
晚熟桃优良芽变新品种‘韦端蜜红’的分子标记鉴定
2022-10-08 10:46李建安陈小梅李成悦尚娟娥魁小花杨宏娟曾志芳马翠兰邱栋梁
福建农林大学学报(自然科学版) 2022年5期
李 文, 李建安, 陈小梅, 李成悦, 尚娟娥, 魁小花, 杨宏娟, 曾志芳, 马翠兰, 邱栋梁
(1.福建农林大学园艺学院,福建 福州 350002;2.古田县农业农村局经济作物站,福建 宁德 352200)
芽变是体细胞突变的一种,既是植物产生新变异的丰富源泉,也是果树选育新品种的一种简易而有效的方法.果树芽变是一种遗传物质变异,而DNA分子标记技术以蛋白质、核酸分子的突变作为基础,能够检测生物的遗传结构及其变异.目前,分子标记技术已被广泛应用于桃[1-2]、枣[3]、葡萄[4]、杧果[5]、杏[6]、李[7]、杨桃[8]等果树遗传多样性分析、品种鉴定、遗传图谱构建、基因定位及克隆中.简单重复序列间扩增(inter-simple sequence repeat, ISSR)是通过PCR技术,对相隔不远的SSR序列间的基因组DNA进行扩增的分子标记技术,因其引物与试验易设计、操作简单、成本低、效率高、多态性高、适用更广泛等优点深受科研工作者青睐[9];相关序列扩增多态性(sequence-related amplified polymorphism, SRAP)是一种基于PCR反应,通过独特的引物设计对可读框进行扩增的分子标记技术[10],具有稳定、简便、在基因组中均匀分布、克隆目标片段方便等优点.Huang et al[11]研究表明,随机扩增多态DNA(RAPD)、简单重复序列(SSR)、反转录转座子间扩增多态性(IRAP)和反转录转座子微卫星扩增多态性(REMAP)等4种分子标记方法不能区分‘砂糖橘’及其芽变株‘无籽砂糖橘’,仅有少数ISSR和SRAP引物获得了特异性条带,说明了这两种分子标记方法在鉴定芽株系中的可行性.分子标记在桃及其芽变株系的鉴定中发挥着重要作用.如:孙淑霞等[12]通过ISSR、SSR标记分析结合物候期观察及果实性状,比较了‘北京28号’桃及其芽变材料的亲缘关系;李海炎等[13]采用SRAP分子标记技术对‘川中岛’桃及其芽变株系的遗传关系进行了鉴定;文露等[1]通过ISSR和SRAP两种分子标记技术,对‘皮球桃’及其黄肉芽变变异株进行了鉴定.
桃在自然和栽培条件下具有较高的芽变频率,其品种(系)多,存在亲缘关系不详或同物异名的问题.‘韦端蜜红’桃是福建省宁德市古田县吉巷乡韦端村1994年定植的‘颐红’桃芽变优良单株的无性系后代,具有晚熟、果大、肉脆、味甜、植株抗流胶病等优良性状.本研究以‘韦端蜜红’和‘颐红’等6份桃材料为研究对象,利用ISSR和SRAP分子标记技术,分析其在遗传上的相似性与差异性,进一步探讨其亲缘关系,旨在为该种质的收集、利用及推广应用提供参考.
1 材料与方法
1.1 材料
供试的6份桃材料信息如表1所示,采集时间为2019年6月.选取新鲜嫩叶,放进自封袋中,置于冰上,带回实验室,经液氮速冻后置于冰箱(-80 ℃)中保存.供试的6份晚熟桃品种(系)分别采自福建省宁德市古田县凤埔乡峦垅村的福建省益康园农场有限公司(简称‘益康园’)、湖南省怀化市的芷江侗族自治县康瑞农生态农业有限公司(简称‘康瑞农’)、福建省宁德市古田县吉巷乡韦端村(简称‘韦端村’).C4是以芷江‘颐红’为接穗,毛桃为砧木嫁接而来.
1.2 方法
1.2.1 桃叶片DNA的提取 采用改良的 CTAB 法提取桃叶片DNA,参照孙月婷等[14]的方法并进行改进,提取的DNA置于冰箱(-20 ℃)保存备用.
