
尖刀唇石斛和翅梗石斛叶绿体全基因组分析
2022-08-16 06:25李卓蔚郎佳琪吴应梅杜慧慧
中草药 2022年16期
李卓蔚,邱 迁,郎佳琪,吴应梅*,杜慧慧,周 浓*
尖刀唇石斛和翅梗石斛叶绿体全基因组分析
李卓蔚1, 2,邱 迁1,郎佳琪1, 2,吴应梅1*,杜慧慧1,周 浓1, 2*
1. 重庆三峡学院 生物与食品工程学院,重庆 404120 2. 三峡库区道地药材绿色种植与深加工重庆市工程实验室,重庆 404120
测定尖刀唇石斛和翅梗石斛叶绿体基因组,分析其序列特征并鉴定石斛属植物物种间亲缘关系。利用Illumina Hisep 2 500测序平台对尖刀唇石斛和翅梗石斛进行二代测序,经组装、注释后得到2条完整的叶绿体全基因组序列,采用生物信息学方法分析其序列结构及石斛属的系统发育关系。尖刀唇石斛的叶绿体全基因组总长为159 786 bp,总GC含量为37.2%,共注释得到131个基因,包括88个蛋白编码基因、37个tRNA基因和6个rRNA基因;翅梗石斛的叶绿体全基因组总长为159 652 bp,总GC含量为37.1%,共注释得到131个基因,包括88个蛋白编码基因、37个tRNA基因和6个rRNA基因。尖刀唇石斛和翅梗石斛分别检测到112和127个简单重复序列(SSR),二者编码亮氨酸的密码子数量最多,编码色氨酸的密码子数量最少。系统发育树显示,尖刀唇石斛、反瓣石斛、翅梗石斛、梳唇石斛和长距石斛聚为一支,翅梗石斛与梳唇石斛和长距石斛的亲缘关系十分相近,支持率达到100%。对石斛属2种植物的叶绿体基因组结构和系统发育关系进行了分析,该研究结果将为石斛属药用植物的准确鉴定、开发利用及其资源保护提供科学依据。
尖刀唇石斛;翅梗石斛;药用植物;叶绿体全基因组;亲缘关系
石斛属Sw.为多年生草本植物,作为兰科(Orchidaceae)第2大属,在全球有1 500多个原生种,广泛分布于亚洲热带及亚热带地区[1]。产于中国的石斛属植物共有76种,包含2个变种,主要分布于西南和华南地区,尤其以云南省南部地区居多[2]。大部分石斛属植物具有极高的药用价值,主要以茎入药,作为传统滋补中药在中国和印度已有上千年的药用历史[3]。石斛主要含有多糖、酚类、萜类、生物碱等100多种化合物[4-5],通过对其药理活性的研究,证明了药材石斛具有增强免疫、降血糖、厚肠胃、护肝、抗肿瘤、抗氧化衰老、抗炎抑菌、抗血小板凝集、抗诱变、生津止渴、保护神经系统等功效[6-12]。此外,由于花色鲜艳、花枝优雅、花期长等特点,石斛常被作为一种观赏花卉,被喻为“四大观赏洋花”之一[13-14]。尖刀唇石斛Lindl.主要分布于云南腾冲、勐腊等地区[2],花色鲜艳,极具观赏价值,其茎常被作为药材使用,具有调脂、抗炎、抗氧化等功效[15-16];翅梗石斛Rchb. f主要分布于云南腾冲、思茅等地区[2],是一种药食同源的植物,具有抗肿瘤、抗凝血等功效[17-18]。近些年来,由于石斛的市场需求量剧增,大量尖刀唇石斛和翅梗石斛被采挖,导致其野生资源逐渐稀缺,目前已被列入《国家一级保护植物名录》和《世界自然保护联盟濒危物种红色名录》。虽然,不同种类的石斛作为药材时,常常被相互替代使用,但不同基原物种间的药用成分含量、药理活性及功效上仍具有很大的差异性。因此,能否对不同石斛植物正品和混淆品进行准确的鉴定,保证合理规范用药,该问题还有待解决。
石斛属植物种系间关系复杂,目前有关该属植物的分类和进化研究还存在较大争议,仅凭形态和解剖学特征分析已经难以满足属内物种的分类需求[19]。DNA条形码技术是一种常用的物种鉴定方法,采用保守的DNA片段,快速准确地对物种进行分析鉴定,此法在物种鉴定方面已被广泛采用[12, 20]。DNA通用条形码(ITS、K、L、H-A)虽然可以有效鉴定大部分石斛属植物[21-24],但基于目标基因的选择与数量等问题的影响,其分析结果往往存在较大分歧。随着测序技术的更新换代,叶绿体全基因序列的测定和比对为物种鉴定提供了新的参考[25]。叶绿体是多数自养生物特有的器官,具有独立的遗传物质,叶绿体基因组与核基因组和线粒体基因组相比,基因组较小,序列容易获得,且叶绿体基因组依赖于母系遗传,基因组成和结构比较保守[26]。与DNA通用条形码相比,叶绿体基因组具有更加丰富的遗传变异信息和更高的物种分辨能力,因此,以叶绿体全基因组为条形码将为石斛属这类亲缘关系复杂的物种提供更加准确有效的鉴定手段。目前,已有大量学者对不同石斛属植物叶绿体基因组的全长序列进行测定,并将多种石斛的叶绿体基因序列进行比对,探究其物种进化及其种间亲缘关系[25,27]。但对于尖刀唇石斛和翅梗石斛的叶绿体基因测定还未见报道。
本研究采用二代高通量测序技术对尖刀唇石斛和翅梗石斛的叶绿体全基因组进行测序、组装及注释,获得其叶绿体基因组的全长序列信息。采用生物信息学方法分析其序列特征,并与其他近缘物种的叶绿体基因序列进行比对,探讨石斛属物种间的亲缘关系,这也将为我国石斛药材正品和混淆品基原植物的分子鉴定提供参考依据。
1 材料
本研究中用于分子材料研究的尖刀唇石斛和翅梗石斛新鲜叶片均采自海拔为1 640 m的云南省保山市腾冲市(25°02′05.3″ N,98°49′09.7″ E)。样本保存于重庆三峡学院植物标本馆,样本凭证号为ZN15124、ZN15125,经重庆三峡学院周浓教授分类学鉴定为尖刀唇石斛Lindl.和翅梗石斛Rchb. f.。
2 方法
2.1 叶绿体全基因组的DNA提取与测序
将尖刀唇石斛和翅梗石斛新鲜叶片采用改良的CTAB法提取总DNA,具体参照江媛等[28]方法进行,通过琼脂糖凝胶电泳检测DNA质量,采用Illumina Hiseq 2500平台对检测合格的总DNA进行二代测序,测序工作由上海元莘生物医药科技有限公司完成。
2.2 叶绿体全基因组的组装、注释和提交
