
干扰素通路调控与自身免疫疾病*
2022-08-20 06:12李伟杨晗穆昕*
生物化学与生物物理进展 2022年8期
李 伟 杨 晗 穆 昕*
(1)泰州医药高新技术产业开发区(泰州市高港区)医药产业园管理办公室,泰州 225300;2)天津大学药物科学与技术学院,天津 300072;3)天津大学-汉氏联合药物创新与转化联合实验室,天津 300072)
天然免疫是机体抵抗病原微生物侵染的第一道屏障,主要由皮肤、黏膜等组织所组成的物理屏障以及天然免疫细胞等组成的化学防御组成[1-2]。如皮肤受损等情况的发生所导致的病原微生物入侵机体,会引发被感染细胞、组织包括干扰素信号(interferon signaling,IFN signaling)通路和炎症信号(如NF-κB signaling)通路等应激信号通路的激活。这些通路既能直接参与抗病原微生物侵染的过程,又参与调动适应性免疫系统对受侵染部位做出防御反应[3-4]。其中,由干扰素信号通路激活而引发的大量防御基因的表达上调,是细胞对抗病原微生物侵染的有力手段,在天然免疫中发挥关键作用。
干扰素是由来自英国的病毒学家Alick Isaacs和瑞士的微生物学家Jean Lindenmann共同发现的[5]。流感病毒感染鸡胚绒毛尿囊膜后,细胞被诱导产生一种“干扰”因子抑制流感病毒感染其他未感染的细胞,这种分泌蛋白被他们命名为“干扰素”[5]。随着对干扰素的研究不断深入,人们对该通路的认识不断加深,对其激活机制、发挥抗病原微生物作用的功能原理和网络调控等方面有了越来越多的认识。很多自身免疫疾病的发生与干扰素信号通路的异常激活密切相关,而调控干扰素信号通路的关键基因发生功能缺失型突变或功能获得型突变,是导致通路异常激活的重要原因[6-8]。本综述将从干扰素通路、自身免疫疾病特点和二者之间的关联这三方面展开讨论。
1 干扰素信号通路
细胞的DNA通常存在于细胞核和线粒体中,其所转录的RNA多为单链形式的mRNA或含有少量二级结构的rRNA、tRNA等[9]。这些核酸分子在各自的细胞器中发挥着各自应有的作用。病原微生物侵染细胞,会打破细胞内不同类型分子分布的平衡:DNA病毒,如单纯疱疹病毒1(herpes simplex virus type 1,HSV-1),在侵染、复制过程中,使病毒DNA在细胞质中积累[10];脑心肌炎病毒(encephalomyocarditis virus,EMCV)、水疱性口炎病毒(vesicular stomatitis virus,VSV)、甲型流感病毒(influenza A virus,IAV)等RNA病毒在基因组RNA复制的过程中,其所形成的双链RNA(double-stranded RNA,dsRNA)中间体,会出现在细胞质或内吞体中[11-14]。此外,线粒体DNA会在如单纯疱疹病毒或登革热病毒感染等情况下,从应激受损的线粒体中释放至细胞质中[12,15]。这种来自于病原微生物或与之直接相关的、具有特定结构特征的分子被统称为病原相关模式分子(pathogen-associated molecular patterns,PAMPs)。模式分子不仅局限于核酸类,还有细菌的脂多糖(lipopolysaccharide,LPS)、鞭毛蛋白等[16]。满足以下两点之一,就可以称之为PAMPs:a.来自于病原微生物的特定类型分子,如dsRNA、LPS等;b.分子类型为细胞已有,但是出现在错误的地点,如细胞质中出现的DNA、内吞体中出现的核酸分子等[17-18]。