1.2.2 ISSR-PCR扩增 20 μL体系总体积包括:10 μL 2×easy taq PCR Super Mix(+dye)(北京全式金生物技术有限公司)、0.3 μmol·L-1引物(福州尚亚生物技术有限公司)、50 ng·μL-1DNA模板,用ddH2O补充至20 μL.PCR扩增程序设置为:94 ℃预变性3 min,94 ℃变性30 s,52 ℃(退火温度因引物不同而有所不同)退火45 s,72 ℃延伸90 s,循环35次;而后于72 ℃延伸7 min;最后置4 ℃保存.在1%琼脂糖凝胶中用1×TAE缓冲液进行电泳分离,之后用JS-3000全自动凝胶成像分析仪(上海培清科技有限公司)进行拍照分析.
1.2.3 SRAP-PCR扩增 25 μL体系总体积包括:12.5 μL 2×easy taq PCR Super Mix(+dye)(北京全式金生物技术有限公司)、0.5 μmol·L-1引物(福州尚亚生物技术有限公司)、80 ng·μL-1DNA模板,加ddH2O补充至25 μL.PCR扩增程序设置为:94 ℃预变性5 min,94 ℃变性1 min,35 ℃退火1 min,72 ℃延伸1 min,循环5次;而后于94 ℃变性1 min,50 ℃退火1 min,72 ℃延伸1 min,循环35次;最后于72 ℃延伸10 min.PCR扩增结束后保持12 ℃,最后置4 ℃保存.在1.5%琼脂糖凝胶中用1×TAE缓冲液进行电泳分离,之后用JS-3000全自动凝胶成像分析仪(上海培清科技有限公司)进行拍照分析.
1.2.4 引物筛选 根据已经优化好的体系,选用高纯度DNA样品为模板,ISSR引物参照哥伦比亚大学(UBC)公布的第9套引物序列进行筛选,根据前人的研究结果[12,15-18],选择55 条引物进行筛选,用退火温度范围内较为合适的温度进行PCR扩增;SRAP引物参考前人研究的结果[10,19-20],选择12条正向引物与14条反向引物进行交叉筛选,以获得适合的引物.
1.3 数据处理
参考孙月婷等[14]的方法,建立二元矩阵,构建聚类树状图并分析遗传相似系数.
2 结果与分析
2.1 6份桃材料的ISSR分析
2.1.1 引物筛选与多态性分析 采用筛选出的17条多态性好、稳定性高、条带清晰的ISSR引物对6份桃材料进行PCR扩增,最终共扩增出91条DNA条带,其中,多态性条带18条,多态百分率为19.78%.ISSR引物扩增结果及其多态性如表2所示.17条引物平均扩增出5.35条条带,能有效区分‘韦端蜜红’与‘颐红’.引物UBC855的扩增结果见图1,UBC880的扩增结果见图2,可以发现‘韦端蜜红’出现了不同于‘颐红’的多态性条带.

图1 ISSR引物UBC855扩增6份桃材料的电泳图谱
2.1.2 聚类分析 用非加权组平均法(unweighted pair-group method with arithmetic means, UPGMA)进行聚类分析,得到了6份桃材料的聚类树状图(图3),其相似系数为0.88~0.94.当以相似系数0.92为阈值时分为两类,第一类包含‘韦端蜜红’(C1、C2、C3),第二类包含芷江‘颐红’(C4、C5)和古田‘颐红’(C6),此时可将‘韦端蜜红’与‘颐红’区分.
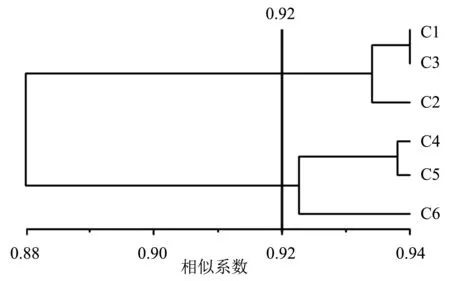
图3 基于ISSR标记的6份桃材料的UPGMA聚类树状图
2.1.3 遗传相似系数分析 采用Ntsys-pc 2.10e软件计算6份桃材料之间的遗传相似系数,结果(表3)显示,‘韦端蜜红’与‘颐红’间的遗传相似系数为0.844 4~0.943 2,平均遗传相似系数为0.900 6.‘韦端蜜红’(C1、C2、C3)间的遗传相似系数为0.931 0~0.943 2,芷江‘颐红’(C4、C5)与古田‘颐红’(C6)间的遗传相系数为0.918 6~0.941 2,遗传相似程度均较高.