使用Trimmomatic v.0.32将序列中的低质量数据过滤掉,再使用GetOrganelle v1.6.4软件对过滤后的reads进行组装,得到高质量的contigs,并使用Circlator(V.2.0.1)软件检测reads映射的覆盖度和contigs各连接处,以检测组装的正确性。检测无误后,使用Cpgavas2软件对组装好的叶绿体全基因组进行注释,然后将注释完成的序列提交到GenBank数据库,获得尖刀唇石斛和翅梗石斛的登录号分别为OM049526和OL891650。使用在线工具OGDRAW(https:// chlorobox.mpimp-golm. mpg. de/OGDraw.html)绘制叶绿体全基因组的环状示意图。
2.3 叶绿体全基因组的简单重复序列及密码子偏好性分析
采用MISA在线工具(https://webblast. ipk-gatersleben.de/misa/)对叶绿体全基因组样本的简单重复序列位点(simple sequence repeat,SSR)进行检测,将参数设置为:单核苷酸重复次数≥10,二核苷酸重复次数≥5,三核苷酸、四核苷酸、五核苷酸、六核苷酸重复次数≥3,2个SSR之间的距离≥100 bp。采用CodonW软件对叶绿体全基因组样本进行密码子使用偏好性分析(relative synonymous codon usage,RSCU),参数参考系统默认值。
2.4 叶绿体全基因组序列比较分析
从NCBI(https://www.ncbi.nim.nih.gov)中下载8种分属于不同组的石斛属叶绿体全基因组序列,分别为长距石斛、束花石斛、梳唇石斛、反瓣石斛、景洪石斛、西畴石斛、重唇石斛和竹枝石斛,与尖刀唇石斛和翅梗石斛的叶绿体基因序列进行比较,分析不同石斛之间的序列差异性。使用在线软件IRscope(https://irscope.shinyapps.Io/irapp/)进行SC/IR边界收缩和扩张分析;使用DnaSP软件进行核苷酸变异性分析,窗口长度设置为600 bp,步长设置为200 bp,筛选出高变异位点;使用基因组在线分析软件mVISTA(http://genome.lbl. gov/vista/mvista)中的Shuffle-LAGAN模式,以尖刀唇石斛序列为参照进行全长序列分析。
2.5 叶绿体全基因组的系统发育分析
选择尖刀唇石斛及翅梗石斛在内的22条石斛属叶绿体全基因组序列,以兰属Sw.的碧玉兰Rchb. f.和虎头兰Rchb. f.作为外类群,使用MAFFT软件进行比对,将比对结果采用RaxML软件,基于最大似然法构建ML树,设置1000次重复以保证进化树稳定性,进而揭示石斛属植物物种间的亲缘关系[29]。
3 结果及分析
3.1 尖刀唇石斛和翅梗石斛的叶绿体基因组结构组成与基本特征
此次测序得到的尖刀唇石斛和翅梗石斛的叶绿体全基因组均为典型的双链环状四分体结构,如图1所示,由1个大单拷贝序列(large single copy,LSC),1个小单拷贝序列((small single copy,SSC)和一对反向重复序列(inverted repeat regions,IRs)组成。其中尖刀唇石斛序列全长为159 786 bp,LSC长度为87 477 bp,SSC长度为17 089 bp,IRs长度为27 610 bp,总GC含量为37.2%。翅梗石斛序列全长为159 652 bp,LSC长度为87 083 bp,SSC长度为18 451 bp,IRs长度为27 059 bp,总GC含量为37.1%(表1)。此外,尖刀唇石斛和翅梗石斛的LSC、SSC和IRs区的GC含量均存在差异,2种石斛GC含量均为IRs>LSC>SSC,其原因可能是在IRs区域存在含有高GC含量的核糖体RNA(rRNA)和转运RNA(tRNA)(表2)。
由表3可知,尖刀唇石斛和翅梗石斛均编码131个基因,包含88个蛋白质编码基因(CDS),37个tRNA和6个rRNA。除去IRs区重复基因,其中尖刀唇石斛有11个CDS基因(2、16、16、12、12、C1、F、B、A、D、B)和5个tRNA基因(E-UUCA-UGCV-UACL-UAAS-CGA)含有1个内含子,2个CDS基因(P3)含有2个内含子。翅梗石斛有9个CDS基因(2、16、16、C1、F、B、A、B、1)和5个tRNA基因(E-UUC、A-UGC、V-UAC、L-UAA、S-CGA)含有1个内含子,2个CDS基因(P和3)含有2个内含子。
3.2 尖刀唇石斛和翅梗石斛叶绿体基因组的简单重复序列及密码子使用情况分析
SSR的分析结果表明(表4):尖刀唇石斛叶绿体基因组中共含有112个SSR位点,分别包含20个单核苷酸、11个二核苷酸、70个三核苷酸、7个四核苷酸和4个五核苷酸,重复次数最多的为三核苷酸,其次为单核苷酸,二者共占比80.36%;翅梗石斛叶绿体基因组中共含有127个SSR位点,包含28个单核苷酸、14个二核苷酸、76个三核苷酸、6个四核苷酸、1个五核苷酸和2个六核苷酸,重复次数最多的为三核苷酸,其次为单核苷酸,二者占比为81.89%。总的来看,翅梗石斛的SSR位点多于尖刀唇石斛,但二者均以三核苷酸和单核苷酸重复居多,单核苷酸主要由A/T组成,其他重复序列也大都以A和T组成,表明二者在碱基组成过程中偏向使用碱基A和T。
图1 尖刀唇石斛、翅梗石斛的叶绿体基因组图谱
Fig. 1 Gene map of D. heterocarpum and D. trigonopus chloroplast genomes
表1 石斛属植物叶绿体基因组序列信息
Table 1 Chloroplast genome sequence information of Dendrobium
