
乌苏里拟鲿igf基因克隆及其与生长性状关联分析
2022-09-21 09:02魏梦雅王思梦朱传坤
水产科学 2022年5期
魏梦雅,王思梦,宁 静,范 帆,朱传坤
( 淮阴师范学院 生命科学学院,江苏省环洪泽湖生态农业生物技术重点实验室,江苏省特色水产繁育工程实验室,江苏省区域现代农业与环境保护协同创新中心,江苏 淮安 223300 )

乌苏里拟鲿(Pseudobagrusussuriensis)是鲇形目鲿科的一种底栖中型鱼类,自然分布于黑龙江至珠江流域的水系中[7]。乌苏里拟鲿肉质细嫩、味道鲜美,粗脂肪含量高,较一般淡水鱼适口性好,深受消费者喜爱,经济价值极高[8]。受环境污染、人类活动干扰及生境破坏等因素的影响,乌苏里拟鲿资源量锐减,野生资源无法满足市场需求,目前已在全国十余省份开展人工养殖[9]。与大多数养殖鱼类面临的问题类似,乌苏里拟鲿养殖过程中也不可避免地出现了品质退化现象,亟需开展其种质改良及优良品种选育工作。然而,由于对乌苏里拟鲿的研究起步较晚,且主要集中于人工繁殖[10-12]、养殖模式改进[13-15]、性别决定[16-17]、遗传分析[18-20]等方面。利用候选基因法筛选与特定性状关联的分子标记,对定向改良水产动物性状具有重要作用,并已在多种鱼类中筛选到与生长性状关联的优势基因型[21]。在乌苏里拟鲿中,生长相关基因数据极少,对于下丘脑—垂体—肝生长调控轴中的相关基因,目前尚未见相关研究报道。因此,笔者通过转录组与二代基因组测序数据结合的基因克隆技术,扩增获得乌苏里拟鲿生长调控轴中igf1基因和igf2基因的DNA和cDNA序列,并分析基因序列中的多态位点与体质量、体长等主要生长性状间的关联性,以期为乌苏里拟鲿生长发育调控机制研究、基因进化研究以及采用候选基因法开展分子标记辅助育种等工作提供参考资料。
1 材料与方法
1.1 试验材料、数据采集及基因组DNA提取
乌苏里拟鲿样本均来自淮安市水产技术指导站,随机挑选80尾当年繁殖、养殖至3月龄的同批次幼鱼作为试验鱼。测量每尾个体的体质量(m)、体宽、体长(L)、头长,并计算其肥满度指数(K)。
K=100m/L3
每尾个体剪取少量尾鳍组织,置于无水乙醇中,并于4 ℃冰箱中保存备用;另取部分尾鳍组织,采用通用的苯酚—氯仿法抽提基因组DNA[22]。
1.2 引物设计与igf基因片段克隆
乌苏里拟鲿igf基因片段克隆所用引物依据转录组和基因组二代测序(数据未发表)所得部分片段及其近缘物种黄颡鱼(Pelteobagrusfulvidraco)(KX434878.1、HM210742.1)和斑点叉尾(AH015082.2、NM_001200295.1、AY615885.1)中igf1和igf2基因序列,利用Primer premier 5.0软件设计(表1)。

表1 乌苏里拟鲿igf1和igf2基因克隆和多态分析引物Tab.1 Primers used for cloning and polymorphic analysis of igf1 and igf2 genes in Ussuri catfish P. ussuriensis
PCR反应体系总体积为50 μL:10×PCR buffer 5 μL、灭菌超纯水37.4 μL、引物(2.5 μmol/L)1.6 μL、dNTPs(2.5 mmol/L)1.6 μL、模板DNA 4 μL(约500 ng)、Taq DNA聚合酶0.4 μL(北京康为世纪公司)。PCR反应在96孔PCR仪中运行,反应程序如下:94 ℃预变性5 min;94 ℃变性40 s,适宜温度条件下退火40 s,72 ℃延伸45 s,共36个循环;72 ℃终延伸10 min。PCR产物利用1%琼脂糖凝胶电泳进行分型,并采用回收试剂盒进行切胶回收纯化。纯化产物于16 ℃条件下过夜连接12 h,连接到pMD18-T载体中[宝生物工程(大连)有限公司],并转化到感受态DH5α大肠杆菌(Escherichiacoli)中。阳性克隆通过PCR检测后,利用武汉艾康健生物科技有限公司的ABI DNA3730测序仪进行测序。