表3 基于ISSR标记的6份桃材料之间的遗传相似系数
2.2 6份桃材料的SRAP分析
2.2.1 引物筛选与多态性分析 17对SRAP引物共扩增出85条DNA条带,其中,多态性条带21条,多态百分率为24.71%.SRAP引物扩增结果及其多态性如表4所示.17对引物平均扩增出5条条带,可将‘韦端蜜红’与‘颐红’区分.引物me1/em8的扩增结果见图4,me4/e11的扩增结果见图5,可以发现‘韦端蜜红’在不同引物中均出现了不同于‘颐红’的多态性条带.

图4 SRAP引物me1/em8扩增6份桃材料的电泳图谱
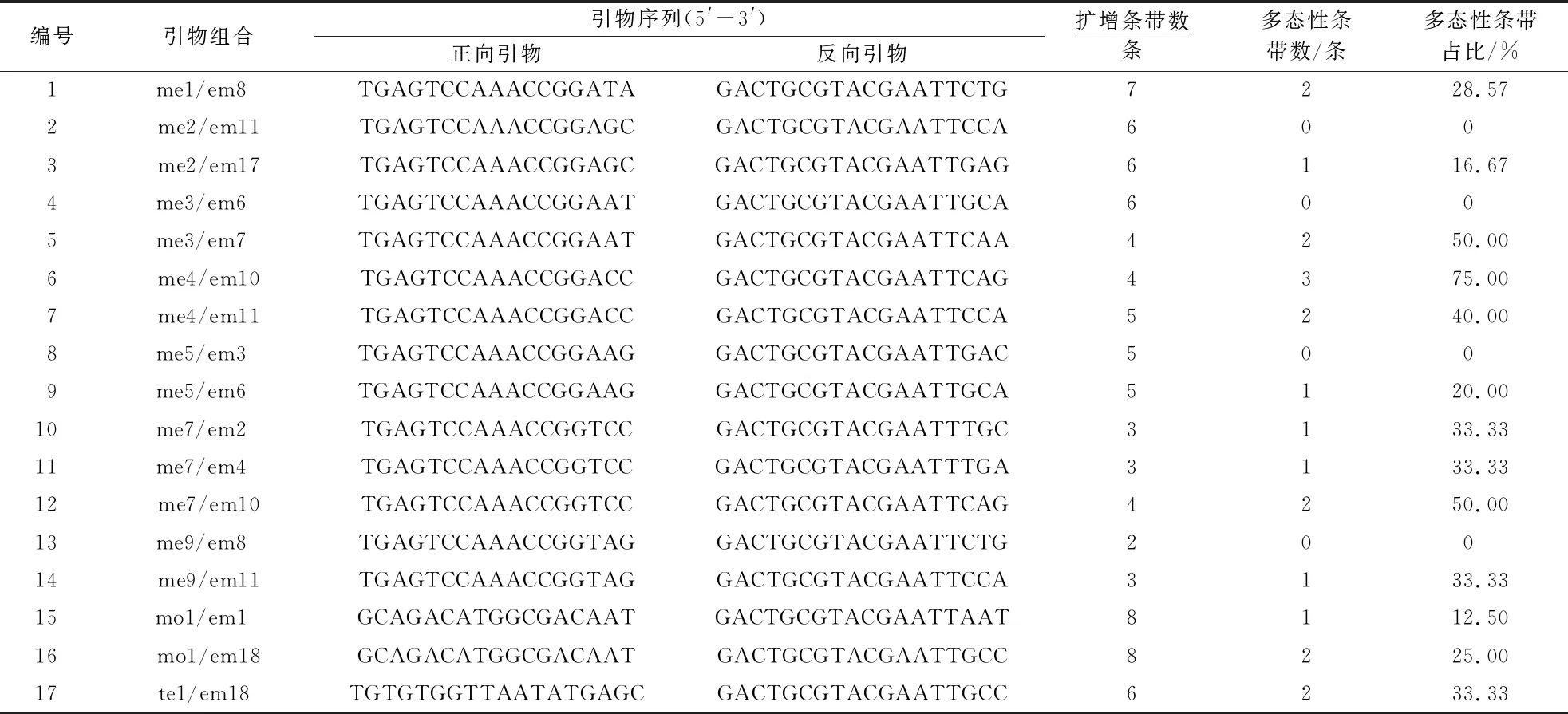
表4 SRAP引物及其产物多态性
2.2.2 聚类分析 使用UPGMA方法进行聚类分析,得到6份桃材料的聚类树状图(图6),其相似系数为0.85~0.94.当以相似系数0.89为阈值时分为两类,第一类包含‘韦端蜜红’(C1、C2、C3),第二类包含芷江‘颐红’(C4、C5)和古田‘颐红’(C6),此时可将‘韦端蜜红’与‘颐红’区分.