物种拉丁名GenBank登录号基因组大小/bpLSC/bpIRs/bpSSC/bp总基因数总GC含量/% 尖刀唇石斛D. heterocarpum Lindl.OM049526159 78687 47727 61017 08913137.2 翅梗石斛D. trigonopus Rchb. f.OL891650159 65287 08327 05918 45113137.1 报春石斛D. primulinum Lindl.NC_035321150 76784 44226 17513 97513237.6 杯鞘石斛D. gratiosissimum Rchb. f.LC192958151 82984 89026 29014 35913237.6 罗河石斛D. lohohense T. Tang et F. T. WangLC193516151 81284 87626 29214 35213237.6 长苏石斛D. brymerianum Rchb. f.LC192954151 83084 85526 29914 37713237.6 紫瓣石斛D. parishiiRchb. f.LC193518151 68984 70326 29514 39613237.6 束花石斛D. chrysanthumLindl.LC193514151 79084 75726 29614 44113237.6 肿节石斛D. pendulumRoxb.NC_029705153 24885 85226 39814 60012937.5 兜唇石斛D. aphyllum(Roxb.) C. E. FischerLC192953151 52484 58826 30814 32013237.6 大苞鞘石斛D. wardianumWarnerLC192961151 78884 83526 29614 36113137.6 长距石斛D. longicornuLindl.MN227146160 02487 47726 28919 96913237.1 反瓣石斛D. ellipsophyllumT. Tang et F. T. WangLC193519152 02684 93026 30414 48813237.5 景洪石斛D. exileSchltr.LC193522151 29484 36326 30814 31513237.7 球花石斛D. thyrsiflorumRchb. f.NC_047439151 68684 74926 29314 31512537.5 少花石斛D. parciflorumRchb. f. ex Lindl.LC193512150 07383 70826 27213 82113137.7 梳唇石斛D. strongylanthumRchb. f.NC_027691153 05985 83626 31614 59113037.6 西畴石斛D. xichouenseS. J. Cheng et C. Z. TangLC193520152 05284 98026 29314 48613237.5 小黄花石斛D. jenkinsiiLindl.LC193515151 71784 73426 28514 41313237.6 针叶石斛D. pseudotenellumGuillaum.NC_045854149 11484 26126 03312 78612537.4 重唇石斛D. hercoglossumRchb. f.LC192959151 93984 92426 30914 39713237.6 竹枝石斛D. salaccense(Bl.) Lindl.LC193510151 10484 27326 25814 31513237.3 碧玉兰Cymbidium lowianumRchb. f.NC_050990155 44784 18426 71017 84311936.8 虎头兰C. hookerianumRchb. f.NC_053268155 44784 18626 71117 83913536.8
表2 尖刀唇石斛、翅梗石斛叶绿体基因组碱基组成
Table 2 Base composition of chloroplast genome of D. heterocarpum and D. trigonopus
区域尖刀唇石斛翅梗石斛 碱基长度A/%T/%G/%C/%GC/%碱基长度A/%T/%G/%C/%GC/% LSC 87 47731.833.217.117.935.0 87 08331.833.417.017.834.8 SSC 17 08934.335.214.715.830.5 20 55534.134.815.016.231.1 IRa 27 61028.728.422.220.742.9 26 00728.328.320.922.443.4 IRb 27 61028.428.720.722.242.9 26 00728.328.320.922.443.4 总量159 78630.931.818.318.937.2159 65231.031.918.218.937.1
表3 尖刀唇石斛、翅梗石斛叶绿体基因组注释信息
Table 3 Annotation information of chloroplast genomes of D. heterocarpum and D. trigonopus