细胞进化出受体,特异性结合这些模式分子,并触发下游的干扰素通路,这一类受体分子统称为模式识别受体(pattern recognition receptors,PRRs)[17],如细胞质中识别DNA的cGAS(cyclic GMP-AMP synthase)、识别dsRNA的RIG-I(retinoic acid-inducible gene I) 和 MDA5(melanoma differentiation-associated protein 5),内吞体中识别dsRNA的TLR3(Toll-like receptor 3)、识别单链RNA(single-stranded RNA,ssRNA)的TLR7或TLR8、识别DNA的TLR9,细胞膜上识别LPS的TLR4和细菌鞭毛的TLR5等[17]。不同的受体-模式分子结合,会激活下游不同的适配蛋白(adaptor),之后会采用相同或类似的策略招募蛋白激酶(如TBK1等)完成对干扰素调节因子(interferon regulatory factors,IRFs)IRF3或IRF7的磷酸化。磷酸化的IRF3或IRF7发生构象变化,形成同源二聚体,进入细胞核行使转录因子的功能,激活I型干扰素(如IFNα或IFNβ等)mRNA的转录[19-20]。I型干扰素表达后分泌至细胞外,通过结合自己和周围细胞的膜上受体(interferon-α/β receptor,IFNAR),引发其形成二聚体并激活下游JAK-STAT信号通路,最终激活细胞的效应基因表达,实现直接的抗病原微生物功能和对细胞生长、代谢状态的调节,以更好地适应病原微生物入侵的应激环境[21](图1)。
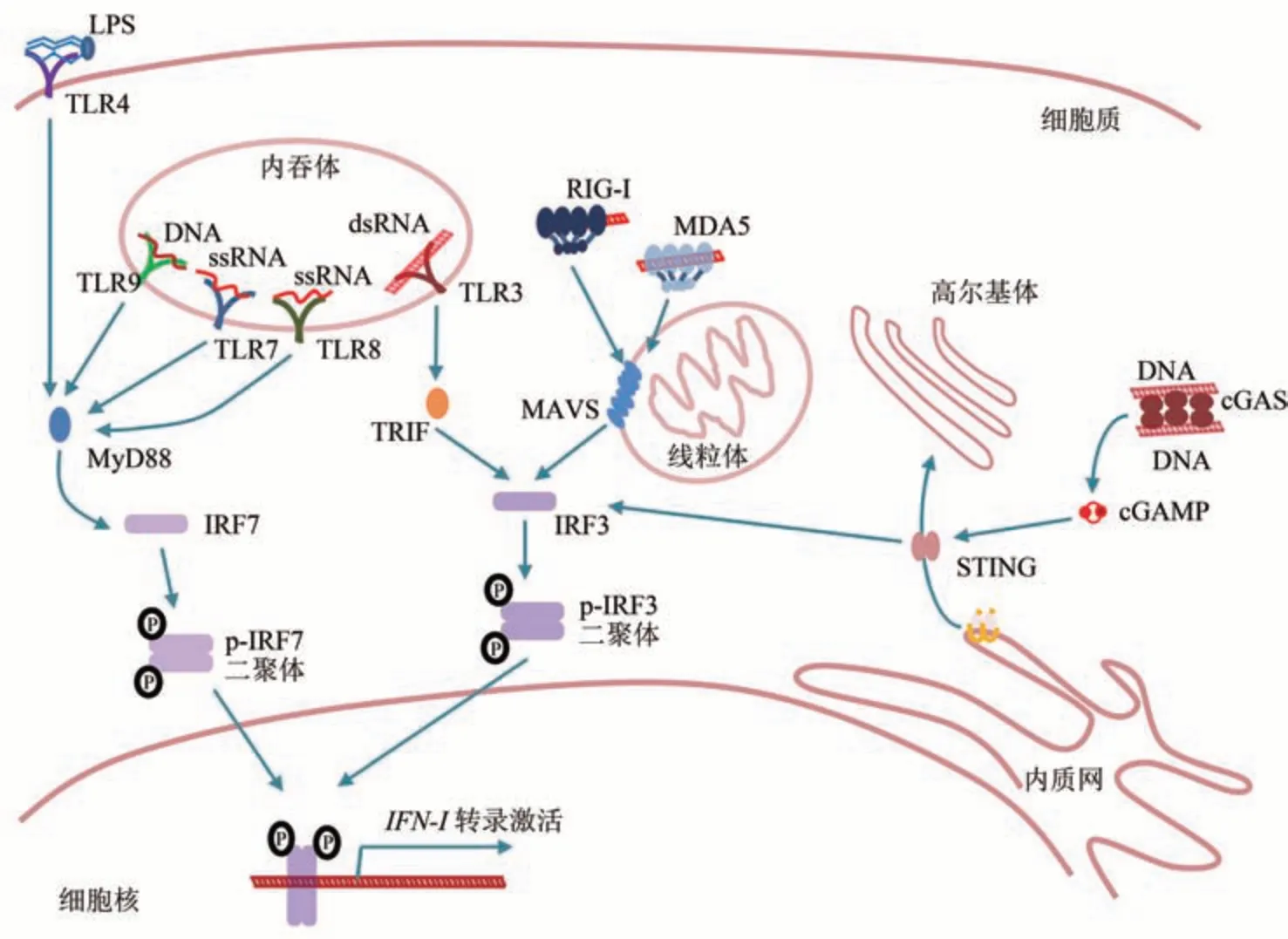
Fig.1 Schematic view of IFN signaling activation[22-23]图1 干扰素信号通路工作示意图[22-23]
2 自身免疫疾病的发生
模式识别受体对“自己”和“非己”分子的正确识别,是干扰素通路发挥功能的关键。基因突变所引发的蛋白质功能改变是导致干扰素通路异常激活的重要原因[7,24]。由单基因突变所引发的干扰素信号通路异常激活并最终引发的自身免疫疾病,在遗传学上符合孟德尔定律,被称为I型干扰素疾病(type I interferonopathy),这一概念由Yanick J.Crow最先提出[25-26]。这类疾病包括Aicardi-Goutières综 合 征(Aicardi-Goutières syndrome,AGS)、 脊 椎 软 骨发 育 不 良(spondyloenchondrodysplasia,SPENCD)、单基因突变引发的红斑狼疮(monogenic lupus)和补体缺乏(complement deficiency)等[25-26]。因为是单基因突变引发,所以这些突变体在研究基因功能上具有重要的提示作用,对理解基因产物对干扰素信号通路调控中如何发挥作用具有重要的指导意义。下面以AGS为例介绍该类疾病的病理特征,探讨致病机理。
AGS是一种罕见的、早发型I型干扰素疾病,以非感染状态下免疫系统的异常激活和I型干扰素的不正常上调为特征,主要影响脑、皮肤和免疫系统[27]。相比较于系统性红斑狼疮这种受到来自于遗传、环境和生活习惯等多种因素影响而引发的疾病,AGS发病较早,1岁新生儿就会出现严重的脑部功能障碍,特定单基因的突变就可致病是其早发性的重要原因。脑部的障碍进一步恶化新生儿的饮食摄入和非感染状态下发烧症状的发生,最终导致智力严重受损、头部尺寸变小等。患者皮肤出现疼痛、瘙痒和红肿的表型,多发病于肢体末端,如手指、脚趾和耳部等。这些症状主要由血管炎症引起,寒冷会加剧病情恶化。多数患者无法存活过童年期,但也有少数病情较轻的患者能够生存至成年[28]。