图6 基于SRAP标记的6份桃材料的UPGMA聚类树状图
2.2.3 遗传相似系数分析 采用Ntsys-pc 2.10e软件计算6份桃材料之间的遗传相似系数为0.807 2~0.935 9,平均遗传相似系数为0.870 0(表5).‘韦端蜜红’(C1、C2、C3)间的遗传相似系数为0.898 7~0.935 9,芷江‘颐红’(C4、C5)与古田‘颐红’(C6)间的遗传相系数为0.881 6~0.910 3,均具有较高的遗传相似系数.
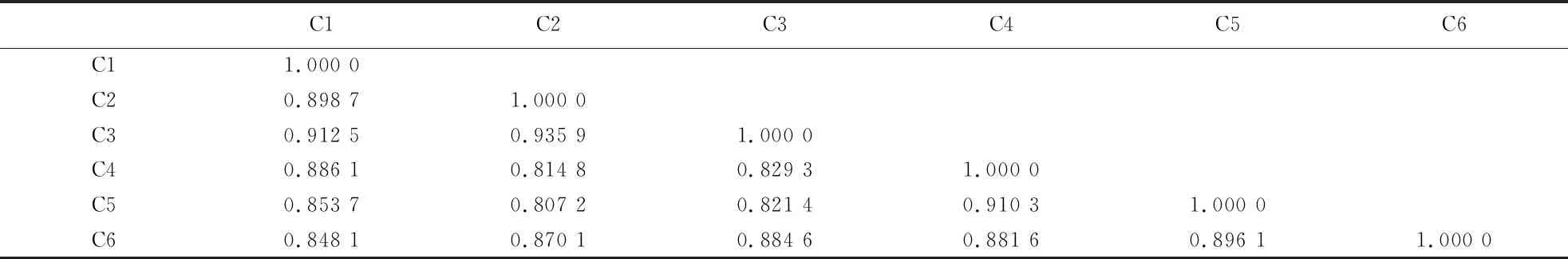
表5 基于SRAP标记的6份桃材料之间的遗传相似系数
2.3 6份桃材料的ISSR与SRAP联合分析
2.3.1 聚类分析 采用UPGMA方法对ISSR和SRAP数据进行聚类分析,得到6份桃材料的聚类树状图(图7),其相似系数为0.86~0.94.当以相似系数0.90为阈值时,可将6份桃材料分为两类,第一类包含‘韦端蜜红’(C1、C2、C3),第二类包含芷江‘颐红’(C4、C5)和古田‘颐红’(C6),此时可将‘韦端蜜红’与‘颐红’区分.

图7 基于ISSR和SRAP标记的6份桃材料的UPGMA聚类树状图
2.3.2 遗传相似系数分析 采用Ntsys-pc 2.10e软件计算6份桃材料之间的遗传相似系数为0.826 6~0.939 4,平均遗传相似系数为0.886 0(表6).‘韦端蜜红’(C1、C2、C3)间的遗传相似系数为0.915 7~0.939 4,芷江‘颐红’(C4、C5)与古田‘颐红’(C6)间的遗传相系数为0.901 2~0.926 4,均具有较高的遗传相似系数.