基因类别基因群基因代码基因名称 尖刀唇石斛翅梗石斛 遗传系统基因核糖体蛋白大亚基rplrpl2ab, rpl23a, rpl32, rpl22, rpl16b, rpl14, rpl36 , rpl20 , rpl33rpl2ab, rpl23a, rpl32, rpl22, rpl16b, rpl14, rpl36 , rpl20 , rpl33 核糖体蛋白小亚基rpsrps19a , rps7a, rps12b, rps12b, rps15, rps3 , rps8, rps11, rps18, rps4, rps14 , rps2, rps16brps19a , rps7a, rps12b, rps12b, rps15, rps3 , rps8, rps11, rps18, rps4, rps14 , rps2, rps16b RNA聚合酶rporpoA, rpoB , rpoC1b, rpoC2r rpoA, rpoB , rpoC1b, rpoC2 转运RNAtrntrnH-GUGa, trnM-CAUa, trnL-CAAa, trnV-GACa, trnE-UUCab, trnA-UGCab, trnR-ACGa, trnN-GUUa, trnL-UAG, trnP-UGG, trnW-CCA, trnM-CAU, trnV-UACb, trnF-GAA, trnL-UAAb, trnT-UGU, trnS-GGA, trnM-CAU, trnG-GCC, trnS-UGA, trnT-GGU, trnE-UUC, trnY-GUA, trnD-GUC, trnC-GCA, trnR-UCU, trnS-CGAb, trnS-GCU, trnQ-UUGtrnH-GUGa, trnM-CAUa, trnL-CAAa, trnV-GACa, trnE-UUCab, trnA-UGCab, trnR-ACGa, trnN-GUUa, trnL-UAG, trnP-UGG, trnW-CCA, trnM-CAU, trnV-UACb, trnF-GAA, trnL-UAAb, trnT-UGU, trnS-GGA, trnM-CAU, trnG-GCC, trnS-UGA, trnT-GGU, trnE-UUC, trnY-GUA, trnD-GUC, trnC-GCA, trnR-UCU, trnS-CGAb, trnS-GCU, trnQ-UUG 核糖体RNArrnrrn16Sa , rrn23Sa , rrn5Sarrn16Sa , rrn23Sa , rrn5Sa 光合系统基因光系统IpsapsaC, psaJ, psaI, psaA, psaBpsaC, psaJ, psaI, psaA, psaB 光系统IIpsbpsbH, psbN, psbT, psbB, psbE, psbF, psbL, psbJ, psbZ, psbC, psbD, psbM, psbI, psbK, psbApsbH, psbN, psbT, psbB, psbE, psbF, psbL, psbJ, psbZ, psbC, psbD, psbM, psbI, psbK, psbA ATP合成酶atpatpB , atpE, atpI, atpH, atpFb, atpAatpB , atpE, atpI, atpH, atpFb, atpA NADH脱氢酶ndhndhBab, ndhH, ndhAb, ndhI, ndhG, ndhE, ndhD, ndhF, ndhC, ndhK,ndhJndhBab, ndhH, ndhAb, ndhI, ndhG, ndhE, ndhD, ndhF, ndhC, ndhK,ndhJ 细胞色素b/f复合体petpetDb , petBb, petG, petL, petA , petNpetD , petBb, petG, petL, petA , petN Rubisco亚基rbcrbcLrbcL 其他基因成熟酶matmatKmatK 蛋白酶clpclpPcclpPc 翻译起始因子infinfAinfA 细胞色素c合成酶ccsccsAccsA 包膜蛋白cemcemAcemA 乙酰辅酶A羧化酶accaccDaccD 未知功能蛋白ycfycf2a , ycf15a , ycf1, ycf4 , ycf3cycf2a , ycf15a , ycf1b, ycf4 , ycf3c
a代表该基因位于反向重复区,b代表该基因含有1个内含子,c代表该基因含有2个内含子
ameans that the gene is located in the inverted repeat region,bmeans that the gene contains one intron, andcmeans that the gene contains two introns
表4 尖刀唇石斛、翅梗石斛叶绿体基因组SSR信息统计
Table 4 SSR information statistics of chloroplast genomes of D. heterocarpum and D. trigonopus
核苷酸类型重复序列尖刀唇石斛翅梗石斛 数量占比/%数量占比/% 单核苷酸A/T2017.862620.47 C/G−−21.57 二核苷酸AG/CTAT/AT291.798.042121.579.45 三核苷酸AAC/GTTAAG/CTTAAT/ATTACT/AGTAGC/CTGAGG/CCTATC/ATGACC/GGT1029134347−8.9325.8911.613.572.683.576.25−103116334727.8724.4112.602.362.363.155.511.57 四核苷酸AAAG/CTTTAAAT/ATTTAATT/AATTACAG/CTGTAGAT/ATCT221111.791.790.890.890.89121110.791.570.790.790.79 五核苷酸AAAAT/ATTTTAAATC/ATTTGAAATT/AATTT2111.790.890.89−1−−0.79− 六核苷酸AAGTAG/ACTTCTAGATGG/ATCTCC−−−−110.790.79
