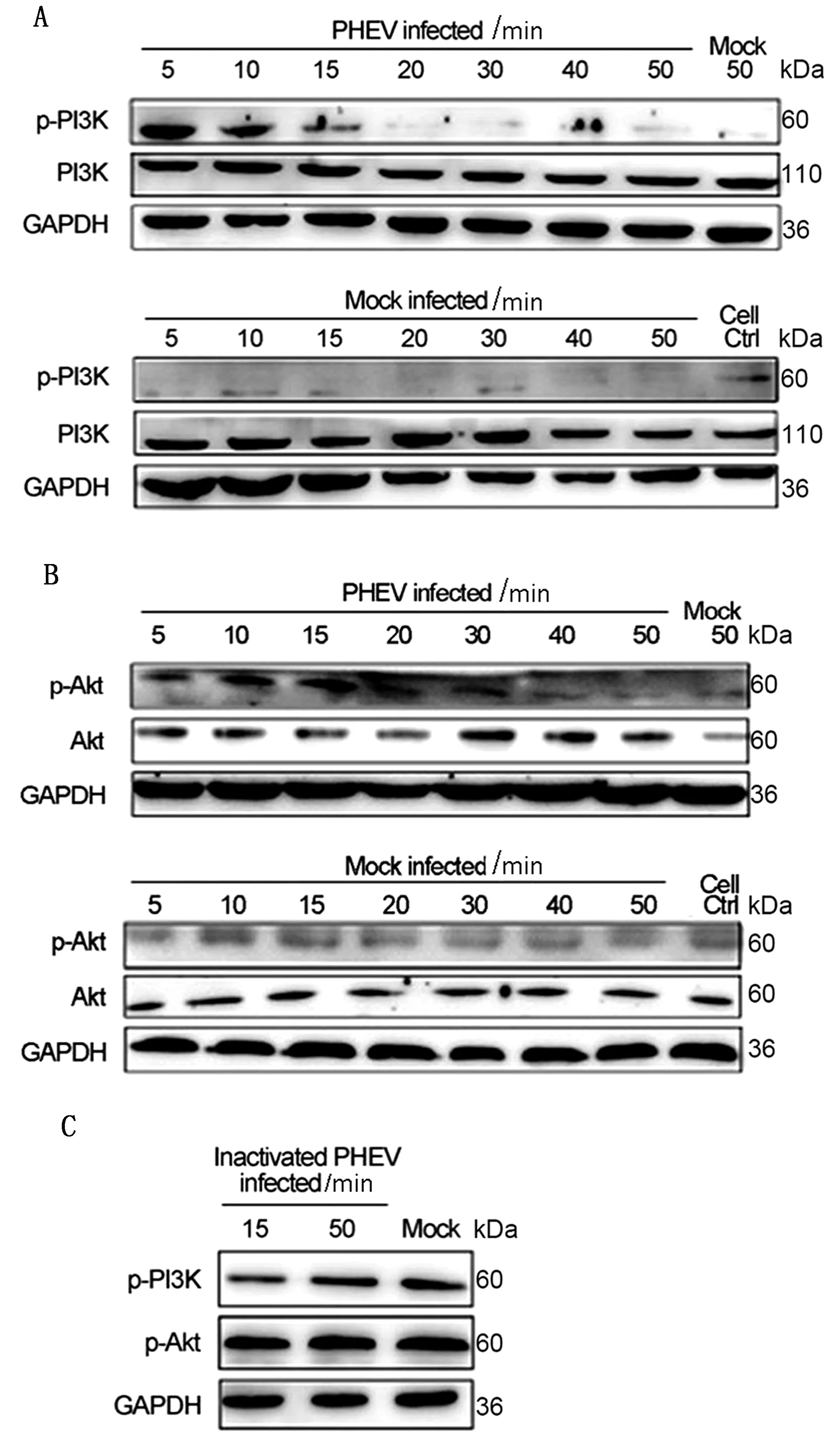
A,B.Western blot检测PHEV入侵N2a细胞后PI3K/p-PI3K及Akt/p-Akt蛋白水平;C.Western blot检测灭活PHEV入侵N2a细胞后p-PI3K和p-Akt蛋白水平图1 病毒感染时PI3K和Akt蛋白水平变化
2.2 PI3K信号失活抑制PHEV入侵N2a细胞为了检测PI3K-Akt信号通路对F-actin动态重塑及PHEV入侵的影响,使用PI3K特异性抑制剂LY294002(LY)预处理N2a细胞,然后接种PHEV 15 min。结果表明,PI3K抑制剂显著抑制PHEV蛋白表达(图2A),并呈剂量依赖性抑制PHEV感染效率(图2B)。免疫荧光标记研究表明,5 μmol/L LY可有效降低F-actin骨架结构的稳定性,同时抑制病毒内化作用(图2C)。上述研究证明,PI3K信号通路障碍可破坏F-actin骨架动态稳定性从而抑制PHEV入侵N2a细胞。
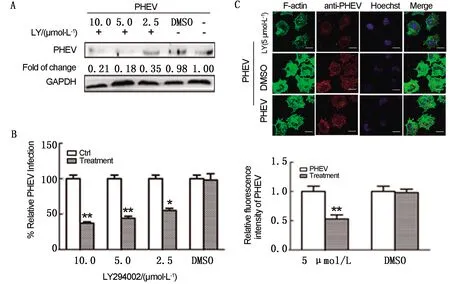
A.Western blot检测PHEV蛋白水平;B.RT-qPCR检测PHEV mRNA水平;C.共聚焦显微镜观察F-actin骨架及病毒入侵图2 PHEV入侵及F-actin骨架重塑
2.3 Rho A调控PHEV入侵N2a过程前期研究已证实,PHEV入侵N2a细胞过程中cofilin(CFL)磷酸化和去磷酸化呈动态变化,CFL活性的动态变化可以引起F-actin骨架发生一系列变化[20]。为了研究PI3K信号通路中Rho GTP酶成员Rho A对F-actin骨架重塑和PHEV入侵的影响,我们用Rho A特异性抑制剂CCG-1423(CCG)对N2a细胞进行预处理,并接种PHEV。如图3A所示,CCG-1423可使PHEV感染效率呈药物浓度依赖性降低。免疫荧光检测表明,1 μmol/L CCG-1423可降低病毒内化,并且降低F-actin骨架稳定性(图3B)。上述研究证明,在PHEV感染早期,增强Rho A信号可以调控PHEV入侵N2a细胞。
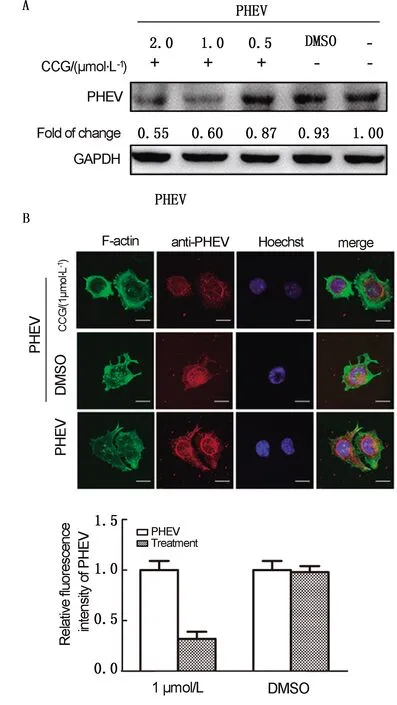
A.Rho A被抑制后p-CFL和PHEV的蛋白水平检测;B.共聚焦显微镜观察PHEV入侵图3 Rho A对F-actin骨架重塑和PHEV入侵的影响