1.3 igf基因序列分析
通过转录组数据中所得igf基因的cDNA序列与克隆所得DNA序列比对分析,参照黄颡鱼(Pelteobagrusfulvidraco)和斑点叉尾的igf基因序列,确定乌苏里拟鲿igf基因的外显子和内含子边界(GT-AG)。igf基因的氨基酸序列通过在线工具包Sequence Manipulation Suite(https://www.biosoft.net/sms/)进行预测,并同时进行氨基酸序列的一致性和相似性分析。分别利用SignalP v.4.0 server(https://www.cbs.dtu.dk/services/SignalP/)和ProP 1.0 web server (https://www.cbs.dtu.dk/servi ces/ProP/)进行信号肽和成熟肽的预测。
为分析乌苏里拟鲿igf基因与其他物种的进化关系,将乌苏里拟鲿igf基因编码序列与其他鱼类igf基因编码序列进行聚类分析,构建系统进化树。用于比较的近源物种共包括鲇形目2种、鲤形目3种、鲈形目2种、鲑形目2种、鲉形目1种、外源物种1个(表2)。利用ClustalX软件进行聚类分析,利用MEGA 7.0软件的邻接法对聚类分析后的igf基因序列构建系统进化树,自展次数设置为1000次,以斑马鱼igf3基因为外类群。

表2 系统进化分析物种的igf基因序列信息Tab.2 Information on igf genes of species used for phylogenetic analysis
1.4 多态位点筛选及其与生长性状的关联分析
利用克隆乌苏里拟鲿igf基因的引物对所得igf基因序列进行分段分析,查找多态位点。将各引物在10尾个体中扩增后,进行PCR产物测序。PCR体系组成和反应程序与1.2中所述一致。测序所得序列通过ClustalX软件比对,结合对测序峰图的观察,来筛选igf基因中的单核苷酸多态性(SNP)位点。
将包含SNP位点的引物在80尾个体中进行PCR扩增及PCR产物测序,通过测序峰图对每个个体进行基因分型。分型完成后,利用软件Popgene 32和Arlequin 3.11进行期望杂合度、观测杂合度、多态信息含量、哈迪—温伯格偏离指数、连锁不平衡等多态性分析。
利用SPSS 18.0软件中的一般线性模型进行多态位点不同基因型与体质量、体宽、体长、头长和肥满度等生长性状间的关联分析。由于试验鱼为同龄且养殖于相同环境,因此关联分析时不考虑世代、环境等因素影响,所用模型为:
Y=μ+G+e
式中,Y为体质量、体宽、体长、头长和肥满度生长性状的观测值,μ为各生长性状的平均值,G为各基因型的固定效应,e为随机残差。
利用邓肯多重比较来评估差异显著性,当P<0.05时为差异显著,当P<0.01时为差异极显著。
2 结 果
2.1 乌苏里拟鲿igf基因结构分析
