
马铃薯帚顶病毒基因组克隆与侵染性克隆构建
2022-10-18 07:06高艳玲刘佳慧马廷帅毛彦芝张丽莉白艳菊程晓非
中国马铃薯 2022年4期
刘 野,高艳玲,刘佳慧,方 月,马廷帅,毛彦芝,张丽莉,白艳菊,程晓非*
(1.寒地粮食作物种质创新与生理生态教育部重点实验室/东北农业大学农学院,黑龙江 哈尔滨 150030;2.黑龙江省农业科学院经济作物研究所,黑龙江 哈尔滨 150086)
马铃薯(Solanum tuberosum)是世界第四大粮食作物,2019年全球马铃薯产量高达3.68亿t,中国是马铃薯产量最大的国家,年产量为9 000万t[1]。病毒病是影响马铃薯产量与品质的重要因素之一,也是马铃薯种薯退化的主要原因。据报道有40余种植物病毒可侵染马铃薯,并造成危害[2]。马铃薯帚顶病毒(Potato mop-top virus,PMTV)是帚状病毒科(Virgaviridae)马铃薯帚顶病毒属(Pomovirus)的代表成员[3]。感染PMTV的马铃薯植株上部叶片出现倒V型黄化斑纹,基部叶片出现不规则黄色斑块或线状纹,块茎出现坏死弧纹或条纹,严重影响马铃薯的产量、品质及商业价值[4]。PMTV主要通过块茎、粉痂菌(Spongospora subterranea)传播,也可通过机械摩擦传播[5]。PMTV曾在广东、云南和四川等地的块茎中被零星检测到,但尚无在中国马铃薯产区大面积流行和传播的报道[6-8]。鉴于该病毒的强致病性和危害性,中国已将其列入进境植物检疫禁止进境物名录。PMTV的病毒粒子为直杆状,基因组由三条正义单链RNA组成,根据长度分别命名为RNA1、RNA2和RNA3[7]。三条RNA的3′末端没有mRNA类似的多聚腺苷酸尾巴,但含有一个类似tRNA的结构[9]。RNA1长约6.1 kb,编码两个复制相关蛋白;RNA2长约3.1 kb,编码两个病毒外壳蛋白(Coat protein,CP);RNA3长约2.9 kb,编码四个阅读框(Open reading frame,ORF),其中ORF1-3部分重叠,组成保守的三基因盒(Triplegene block,TGB),编码3个运动蛋白,ORF4编码一个具有RNA沉默抑制子功能的富含半胱氨酸蛋白(Cysteine-rich protein,CRP)[10]。本研究报道了一株采自河北省马铃薯块茎的PMTV全基因组序列,对其遗传进化进行了分析,并构建了其侵染性克隆,以期为研究PMTV的致病和传播机制、马铃薯的抗病机制以及建立科学防控策略提供理论基础。
1 材料与方法
1.1 材料
1.1.1 植物材料
感染PMTV的马铃薯块茎采自河北省张北县(由黑龙江省农业科学院克山分院马铃薯资源研究所提供)。
1.1.2 菌株与载体
大 肠 杆 菌(Escherichia coli)DH5α、酵 母(Saccharomyces cerevisiae)Y2H Gold、农杆菌(Agrobacterium rhizogenes)GV3101均 由 本 实 验 保存;pCB301-2u-HDV载体来自浙江大学[11]。
1.1.3 培养基与生化试剂
本研究所用培养基及普通生化试剂均为国产分析纯。
1.2 方法
1.2.1 植物培养条件
野生型本氏烟(Nicotiana benthamiana)种植于植物生长箱中,温度为白天25℃,夜间18℃,相对湿度50%和光周期16∶8(光照∶黑暗)。接种病毒以后,N.benthamiana置于温度18℃的植物生长箱,湿度和光周期不变。马铃薯品种‘大西洋’‘中薯5号’和‘Favorita’组培苗在16℃无菌操作间培养,组培苗移至土里后在23℃的植物生长箱中,相对湿度50%和光周期16∶8。
1.2.2 植物总RNA提取