3 讨论
前期研究工作已证实,PHEV入侵神经细胞可激活整合素α5β1-FAK-CFL信号通路,从而诱导细胞F-actin骨架动态重排[15]。PI3K对整合素信号敏感,且其效应底物Akt可以通过活化下游因子进一步调节细胞功能[21-22]。为了明确PHEV入侵N2a细胞时PI3K-Akt这一经典通路的激活状态,本研究进行了系统地检测。结果显示PI3K/p-PI3K和Akt/p-Akt在PHEV感染早期出现动态变化,提示PHEV有效入侵可以激活PI3K-Akt信号通路。结合PI3K抑制剂处理及F-actin骨架形态学观察,我们进一步证明PI3K-Akt信号通路可以调控PHEV入侵N2a细胞过程。
Rho A作为Rho家族中的一个成员,是调节细胞骨架重构的关键分子,激活后的Rho A可参与调节许多细胞形态和功能变化过程,包括细胞骨架的组装、转录因子的激活、细胞周期的调节和屏障功能的调节等[23]。因此,Rho A又被称为肌动蛋白细胞骨架和细胞形态异质性的分子开关。有研究证明PI3K-Akt信号调控F-actin骨架重塑过程与其下游Rho A信号密切相关[14,24]。因此Rho A信号分子
在PHEV入侵宿主细胞过程中可能发挥重要作用,本研究发现,抑制Rho A功能显著降低PHEV感染效率,表明在PHEV感染早期,增强Rho A信号可以调节PHEV入侵N2a细胞。
综上所述,本研究证实PHEV入侵能够激活PI3K-Akt信号通路,且Rho A作为该信号通路的下游调控因子参与调控F-actin骨架稳定性及PHEV入侵神经细胞过程。该研究为阐明PHEV诱导神经损伤性疾病的发病机制奠定了理论依据,并为抗病毒药物筛选提供了潜在靶标。
猜你喜欢
中国典型病例大全(2022年11期)2022-05-13
健康体检与管理(2022年4期)2022-05-13
医学概论(2022年4期)2022-04-24
中国药房(2022年7期)2022-04-14
电子乐园·上旬刊(2022年5期)2022-04-09
电子乐园·上旬刊(2022年5期)2022-04-09
发明与创新·大科技(2020年6期)2020-06-22
中国医药导报(2019年7期)2019-05-13
科教导刊·电子版(2018年9期)2018-06-07
分析化学(2018年2期)2018-03-02