密码子使用偏好性又称相对同义密码子使用度,表示对于某一特定密码子在编码对应氨基酸的同义密码子之间的使用频率。当RSCU>1时,表示该密码子使用频率较高;当RSCU=1时,表示该密码子没有偏好性;当RSCU<1时,表示该密码子使用频率较低。由表5可知,尖刀唇石斛共编码53 262个氨基酸,其中编码亮氨酸(Leu)的密码子数量最多,达到5592个,占比10.50%,编码色氨酸(Trp)的密码子数量最少,仅有733个,占比1.38%;翅梗石斛共编码53 217个氨基酸,数量最多为编码亮氨酸的密码子,达到5560个,占比10.45%,数量最少为编码色氨酸密码子,仅有718个,占比1.35%。RSCU分析结果表明,在尖刀唇石斛的64种密码子中,RSCU大于1的密码子有31种,其中28种以A/U结尾,3种以G/C结尾,RSCU最大值为2.19,为编码精氨酸(Arg)的AGA,最小值为0.42,为编码精氨酸的CGC。在翅梗石斛的64种密码子中,RSCU大于1的密码子有31种,其中28种以A/U结尾,3种以G/C结尾,RSCU最大值为2.18,为编码精氨酸的AGA,最小值为0.44,为编码精氨酸的CGC。二者比较分析可以看出,尖刀唇石斛和翅梗石斛的密码子使用偏好性有细微的差别,但总体上大致相同。二者均以编码亮氨酸的密码子数量最多,编码色氨酸的密码子数量最少,偏好性最强的密码子均为AGA,最弱的均为CGC,且RSCU大于1的密码子几乎都已A/U结尾,表明2种石斛属植物均偏好使用A/U结尾的密码子,这与SSR分析结果一致。
3.3 10种石斛属植物叶绿体全基因组的SC/IR边界变化分析
将尖刀唇石斛、翅梗石斛、长距石斛、束花石斛、梳唇石斛、反瓣石斛、景洪石斛、西畴石斛、重唇石斛和竹枝石斛在内的10种石斛属植物的叶绿体基因序列的SC/IR边界进行分析(图2)。结果显示,石斛属植物叶绿体全基因组均存在4个边界,即为LSC-IRb、SSC-IRb、SSC-IRa、LSC-IRa。其中LSC-IRb边界较为保守,大部分位于22基因编码区内,仅尖刀唇石斛位于22基因右侧非编码区;SSC-IRb边界存在较大的分化,尖刀唇石斛和翅梗石斛位于F基因编码区内,左侧缺失1基因,梳唇石斛和重唇石斛均位于F基因左侧非编码区内,且梳唇石斛边界左侧缺失1基因,长距石斛的1和F基因均缺失,边界位于N基因右侧非编码区,景洪石斛边界位于1和F基因交界处,其余石斛均位于1基因和F基因重叠区,且向F基因编码区内扩展;SSC-IRa边界大部分位于1基因内侧,只有长距石斛1基因缺失,边界位于15和N基因交界处;LSC-IRa边界大部分位于22基因和A基因之间的非编码区,并偏向于靠近22基因,尖刀唇石斛、翅梗石斛和梳唇石斛缺少22基因,边界位于19和A基因之间的非编码区。总体上看,石斛属植物叶绿体全基因序列的4个边界中,SSC-IR边界变化较大,LSC-IR边界则较为保守。尖刀唇石斛和翅梗石斛的边界变化相似,长距石斛边界区域变化较大。
3.4 石斛属植物叶绿体基因组变异分析
以尖刀唇石斛叶绿体全基因组为参考序列,使用mVISTA在线工具将10种石斛的叶绿体基因组进行全长序列比对(图3)。结果表明:长距石斛叶绿体基因的变异性大,而其余石斛的基因序列则较为保守。整体上看,变异多发生在LSC区和SSC区,而且在相邻基因的间隔区,发生变异的机率更大,而IR区的变异程度整体较低,绝大部分基因序列的相似度均保持在较高水平。
表5 尖刀唇石斛、翅梗石斛叶绿体基因组密码子使用情况
Table 5 Codon usage in chloroplast genome of D. heterocarpum and D. trigonopus
氨基酸密码子尖刀唇石斛翅梗石斛氨基酸密码子尖刀唇石斛翅梗石斛 数量RSCU数量RSCU数量RSCU数量RSCU AlaGCA4651.194871.22ProCCA7641.287451.26 GCC3490.893690.93CCC5880.995770.97 GCG2370.612400.60CCG3660.613750.63 GCU5151.324971.25CCU6681.126761.14 CysUGC3900.704400.81GlnCAA10681.3611021.38 UGU7291.306511.19CAG4970.644990.62 AspGAC4350.554270.54ArgAGA12342.1912132.18 GAU11341.4511641.46AGG6001.065711.03 GluGAA14751.4014261.39CGA5791.035661.02 GAG6260.606330.61CGC2350.422470.44 PheUUC15820.7416030.84CGG3630.643620.65 UUU21591.1522051.16CGU3710.663820.69 GlyGGA8421.507971.43SerAGC4390.524020.50 GGC3180.573620.65AGU7340.876690.84 GGG5571.005310.96UCA9951.189691.21 GGU5210.935340.96UCC9331.118981.12 HisCAC3950.584010.59UCG5780.695750.72 CAU9571.429581.41UCU13801.6412921.61 IleAUA15461.0216321.06ThrACA6861.236781.19 AUC12120.8011560.75ACC5280.945410.95 AUU18041.1918351.19ACG3640.653590.63 