克隆获得了乌苏里拟鲿igf1和igf2的基因序列。其中所得igf1基因cDNA长度为1241 bp,包括5′非编码区(UTR)290 bp,开放阅读框(ORF)477 bp,3′非编码区474 bp。由于内含子序列过长,未获得igf1基因DNA完整序列,获得的igf1基因部分DNA序列长度为10 922 bp;乌苏里拟鲿igf1基因共包含5个外显子和4个内含子;外显子1~5的序列长度分别为33、166、182、36、60 bp;内含子1所得部分长度为1101 bp,内含子2所得部分长度为6222 bp,内含子3长度为995 bp,内含子4部分长度为2127 bp(图1a)。所得igf2基因cDNA长度为2008 bp,包括5′非编码区 26 bp,开放阅读框741 bp,3′非编码区1241 bp。所得igf2基因DNA序列分析结果显示:该基因由4个外显子和3个内含子组成;外显子1长度为171 bp,外显子2为151 bp,外显子3为182 bp,外显子4为237 bp;内含子1部分长度为789 bp,内含子2长度为1649 bp,内含子3长度为521 bp(图1b)。乌苏里拟鲿igf1和igf2基因内含子和外显子间均具有显著的GT-AG边界特征。
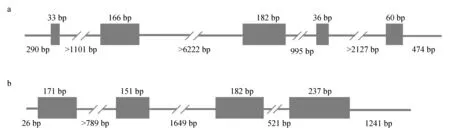
图1 乌苏里拟鲿igf1(a)和igf2(b)基因结构示意Fig.1 Diagram of the gene structures for igf1 (a) and igf2 (b) in Ussuri catfish P. ussuriensis
乌苏里拟鲿igf1基因编码多肽链长度为158个氨基酸,包括含41个氨基酸的信号肽序列,含70个氨基酸的B-C-A-D结构域以及含47个氨基酸的E结构域,此外,在第47、59、88、89、93和102位为6个保守的参与形成二硫键的半胱氨酸(图2)。与其他物种IGF1氨基酸序列相似性分析结果显示,乌苏里拟鲿与鲇形目鲿科鱼类黄颡鱼IGF1一致性(96.9%)和相似性(97.5%)均最高,其次为鲇形目科的斑点叉尾(93.7%和95.6%),与鲤形目3种鱼类的一致性相同,均为76.1%,与其他目鱼类的一致性相对较低(表3)。乌苏里拟鲿igf2基因编码246个氨基酸,其中信号肽长度为47个氨基酸,B-C-A-D结构域长度为70个氨基酸,E结构域由97个氨基酸组成,序列中也包含6个保守的半胱氨酸,分别位于第56、68、96、97、101和110位(图3)。与IGF1类似,乌苏里拟鲿IGF2与黄颡鱼的一致性(96.7%)和相似性(97.2%)最高,其次为斑点叉尾(88.8%和90.2%),所不同的是IGF2与鲑形目、鲈形目等鱼类间的一致性和相似性高于IGF1(表3)。

图2 乌苏里拟鲿与其他物种IGF1氨基酸序列比对结果Fig.2 Alignment results between Ussuri catfish P. ussuriensis and other species based on amino acid sequences of IGF1 peptides单线标注的为信号肽,双线标注的为B-C-A-D结构域,虚线标注的为E结构域,箭头所示的为6个保守的半胱氨酸;下同.The signal peptide is marked with a single line,the B-C-A-D domain is marked with a double line,the E domain is marked with a dotted line,and the six conserved cysteines are marked with arrows;et sequentia.

图3 乌苏里拟鲿与其他物种IGF2氨基酸序列比对结果Fig.3 Alignment results between Ussuri catfish P. ussuriensis and other species based on amino acid sequences of IGF2 peptides