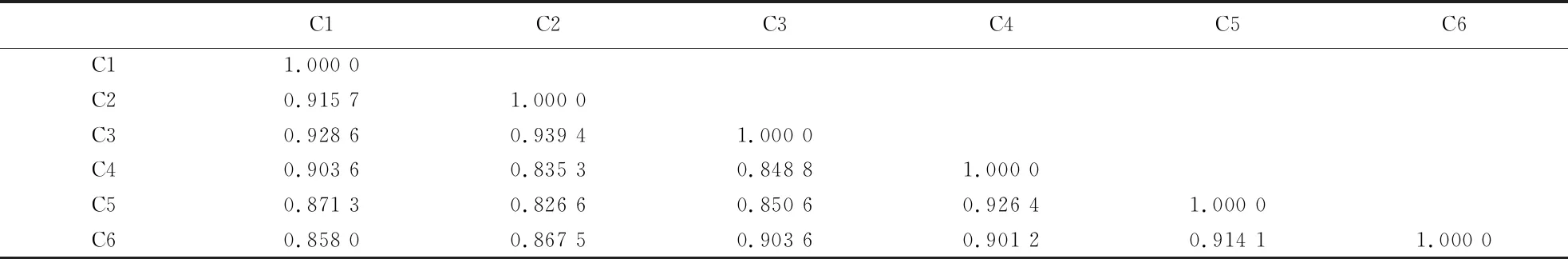
表6 基于ISSR和SRAP标记的6份桃材料之间的遗传相似系数
3 讨论与结论
ISSR分子标记具有RAPD、SSR分子标记的较高多态性、较少的非特异性扩增和操作简捷的特点;而SRAP在保持RAPD、AFLP优点的基础上,更具稳定和简单的特点.本研究比较ISSR、SRAP扩增的总条带数和多态性条带比率,发现ISSR扩增条带总数为91条,多于SRAP的扩增条带总数85条;而SRAP标记多态性为24.71%,高于ISSR标记,此结果与前人[21-23]在其他植物进行的遗传多样性研究结果相一致.通过比较ISSR、SRAP分子标记的遗传相似系数范围,发现SRAP所揭示的遗传相似系数范围(0.807 2~0.935 9)广于ISSR分子标记(0.844 4~0.943 2),SRAP相对于ISSR而言能反映较多的遗传信息,检测到种质间较高的遗传差异,Huang et al[11]和陈大霞等[24]也得出了相类似的结论.‘韦端蜜红’和‘颐红’等6份桃材料的扩增电泳结果显示所有材料之间具有相同的主扩增带,说明桃品种(系)间具有一定的同源性,同时在DNA水平上存在差异.ISSR呈显性遗传因子,不能直接区分显性纯合基因型和杂合基因型[25],而SRAP则是共显性标记,二者的扩增序列和原理均存在差别,将ISSR和SRAP标记结合使用,可以达到优缺点互补的目的,也可以增加基因组的覆盖面积,实现更完整的遗传多样性分析[26-27].ISSR、SRAP联合分析发现,在以相似系数0.90为阈值时可区分‘韦端蜜红’和‘颐红’.ISSR、SRAP及其联合分析的结果具有较好的一致性,其聚类结果均显示,相似系数为0.89~0.92时‘韦端蜜红’与‘颐红’被分为两个组,而在相似系数小于0.85时则聚为同一组,说明‘颐红’与‘韦端蜜红’之间在具有较高遗传相似度的同时也存在一定的遗传差异.
陈小梅等[28]对‘韦端蜜红’、‘颐红’的感官品质和营养品质进行了测定,发现‘韦端蜜红’和‘颐红’的花期均集中在3月中上旬,‘颐红’的成熟期在7月10—30日,而‘韦端蜜红’的成熟期在7月20日至8月10日,比‘颐红’晚10~15 d成熟;‘韦端蜜红’果大(平均单果重206.58 g)且果顶微凸,果实有缝合线且较明显,而‘颐红’果小(平均单果重190.50 g)且果顶深凹,果实缝合线不明显;‘韦端蜜红’的营养品质高于‘颐红’.
综上所述,ISSR、SRAP标记及其联合分析均显示‘韦端蜜红’与‘颐红’在遗传背景上有较高的相似性,在DNA水平上又存在一定的差异,物候期、外观形态等也存在一定的差异,说明‘韦端蜜红’是由‘颐红’芽变而来,是一个新的晚熟桃优良种质资源.
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
计算机应用与软件(2022年6期)2022-07-12
中国农学通报(2022年12期)2022-06-01
油气地质与采收率(2022年3期)2022-05-20
自然灾害学报(2022年2期)2022-05-10
中国糖料(2022年2期)2022-04-06
昆明医科大学学报(2022年2期)2022-03-29
中国种业(2021年11期)2021-11-25
昆明医科大学学报(2021年6期)2021-07-31
昆明医科大学学报(2021年3期)2021-07-22