采用普洛麦格(北京)生物技术有限公司的Eastep®Super总RNA提取试剂盒进行,具体操作根据产品说明书进行。
1.2.3 引物设计
从GenBank下载已报道的PMTV全基因组序列,用Clustal Omega[12]进行多序列比对,根据比对结果在保守位置设计引物(表1)。
1.2.4 cDNA合成、PCR扩增与克隆
反转录采用南京诺唯赞生物科技股份有限公司HiScriptⅢ1stStrand cDNA Synthesis Kit(+gDNA wiper),具体操作根据产品说明书进行。PCR扩增采用50 μL体系,包括25 μL的2×Phanta高保真DNA聚合酶(用于目的片段扩增)或2×Taq Plus Master Mix(用于菌落鉴定),10 mmol/L上下游引物各1 μL,cDNA 1 μL。PCR程序设为95℃预变性3 min,30个热循环(95℃变性30 s,60℃退火15 s,72℃延伸时间为:15 s每1 kb),72℃延伸7 min。pCB301-2μ-HDV载体的线性化PCR后,加入1 μLDpnI(NEB北京有限公司),混匀,37℃处理1 h,以降解模板质粒。PCR产物用1%琼脂糖凝胶电泳,用南京诺唯赞生物科技股份有限公司的FastPure Gel DNA Extraction Mini Kit回收扩增片段,回收的片段插入北京全式金(Transgen)生物技术有限公司pEASY-Blunt Zero载体,转化大肠杆菌DH5α感受态细胞,PCR鉴定阳性菌落,用南京诺唯赞生物科技股份有限公司的FastPure Plasmid Mini Kit提取质粒,并送至上海生工生物工程有限公司进行序列测定,每个扩增片段最少测序4个克隆。
1.2.5 序列分析
克隆的DNA片段用Lasergene DNAstar7.0软件包(DNAstar公司)Seqman Pro7.1.0进行拼接,用Seqbuilder Pro7.1.0进行ORF预测和蛋白翻译。系统进化树采用MEGA-X软件中的最大似然法(Maximum-likelihood method),分支支持率采用1 000次重复检测。
1.2.6 酵母同源重组
将线性化的pCB301-2μ-HDV载体与目的片段以等摩尔比混合后,用北京酷来搏科技有限公司的毕赤酵母感受态制备与转化试剂盒转化酵母。转化产物经低速离心后,全部涂布于缺色氨酸的SD固体培养基,用Parafilm封好口后,倒置于30℃培养2~4 d。
1.2.7 酵母质粒提取
采用北京酷来搏科技有限公司酵母质粒小提试剂盒进行,具体操作根据产品说明书进行。
1.2.8 农杆菌转化与侵染
将pCB301-PMTV-RNA1、pCB301-PMTV-RNA2和pCB301-PMTV-RNA3通过电击法分别转入农杆菌GV3101。取15 μL转化产物,涂布于含有50 μg/mL卡那霉素(Kanamycin)和50 μg/mL利福平(Rifampicin)的LB固体培养基,28℃培养2 d;挑取单克隆进行菌落PCR验证。用枪头将含有正确质粒的农杆菌用牙签接种于3 mL含有50 μg/mL卡那霉素和50 μg/mL利福平的LB液体培养基中,28℃摇床中120 r/min过夜培养;室温下,9 000 r/min离心1 min收集菌体。用1 mL浸润缓冲液(10 mmol/L MES,pH 5.6;10 mmol/L MgCl2;100 μmol/L乙酰丁香酮)悬浮菌体,9 000 r/min离心1 min收集菌体;再用1 mL浸润缓冲液悬浮与离心一次,用可见分光光度计将农杆菌稀释至OD600=0.5;取等量含有pCB301-PMTV-RNA1、pCB301-PMTV-RNA2和pCB301-PMTV-RNA3的农杆菌悬浮混匀后,用1 mL无针头注射器注射接种本氏烟草与马铃薯叶片。