表3 乌苏里拟鲿igf1和igf2基因编码氨基酸序列与部分物种的一致性和相似性分析结果Tab.3 Results of identity and similarity analyses between Ussuri catfish P. ussuriensis and other species based on amino acid sequences coding by igf1 and igf2 genes
2.2 乌苏里拟鲿与其他物种igf基因系统进化分析
将乌苏里拟鲿igf1和igf2基因编码区与鲇形目的黄颡鱼、斑点叉尾,鲤形目的斑马鱼、草鱼、团头鲂,鲈形目的尼罗罗非鱼、金头鲷,鲑形目的北极红点鲑、大西洋鲑,鲉形目的若鲉杜父鱼及外类群牛,进行系统进化分析。邻接系统发生树显示,igf1基因和igf2基因明显分为两大分支,分别包含上述物种。在每一分支中,乌苏里拟鲿首先与其所在鲇形目的鱼类黄颡鱼和斑点叉尾聚为一支,然后与鲤形目鱼类聚为一支,而鲈形目、鲑形目和鲉形目则聚为另一支,因此,这也反映出乌苏里拟鲿所在鲇形目与鲤形目的亲缘关系较近(图4)。

图4 基于乌苏里拟鲿与其他鱼类igf基因编码区的邻接系统发生树Fig.4 The Neighbor-Joining phylogenetic tree between Ussuri catfish P. ussuriensis and other species based on coding regions of igf genes
2.3 乌苏里拟鲿igf1和igf2基因多态位点筛选
对乌苏里拟鲿igf1和igf2基因的多态性进行了分段分析,结果显示,在igf1基因中共检测到5个SNP位点,其中1个位于外显子3上(g.8703A>G),其余4个集中于内含子4中(g.10021T>C、g.10034G>T、g.10210C>T和g.10211C>A)(图5)。此外,在igf1基因内含子中共发现7个微卫星序列,其中:内含子1中包含3个,分别为(GAAA)8、(TC)5和(TC)11CCTTC(CT)7;内含子2中包含3个,分别为(TCT)11、(TATT)8和(TG)5;内含子4中包含1个(CT)15微卫星序列。在乌苏里拟鲿igf2基因中共发现SNP位点5个,其中:1个位于外显子3中(g.2975C>T),2个位于内含子1中(g.228A>C和g.296A>C),2个位于内含子2中(g.1371A>G和g.1431C>T)(图6)。在igf2基因中共检测出5个微卫星位点,其中:内含子1中包含3个,分别为(TG)15N(TG)4(AG)5CGAGT(GA)4GTA(TG)13、(GA)5TAA(AG)5和(AG)6;内含子2中包含2个,分别为(TA)6CAC(GT)5和(TC)7。

图6 乌苏里拟鲿igf2基因中的SNP位点Fig.6 Positions of SNPs in the igf2 gene of Ussuri catfish P. ussuriensis
2.4 乌苏里拟鲿igf1和igf2基因中SNP与生长性状关联性分析
选取igf1基因外显子3中的SNP位点g.8703A>G和igf2基因外显子3中的SNP位点g.2975C>T开展基因型与体质量、体长、体宽、头长和肥满度5个生长性状间的关联性分析。多态性分析结果显示:g.8703A>G含2个等位基因,观测杂合度、期望杂合度和多态信息含量分别为0.374、0.502和0.450,符合哈迪—温伯格平衡(P>0.05)(表4);g.2975C>T也含2个等位基因,观测杂合度、期望杂合度和多态信息含量分别为0.374、0.501和0.413,也未偏离哈迪—温伯格平衡(P>0.05)(表4),两位点间不存在连锁不平衡现象(P>0.05)。在位点g.8703A>G处,A和G等位基因频率分别为47.50%和52.50%,共检测到3种基因型,其中:杂合基因型AG的基因型频率最高,为45.00%;AA和GG的基因型频率分别为25.00%和 30.00%(表5)。在g.2975C>T位点处,碱基C和T的等位基因频率分别为46.88%和53.12%,CC、CT和TT 3种基因型频率分别为26.25%、41.25%和32.50%,仍然是杂合基因型的频率最高(表5)。关联分析结果显示,这两个多态位点的基因型与生长性状间均无显著关联性(P>0.05)(表5)。