LysAAA20851.2921151.30ACU6601.187001.23 AAG11360.7111310.70ValGUA7271.287501.27 LeuCUA8870.958910.96GUC4200.744120.70 CUC7120.766830.74GUG4070.724370.74 CUG5430.585480.59GUU7211.277671.30 CUU12201.3112331.33TrpUGG7331.007181.00 UUA11161.2011001.19TyrUAC6950.626950.60 UUG11141.2011051.19UAU15351.3816081.40 MetAUG8741.009251.00TerminaterUAA10731.0810581.09 AsnAAC6910.566890.56UAG8640.878230.85 AAU17831.4417601.44UGA10481.0510231.06

图2 10种石斛属植物叶绿体基因组的IR/SC边界变化情况
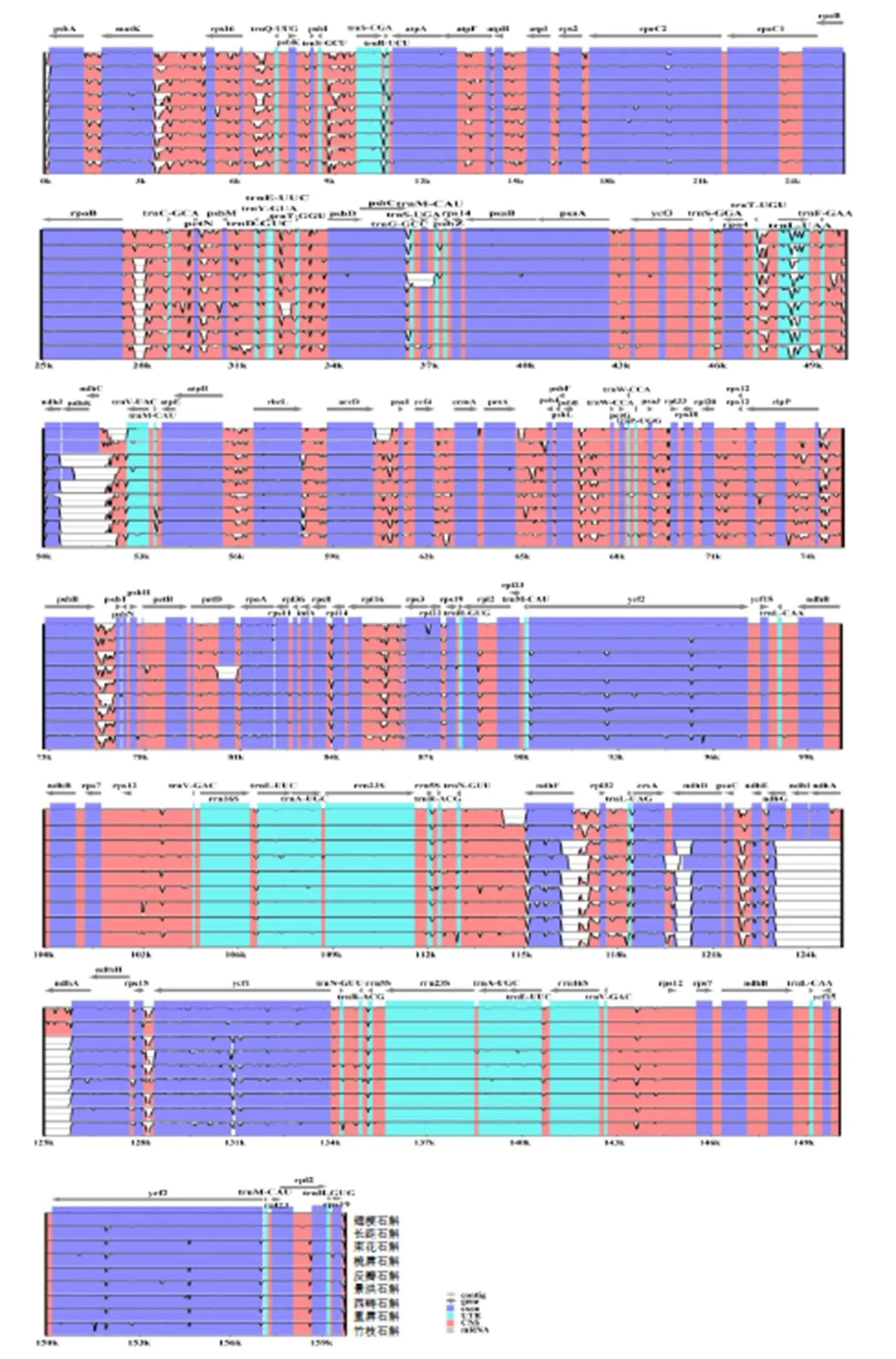
图3 石斛属植物叶绿体基因组全局比对分析
使用DnaSP6软件对10种石斛属药用植物叶绿体基因组高变异位点进行筛选分析(图4)。结果表明:LSC区的变异性整体较高,IR区的核苷酸变异性显著低于LSC区和SSC区,表明IR区为叶绿体基因组中最为保守的区域,且变异较多发生在相邻基因间隔区,这与全长序列比对分析结果保持一致。此外,还筛选出来4个高变异片段,分别为:J-V-UAC、P-B、16、32-L-UAG。
3.5 石斛属植物叶绿体基因序列的系统发育分析
以碧玉兰和虎头兰为外类群,使用MAFFT软件对22条石斛属植物的叶绿体基因序列进行多重比对,基于比对结果建立ML系统发育树(图5),分支节点上的数字代表该分支的可信度。结果表明,在石斛属这一大类分支中,尖刀唇石斛、反瓣石斛、翅梗石斛、梳唇石斛和长距石斛聚为一支,且支持率高达96%,其次才与其他石斛相聚,其中翅梗石斛与梳唇石斛和长距石斛的亲缘关系十分相近,支持率达到100%,竹枝石斛与其他石斛的亲缘关系相对较远。

图4 10种石斛属植物叶绿体基因组核苷酸变异性分析

图5 基于叶绿体基因组构建石斛属及其他植物的ML树
4 讨论
4.1 叶绿体基因序列特征
对比尖刀唇石斛和翅梗石斛的叶绿体全基因组发现,二者同为石斛属植株,在序列构成上差异不大,翅梗石斛的序列全长稍短于尖刀唇石斛,可能是由基因缺失所引起,二者在遗传系统基因组成上差异较小,其他功能基因组成也相对保守。将二者与石斛属其它叶绿体基因序列比较可以看出,大多数石斛叶绿体基因在总基因数、总GC含量等方面都与尖刀唇石斛和翅梗石斛相近,且较为保守。有研究表明,叶绿体基因组IR/SC边界区域的变异是叶绿体基因组结构变异的主要驱动力[30],其边界的收缩与扩张对叶绿体基因组的演化影响深远[31-32]。从对10种石斛属植物的叶绿体基因组IR/SC边界分析结果可以看出,SSC/IR边界区域差异性较大,而在通常认为比较保守的LSC/IR区域也存在细微的差异,其中长距石斛的差异性尤其明显。
大多数植物的叶绿体全基因组都富含SSR位点,SSR位点具有可稳定遗传、多态性广泛、丰度高、分布广等特点,因此常被用作物种鉴定、遗传多样性分析及分子标记辅助育种等方面[33-35]。通过对尖刀唇石斛和翅梗石斛的SSR位点检测结果可以看出,翅梗石斛的SSR位点多于尖刀唇石斛,且二者的重复序列存在较大的变化,因此可以看出石斛属种间叶绿体基因组的SSR多态信息含量丰富,可为石斛属植物种间鉴别提供参考依据[27]。总体上对尖刀唇石斛和翅梗石斛的SSR位点信息和密码子偏好性信息进行分析可以看出:二者均以三核苷酸重复居多,其次为单核苷酸,且具有碱基偏好性,偏好使用A/T碱基。偏好性最大的密码子均为AGA,且RSCU大于1的大都以A/U结尾,这与二者叶绿体全基因组富含A/T碱基的结果相一致,这与杜致辉等[25]及朱斌等[27]对黑喉石斛和球花石斛的分析结果相符,这种现象还可能广泛存在与其他石斛属植物中。McCoy等[36]和Niu等[37]研究表明,A/T碱基中氮原子含量明显少于G/C碱基,因此,在碱基突变过程中,A/T碱基消耗的能量比G/C少,这种优势就导致了SSR和密码子中A/T碱基偏好性的结果。此外,Mukhopadhyay等[38]证明了这种碱基偏好性还与叶绿体基因组各分区的变异程度相关联。从对10种石斛属植物的核苷酸变异性结果分析可以发现,GC含量较高的IR区变异程度显著低于GC含量较低的LSC区和SSC区,因此可以推断,碱基偏好性可能与序列的变异程度呈正相关。
4.2 石斛属植物的物种鉴定
石斛属植物在全球约有1500个种,产于中国的共有76种[39],大部分石斛种间表型较为相似,在相同生境中,不同种类的石斛之间可以相互杂交[40],因此,造成石斛属植株之间亲缘关系较为复杂,难以依据简单的形态和解剖学特征将其准确区分。叶绿体基因序列与线粒体基因序列和核基因序列相比,基因组小且结构较为保守,更加适合作为物种鉴定的依据,与DNA通用条形码相比,叶绿体基因全长序列具有更加丰富的遗传信息,可以更加准确地鉴定物种。随着测序技术的飞跃发展,叶绿体基因测序技术逐渐成熟,测序成本也在不断降低,因此,以叶绿体全基因组对物种进行分析的手段已被广泛采用,目前,已有大量的石斛属植物叶绿体全基因序列被测得,这也使得基于叶绿体基因组探究石斛亲缘关系成为可能。本研究以2种兰属植物作为外类群,将其与22种石斛属植物的叶绿体全基因序列结合,构建ML树。建树结果可以看出:石斛属种间近缘关系的支持率较高,石斛组的尖刀唇石斛与心叶组的反瓣石斛、黑毛组的翅梗石斛和长距石斛以及草叶组的梳唇石斛聚为一支,表明这五者亲缘关系较近,这与形态学分类的结果相违背[2],因此,对于石斛属这类种系关系复杂的物种而言,仅凭简单宏观的形态特征,还无法将其准确分类,还应在分子水平上对它们进行更加准确地定义。