2 结果与分析
2.1 全基因组克隆与同源性分析
研究所用的马铃薯块茎品种未知,内部呈现坏死弧纹症状,经ELISA检测为PMTV阳性。根据GenBank中PMTV的序列设计表1引物以扩增该PMTV分离物的全基因组。经克隆、测序和拼装发现该PTMV的RNA1全长6 042 nt,RNA2全长3 134 nt,RNA3全长2 964 nt(GenBank序列号为OP221272-OP221274)。RNA1-3的编码框与报道完全一致,但RNA3编码的CRP前两个密码子分别为GTG和GTG。分析发现,GenBank中所有33条PMTV全长RNA3仅有3个PMTV分离物(Sw、Yunnan和C52_P11)编码CRP的第二个密码子为ATG,而非GTG,因此CRP可能通过非ATG起始翻译。将本PMTV的RNA1、RNA2和RNA3分别与GenBank数据库中不同PMTV分离物的相应RNA片段进行多序列比对,并分析计算同源性。结果发现,本PMTV的RNA1与RNA2和中国Yunnan分离物的核苷酸同源性最高,分别为99.80%和99.90%,RNA3与哥伦比亚的CO3核苷酸同源性最高,为99.90%(表2)。

表2 河北分离物与其他分离物核苷酸序列同源性Table 2 Nucleotide sequence identities between Hebei and other PMTV isolates
2.2 系统进化分析
为明确该PMTV分离物与其他PMTV分离物的系统进化关系,从GenBank数据库中下载了所有PTMV RNA1、RNA2和RNA3全长序列(表1),并构建了RNA1、RNA2以及RNA3的最大似然系统进化树。结果发现,PMTV的RNA1可明显分为两个分支,本PMTV分离物的RNA1处于第一个分支中,与PMTV云南分离物关系最近(图1A);RNA2分为三个分支,绝大多数分离物处于第一分支,而分支III仅包括秘鲁的J20_J203一个分离物,本PMTV分离物的RNA2处于第一个分支中,与中国Yunnan、日本Hiroshima、丹麦54-8、54-11、54-21以及54-26聚于同一小分支(图1B);RNA3分为四个分支,分支III包括瑞典Sw和云南分离物,而分支IV仅包括秘鲁的J20_P157一个分离物,本PMTV分离物的RNA3处于第一个分支中,与丹麦Rn23分离物关系最近(图1C)。

图1 PMTV最大似然系统进化树Figure 1 Maximum likelihood phylogenetic trees of PMTV

表1 试验所用引物序列Table 1 Primers used in the study
2.3 侵染性克隆构建
根据PMTV RNA1、RNA2和RNA3的5′和3′最末端非编码区的保守序列,设计引物以直接扩增RNA2和RNA3全长序列,RNA1分为2个约3 000 bp的片段(分别记为RNA1-F1和RNA1-F2)扩增(图2A)。用引物RZ-F和CaMV 35S-R,以pCB301-2μ-HDV为模板,进行PCR扩增,获得线性化的pCB301-2μ-HDV载体(图2A)。所有片段两端与载体含有20 bp左右的同源臂。将线性化的pCB301-2μ-HDV载体分别与RNA2、RNA3、以及RNA1-F+RNA1-F2共转化酵母。