表4 乌苏里拟鲿igf1和igf2基因中外显子SNP位点的多态性信息Tab.4 Polymorphic information of SNPs in exons of igf1 and igf2 genes in Ussuri catfish P. ussuriensis
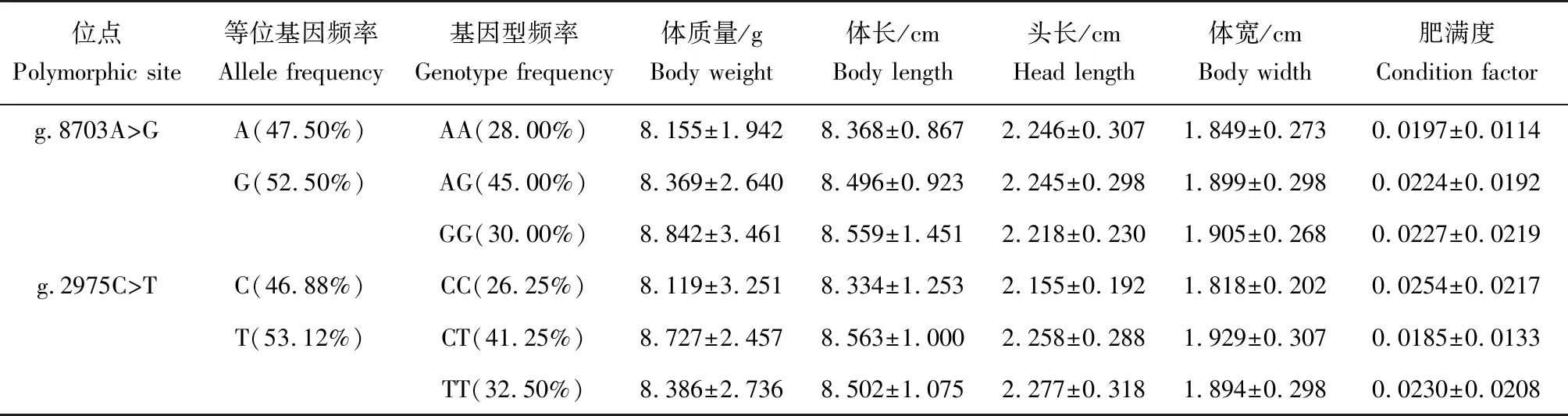
表5 乌苏里拟鲿igf1和igf2基因编码区SNP位点与生长性状关联分析结果Tab.5 Results of association analysis between growth traits and SNPs from coding region of igf1 and igf2 genes in Ussuri catfish P. ussuriensis
3 讨 论
3.1 igf1和igf2基因结构
在鱼类中,igf1和igf2基因的cDNA序列已被广泛报道,DNA则相对较少。目前的研究结果显示,鱼类igf1基因的序列长度差异较大,但均不超过20 kb,绝大多数鱼类的igf1基因均由5个外显子和4个内含子构成,并且内含子2的长度为4个内含子中最长[3,23-25]。本研究中,获得的乌苏里拟鲿igf1基因部分DNA长度10 922 bp,由5个外显子和4个内含子组成,与其他鱼类中报道结果类似,内含子2是4个内含子中最长的,长度大于6222 bp。在斑马鱼、金鱼(Carassiusauratus)等已报道鱼类中,igf2基因的DNA长度均小于igf1基因,外显子和内含子数目也不同于igf1基因,由4个外显子和3个内含子组成[26-27]。本研究中,所得乌苏里拟鲿igf2基因部分DNA序列长度为3700 bp,也由4个外显子和3个内含子组成。以上研究结果表明,乌苏里拟鲿igf1和igf2基因的结构与目前在其他鱼类中已报道的结构类似,这也反映出这两个基因在鱼类中具有较高的保守性。
3.2 igf1和igf2基因编码多肽的结构特征及进化分析
乌苏里拟鲿igf1和igf2基因的开放阅读框长度分别为477 bp和741 bp,编码氨基酸数目分别为158个和246个。乌苏里拟鲿IGF1的158个氨基酸序列由41个氨基酸组成的信号肽、70个氨基酸构成的成熟肽(B-C-A-D结构域)和47个氨基酸形成的E肽(E结构域)构成,其中包含6个保守的参与形成二硫键的半胱氨酸,这与在黄颡鱼[28]、金钱鱼[4]、白梭吻鲈(Sanderlucioperca)[29]等鱼类中报道的结构类似。乌苏里拟鲿IGF2肽链由246个氨基酸组成,成熟肽长度也为70个氨基酸,与IGF1成熟肽长度一致,序列中也包含6个保守的半胱氨酸,其结构与金钱鱼[4]、双棘黄姑鱼(Protonibeadiacanthus)[30]、白梭吻鲈[29]的IGF2结构类似。乌苏里拟鲿IGF1和IGF2氨基酸与鲇形目鲿科的黄颡鱼相应序列的一致性和相似性均最高,其次为鲇形目科的斑点叉尾和与鲤形目鱼类,这也与它们的分类地位相对应。此外,所不同的是乌苏里拟鲿的IGF2与鲑形目、鲈形目等鱼类IGF2间的一致性和相似性均高于IGF1,说明igf1基因进化过程中的变异程度高于igf2基因,因此,IGF2在鱼类中保守性较IGF1高。系统进化分析结果显示,igf1基因和igf2基因明显分为两支,乌苏里拟鲿的igf1基因和igf2基因分别首先与鲇形目鱼类聚为一支,然后和鲤形目鱼类聚为一支,最后和外类群牛聚为一支。该聚类结果表明,脊椎动物igf1基因和igf2基因间的分化时间要早于物种的分化,其他鱼类中的研究结果也均支持这一观点[4,31-32]。