高通量测序和生物信息学分析发现,单基因突变是AGS的致病原因,目前已经发现的突变基因有9种,这些基因被赋予一个AGS编号,以便于对其进行归纳研究:TREX1(AGS1)、RNASEH2A(AGS4)、RNASEH2B(AGS2)、RNASEH2C(AGS3)、SAMHD1(AGS5)、ADAR1(AGS6)、IFIH1(编码MDA5蛋白)(AGS7)、LSM11(AGS8)和RNU7-1(AGS9)[29-34]。基因突变分为功能获得型(gain-of-function,GOF)和功能丧失型(loss-of-function,LOF)两种。除IFIH1基因为功能获得性突变致病之外,其余基因都发生了功能丧失型突变。有趣的是,在这些基因的编码产物中,MDA5直接参与干扰素通路的激活,其余均参与细胞内核酸(DNA、RNA)的代谢调控。这些突变体的发现,将细胞内环境稳态与调控和干扰素通路紧密联系到了一起,提示干扰素通路本身可能是作为一种监控细胞状态的机制,病原微生物侵染只是代表了一种应激状态。
3 细胞内环境调控与干扰素通路和自身免疫疾病
细胞内的核酸包括DNA和RNA,前者主要分布于细胞核和线粒体,后者广泛分布于细胞核、线粒体和细胞质等。DNA通过自身复制,保证了细胞分裂中遗传物质的稳定传代,其转录的RNA,形成mRNA、tRNA、rRNA和多种ncRNA,共同参与细胞的正常生理功能。
细胞核中的染色质DNA在复制的过程中,偶尔会错误地掺入NTP,产生短的RNA序列,这种RNA∶DNA杂合链会引起染色质损伤,造成DNA的断裂,引发cGAS对破损DNA分子的识别,进而激活干扰素信号通路[35-36];RNASEH2A/B/C复合物通过降解这些RNA序列,维持了细胞核中DNA的稳定性,避免了干扰素信号通路的激活[37-38]。这3个基因发生功能丧失型突变,会导致细胞内干扰素信号通路的异常激活[39]。
基因组DNA中存在大量的转座子序列[40]。有证据表明,SAMHD1(SAM domain and HD domain-containing protein 1)不仅在稳定细胞内dNTP水平中发挥重要作用,在抑制转座子活性中也至关重要[41-43]。该蛋白质抑制内源性转座子LINE-1的逆转座活性,降低细胞质中DNA水平。转座子活性的提高将导致细胞质中DNA序列的积累,激活cGAS受体。SAMHD1基因发生功能丧失型突变或表达沉默,将导致细胞中干扰素信号通路的异常激活[44]。
上述机制保证了细胞核中DNA的稳定和细胞质中DNA的缺失。一旦出现残留于细胞质中的DNA分子,将由TREX1(three prime repair exonuclease 1)负责清除[45]。在TREX1敲除的细胞中,发现大量转座子DNA积累于细胞质中,成为cGAS的激活配体[46-47]。此外,从损伤的线粒体中泄漏的DNA,也会在细胞质中被TREX1降解,避免被cGAS识别[48]。因此,该基因的功能丧失型突变或表达沉默,将导致干扰素信号通路的异常激活[49]。
最近的研究发现,结合于基因组DNA的组蛋白功能受损也会引起cGAS介导的干扰素上调[34]。在对AGS病人的基因突变筛查中,Crow课题组[34]发现了两种新的基因突变,分别位于LSM11和RNU7-1基因中。这两种基因的编码产物对组蛋白mRNA前体的正确加工至关重要,其功能突变导致组蛋白表达水平的降低,使得基因组DNA暴露于外,被cGAS识别,引发干扰素信号通路的激活。