本研究采用生物信息学方法对已注释完成的尖刀唇石斛和翅梗石斛叶绿体全基因组进行了全方位比对分析。总体上看,尖刀唇石斛和翅梗石斛与其他石斛的叶绿体全基因序列相似度较高,通过基因组成、IR边界比对、全长序列比对及ML建树所得的结果可以看出,石斛属植株叶绿体基因序列大体相似,但不同种类的叶绿体基因序列之间都会存在微小的变化,而这些变化将作为物种鉴定强有力的依据。通过对尖刀唇石斛和翅梗石斛的叶绿体基因序列进行比较分析,揭示了石斛属间物种亲缘关系,为石斛属药用植物的准确鉴定、开发利用及其资源保护提供了科学依据。
利益冲突 所有作者均声明不存在利益冲突
[1] Cakova V, Bonte F, Lobstein A.:Sources of active ingredients to treat age-related pathologies [J]., 2017, 8(6): 827-849.
[2] 张志耘. 中国植物志(第19卷) [M]. 北京:科学出版社, 1990: 125.
[3] 张雪琴, 赵庭梅, 刘静, 等. 石斛化学成分及药理作用研究进展 [J]. 中草药, 2018, 49(13): 3174-3182.
[4] Lam Y, Ng T B, Yao R M,. Evaluation of chemical constituents and important mechanism of pharmacological biology inplants [J]., 2015, 2015: 841752.
[5] 王宪楷, 赵同芳. 石斛属植物的化学成分与中药石斛 [J]. 中国药学杂志, 1986, 21(11): 666-669.
[6] Teixeira da Silva J A, Ng T B. The medicinal and pharmaceutical importance ofspecies [J]., 2017, 101(6): 2227-2239.
[7] Ng T B, Liu J Y, Wong J H,. Review of research on, a prized folk medicine [J]., 2012, 93(5): 1795-1803.
[8] Yoo S R, Jeong S J, Lee N R,. Simultaneous determination and anti-inflammatory effects of four phenolic compounds in[J]., 2017, 31(24): 2923-2926.
[9] 令狐楚, 谷荣辉, 秦礼康. 金钗石斛的化学成分及药理作用研究进展[J]. 中草药, 2021, 52(24): 7693-7708.
[10] 宋广青, 刘新民, 王琼, 等. 石斛药理作用研究进展 [J]. 中草药, 2014, 45(17): 2576-2580.
[11] 颜美秋, 陈素红, 吕圭源. 石斛“厚肠胃”相关功效药理学研究及应用进展 [J]. 中草药, 2016, 47(21): 3918-3924.
[12] 税小红, 税璘, 牛曼思, 等. 金钗石斛破壁粉对环磷酰胺致免疫低下小鼠免疫功能的调节作用[J]. 药物评价研究, 2018, 41(12): 2189-2194.
[13] 黄海, 李劲松, 符岸军, 等. 石斛属植物DNA条形码序列的筛选 [J]. 热带作物学报, 2010, 31(10): 1769-1777.
[14] 任羽, 尹俊梅, 杨光穗. 海南石斛属植物亲缘关系的SRAP分析 [J]. 热带作物学报, 2008, 29(6): 767-770.
[15] 颜莎. 尖刀唇石斛化学成分及降脂活性天然产物研究 [D]. 昆明: 云南大学, 2019.
[16] 杨晓蓓, 颜莎, 胡江苗, 等. 尖刀唇石斛化学成分研究 [J]. 天然产物研究与开发, 2019, 31(10): 1745-1752.
[17] 张婷, 张朝凤, 王峥涛, 等. 翅梗石斛的化学成分研究 [J]. 中国天然药物, 2005, 3(1): 28-30.
[18] 王彦兵, 周侯光, 尹红星, 等. 黑毛组6种石斛药效成分分析及营养价值评价 [J]. 天然产物研究与开发, 2020, 32(1): 95-102.
[19] Freudenstein J V, Rasmussen F N. What does morphology tell us about orchid relationships? —A cladistic analysis [J]., 1999, 86(2): 225-248.
[20] 钟志敏. 石斛DNA条形码鉴定及系统分类研究 [D]. 广州: 广州中医药大学, 2018.
[21] 栗丹, 李振坚, 毛萍, 等. 基于ITS序列石斛材料的鉴定及系统进化分析 [J]. 园艺学报, 2012, 39(8): 1539-1550.
[22] 刘静, 何涛, 淳泽. 药用石斛的叶绿体matK基因序列分析及鉴别 [J]. 药学学报, 2009, 44(9): 1051-1055.
[23] 袁佐清, 张建勇, 刘涛. 石斛属植物rbcL基因序列变异和系统发育初步研究 [J]. 时珍国医国药, 2009, 20(7): 1836-1837.
[24] 邵世光, 韩丽, 马艳红, 等. 枫斗类石斛cpDNA psbA-trnH的序列分析与鉴别 [J]. 药学学报, 2009, 44(10): 1173-1178.