3.3 igf1和igf2基因多态性及其与生长性状关联分析
对igf1和igf2基因多态性及其与经济性状的关联分析在畜禽中开展较早,并有大量研究报道。目前igf1和igf2基因的多态性及与目标性状的相关性分析在鱼类中也有较多报道,如:在鲤(Cyprinuscarpio)中,研究人员从igf1基因中检测到4个SNP位点,发现其中有2个与体质量和体长相关[33];阮瑞霞等[34]在吉富罗非鱼(Oreochromisniloticus)igf1基因中检测出3个SNP位点,发现2个与生长性状相关的SNP位点,并且其中一个与雄鱼质量增加、体宽/体长显著相关,另一个除与雄鱼体高/体长相关外,也与雌、雄鱼的质量增加显著相关;此外,在大西洋鲑、大口黑鲈(Micropterussalmoide)、白梭吻鲈等鱼类的igf1基因中也发现了较多与生长性状关联的多态位点[29,35-36]。鱼类igf2基因中多态性及关联分析也已有报道,如:在半滑舌鳎(Cynoglossussemilaevis)中,igf2基因编码区的SNP位点多态性与雄性生长性状、血液生化和基因表达显著相关[37];在罗非鱼中,研究人员在igf2基因内共鉴定出11个SNP位点,其中部分位点与罗非鱼体型和体质量等性状显著相关[38]。笔者在乌苏里拟鲿igf1基因中共检测到5个SNP位点和7个微卫星序列,其中1个SNP位点(g.8703A>G)位于外显子3上;在igf2基因中共发现5个SNP位点和5个微卫星位点,其中1个SNP位点(g.2975C>T)位于外显子3上。在igf1和igf2基因的各5个SNP位点中,均有3个是由碱基转换产生,另外2个为颠换;微卫星位点的比较发现,igf1基因中多碱基重复类型所占比例较高,而igf2基因中则均为二碱基重复类型。将igf1和igf2基因外显子中的SNP位点与生长性状进行了关联分析,其不同基因型个体的体质量、体长、体宽、头长、肥满度等生长性状均差异不显著,因此未检测到这两个位点与生长性状的关联性。类似结果在其他鱼类中也有广泛报道[33-34,39]。通过对编码氨基酸的分析,发现主要原因是乌苏里拟鲿igf1和igf2基因外显子上这两个SNP位点均为同义突变,碱基的改变并未引起氨基酸的变化,对表达产物也没有影响,无法影响生长性状。笔者所获得的igf1和igf2基因序列及多态性信息可为在乌苏里拟鲿中深入挖掘该基因功能、摸清其对生长发育的调控机制及分子标记辅助育种等研究提供基本数据。
4 结 论
乌苏里拟鲿igf1和igf2基因外显子和内含子数均与已报道硬骨鱼类相应基因中的数目一致,两基因编码氨基酸序列组成也与其他鱼类中报道的类似。乌苏里拟鲿igf1和igf2基因中均存在较多多态位点,并且均包含位于外显子中的SNP位点,但均为同义突变,与生长性状无显著关联性。
猜你喜欢
分子催化(2022年1期)2022-11-02
分子诊断与治疗杂志(2022年9期)2022-10-09
分析测试学报(2022年9期)2022-09-21
中国农业科学(2022年16期)2022-09-19
儿童时代·幸福宝宝(2020年9期)2020-09-08
电脑知识与技术(2018年19期)2018-11-01
小星星·阅读100分(高年级)(2017年3期)2017-03-22
湖北农业科学(2014年11期)2014-09-10