前面介绍的都是在DNA代谢中,关键基因发生功能丧失型突变会引发干扰素的异常激活。在RNA代谢中,细胞核中转录的RNA会因序列互补配对而含有二级结构,特别是人源细胞中一类称为Alu的转座子元件[50]。这是一种长约300个核苷酸的散在重复序列,在整合进基因组的过程中两个元件头对头或尾对尾整合在相邻的位置,那么其在所插入基因的转录过程中,就会形成一条约300 bp的双链结构[51]。这种双链结构被称为反向重复Alu,简称为IR-Alu(inverted repeat Alu)元件。细胞中编码一种RNA特异性腺苷脱氨酶(RNA-specific adenosine deaminase 1,ADAR1),能结合dsRNA并具有脱氨酶活性,将dsRNA中的腺嘌呤A催化为次黄嘌呤I。通常情况下,由ADAR1将IR-Alu中的部分A修饰为I,从而将A:U配对改变成I和U的组合,无法形成配对结构[51]。ADAR1通过这样的修饰作用,破坏了IR-Alu的双链结构,使得该RNA在细胞质中,不会被MDA5识别。但在发生功能丧失型ADAR1突变或功能获得型突变IFIH1(编码MDA5蛋白)的情况下,IR-Alu元件被错误识别为“非己”分子,从而通过MDA5-MAVS(mitochondrial antiviral signaling protein)途径,使细胞产生干扰素信号通路的异常激活[52-53]。
由此可见,干扰素通路的功能发挥,需要以细胞内环境的稳定作为前提的,一旦这一平衡被打破,干扰素通路将发生异常激活,从而引发机体炎症和自身免疫疾病的发生(图2)。
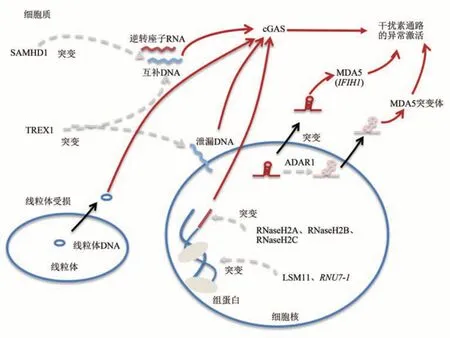
Fig.2 Aberrant IFN signaling due to gene mutation[27,34]图2 基因突变导致干扰素通路的异常激活[27,34]
4 总结与展望
综上所述,干扰素抗病原微生物信号通路作为细胞的一种应激反应,其正常活性需要精确调控,即使在非感染情况下,细胞新陈代谢、分裂复制的过程中,也会产生很多刺激干扰素信号的化学分子。通过调控因子的作用,保证了这些危险信号分子的降解。
随着对自身免疫疾病致病机理的研究越来越深入,这些调控因子对干扰素信号通路网络调控的作用越来越清晰,核酸代谢稳态对细胞在干扰素通路在非感染状态下的静息沉默至关重要。在未来的治疗策略研究中,这些调控因子也许会成为新的精准化检测目标和药物靶点,而不仅限于依赖于抑制已经被异常激活的干扰素信号通路[54]。近年来,针对DNA代谢失调所引发的AGS病例,已经有使用逆转录酶抑制剂进行治疗的临床探索[27,55]。在未来的数年间,预期将有越来越多的特异性抑制剂或生物治疗手段用来进行相关测试、研究[56]。
猜你喜欢
中国现代医生(2022年19期)2022-11-04
按摩与康复医学(2022年19期)2022-09-27
中国现代医生(2022年19期)2022-08-25
保健与生活(2021年8期)2021-04-19
农民致富之友(2020年18期)2020-06-19
中学生理科应试(2017年6期)2017-09-27
中学生理科应试(2017年2期)2017-04-01
中国动物保健(2015年6期)2015-10-21
销售与管理(2006年9期)2006-09-17
中国青年(1984年7期)1984-08-20