[25] 杜致辉, 杨澜, 张朝君, 等. 黑喉石斛叶绿体基因组特征及比较分析 [J]. 热带作物学报, 2021, 42(11): 3111-3119.
[26] Wicke S, Schneeweiss G M, dePamphilis C W,. The evolution of the plastid chromosome in land plants: Gene content, gene order, gene function [J]., 2011, 76(3/4/5): 273-297.
[27] 朱斌, 甘晨晨, 王洪程. 球花石斛()叶绿体基因组特征及亲缘关系解析 [J]. 生物技术通报, 2021, 37(5): 38-47.
[28] 江媛, 杨青淑, 王婧, 等. 毛重楼叶绿体基因组序列特征及其系统发育分析 [J]. 中草药, 2021, 52(13): 4014-4022.
[29] 胡海粟, 张德全. 几种滇产龙胆属药用植物的DNA超级条形码研究 [J]. 中国中药杂志, 2021, 46(20): 5260-5269.
[30] Kim K J, Lee H L. Complete chloroplast genome sequences from Korean ginseng (Nees) and comparative analysis of sequence evolution among 17 vascular plants [J]., 2004, 11(4): 247-261.
[31] Wang W C, Chen S Y, Zhang X Z. Whole-genome comparison reveals divergent IR borders and mutation hotspots in chloroplast genomes of herbaceous bamboos (Bambusoideae: Olyreae) [J]., 2018, 23(7): 1537.
[32] Park S, An B, Park S. Reconfiguration of the plastid genome in: IR boundary shifting, inversion, and intraspecific variation [J]., 2018, 8(1): 13568.
[33] 丁鸽, 张代臻, 丁小余. 石斛资源分子水平研究进展 [J]. 中国实验方剂学杂志, 2018, 24(4): 208-215.
[34] Powell W, Morgante M, McDevitt R,. Polymorphic simple sequence repeat regions in chloroplast genomes: Applications to the population genetics of pines [J]., 1995, 92(17): 7759-7763.
[35] Pugh T, Fouet O, Risterucci A M,. A new cacao linkage map based on codominant markers: Development and integration of 201 new microsatellite markers [J]., 2004, 108(6): 1151-1161.
[36] McCoy S R, Kuehl J V, Boore J L,. The complete plastid genome sequence of: An unusually compact plastome with accelerated divergence rates [J]., 2008, 8: 130.
[37] Niu Z T, Xue Q Y, Wang H,. Mutational biases and GC-biased gene conversion affect GC content in the plastomes ofgenus [J]., 2017, 18(11): E2307.
[38] Mukhopadhyay P, Basak S, Ghosh T C. Nature of selective constraints on synonymous Codon usage of rice differs in GC-poor and GC-rich genes [J]., 2007, 400(1/2): 71-81.
[39] 李清, 李标, 郭顺星. 兰科石斛属植物分子生物学研究进展 [J]. 中国中药杂志, 2016, 41(15): 2753-2761.
[40] 黄捷. 石斛属植物交配亲和性研究 [D]. 广州: 华南农业大学, 2016.
Sequence analysis of complete chloroplast genome of Dendrobium heterocarpum and
LI Zhuo-wei1, 2, QIU Qian1, LANG Jia-qi1, 2, WU Ying-mei1, DU Hui-hui1, ZHOU Nong1, 2
1. School of Biology and Food Engineering, Chongqing Three Gorges University, Chongqing 404120, China 2. Chongqing Engineering Laboratory for Green Planting and Deep Processing of Authentic Medicinal Materials in the Three Gorges Reservoir Area, Chongqing 404120, China
To determine the chloroplast genomes ofand, analyze their sequence characteristics, and identify the relationship betweenspecies.In this study, the next-generation sequencing ofandwas performed on the Illumina Hisep 2 500 sequencing platform. Two complete chloroplast genome sequences were obtained after assemblying and annotationing,. Then, the two sequence structure and the phylogenetic relationship ofwere analysed by bioinformatics methods.The study found that the total length of the chloroplast genome ofwas 159 786 bp and the total GC content was 37.2%. The whole genes annotated were 131, including 88 protein-coding genes, 37 tRNA genes and six rRNA genes. Similarly, the total length of the chloroplast genome ofwas 159 652 bp and the total GC content was 37.1%. A total of 131 genes were annotated, which including 88 protein-coding genes, 37 tRNA genes and six rRNA genes in all. There were 112 and 227 simple repeat sequences were detected fromandrespectively, and they both had the largest number of codons encoding leucine and the least number of codons encoding tryptophan. The phylogenetic tree showed that,,,andwere clustered together into one family.was very similar toand, and the support rate reached 100%.The chloroplast genome structure and phylogenetic relationship of two species ofwere analysed. The results of this study will provide a scientific basis for the accurate identification, development and utilization ofmedicinal plants and resource protection.
Lindl.;Rchb. f.; medicinal plants; complete chloroplast genome; genetic relationship
R286.12
A
0253 - 2670(2022)16 - 5159 - 11
10.7501/j.issn.0253-2670.2022.16.025
2022-02-15
重庆市自然科学基金资助项目(cstc2018jcyjAX0267)
李卓蔚(1997—),硕士,主要从事环境微生物研究。Tel: 15808197965 E-mail: lizhuowei0@126.com
吴应梅(1981—),实验师,博士,主要从事食品药品功能研究。Tel: (023)58102522 E-mail: wuyingmei0927@126.com
周 浓(1978—),教授,硕士生导师,主要从事中药炮制与资源研究。Tel: (023)58576130 E-mail: erhaizn@126.com
[责任编辑 时圣明]
猜你喜欢
医学信息(2019年4期)2019-10-08
课程教育研究·学法教法研究(2019年18期)2019-10-08
少儿科技(2019年9期)2019-09-10
烹调知识(2019年3期)2019-03-01
当代党员(2018年21期)2018-11-29
党员生活(2018年14期)2018-08-11
环球时报(2016-08-23)2016-08-23
江苏农业科学(2016年2期)2016-04-11
轻兵器(2016年5期)2016-03-15
分析化学(2014年4期)2014-10-24