用引物35SF和RNA1-5360R对随机选择的15个转化RNA1-F+RNA1-F2的酵母克隆进行菌落PCR检测,结果发现有6个克隆能扩增到5 000 bp大小的目的条带(图2B),用引物RNA2-620F和NOS-R对随机选择的15个转化RNA2的酵母克隆进行菌落PCR检测,结果表明其中有5个克隆能扩增到2 500 bp左右的目的条带(图2C),用引物RNA3-800F和RNA3-2200R对随机选择的15个转化RNA3的酵母克隆进行菌落PCR检测,结果发现有13个克隆能扩增到1 400 bp左右的目的条带(图2D)。分别选取PCR扩增条带最亮的3个克隆,提取酵母质粒,直接转化E.coliDH5α。经再次菌落PCR验证后,提取质粒送于公司测序,获得含有PMTV河北省马铃薯分离物RNA1、RNA2和RNA3的质粒,并命名为pCB301-PMTV-RNA1、pCB301-PMTV-RNA2和pCB301-PMTV-RNA3。
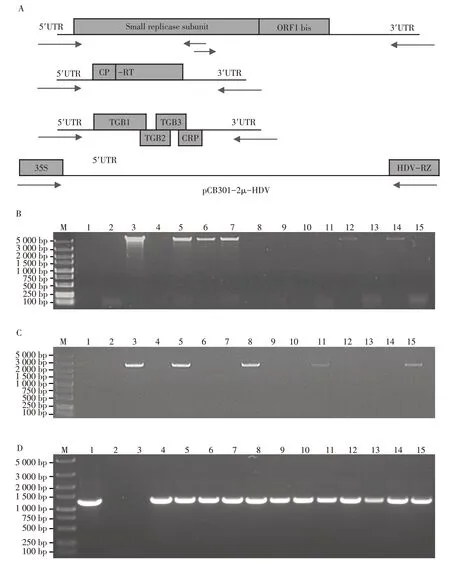
图2 PMTV侵染性cDNA克隆的构建Figure 2 Construction of PMTV infectious cDNA clone
2.4 PMTV侵染性克隆在本氏烟上的侵染性测定
用本氏烟验证所构建PMTV侵染性克隆的活性。将含有pCB301-PMTV-RNA1、pCB301-PMTVRNA2和pCB301-PMTV-RNA3的农杆菌菌液等比例混合后,注射6株4周大小的本氏烟。接种后9 d时,本氏烟的系统叶片开始出现褪绿症状,随着时间的延长,症状逐渐加重,并出现明显的花叶和叶片轻微皱缩的症状(图3A)。在接种14 d,采集本氏烟带有症状的叶片,提取总RNA,分别用引物RNA1-730F+RNA1-5360R、RNA2-620F+RNA2-2800R、RNA3-800F+RNA3-2200R进 行RT-PCR,以 检 测RNA1、RNA2和RNA3。RT-PCR结果显示,在6株本氏烟叶片中均可以检测到病毒RNA1、RNA2和RNA3的存在(图3B~D),表明PMTV的侵染效率达到100%(图3B~D)。

图3 PMTV侵染性cDNA克隆在本氏烟上的侵染性测定Figure 3 Infectivity analysis of PMTV infectious cDNA clone on N.benthamiana
2.5 PMTV侵染性克隆在马铃薯上的侵染性测定
将含有pCB301-PMTV-RNA1、pCB301-PMTVRNA2和pCB301-PMTV-RNA3的农杆菌菌液等比例混合后,分别注射‘大西洋’‘中薯5号’和‘Favorita’无毒组培苗(每个品种各6株),以进一步检测PMTV侵染性克隆对马铃薯的侵染能力。接种20 d后,‘大西洋’叶片出现轻微的黄化,而‘费乌瑞它’和‘中薯5号’没有明显症状(图4A)。接种20 d时,采集组培苗上部带有症状的叶片,提取6株混合样的总RNA,用引物RNA2-1230F与RNA2-2800R进行RT-PCR检测,结果发现‘大西洋’中可以扩增到PMTV RNA2的目的片段,而‘费乌瑞它’和‘中薯5号’中不能扩增到预期大小的目的片段(图4B)。分别提取接种PMTV的6株‘大西洋’总RNA,用引物RNA2-1230F与RNA2-2800R进行RT-PCR检测,以分析PMTV的侵染效率。结果发现4株‘大西洋’可以检测到病毒的存在,发病率为66.67%(图4C)。

图4 PMTV侵染性cDNA克隆在马铃薯上的侵染性测定Figure 4 Infectivity analysis of PMTV infectious cDNA clone on potato
3 讨论
本研究从一个采自河北省张北县的马铃薯薯块上克隆了一株PMTV的全基因组序列,多序列比对和系统进化树分析表明,PMTV的RNA1与此前报道一致分为两个分支[13,14],RNA2分为3个分支,该分离物的RNA1与RNA2均与国内报道的云南分离物关系最近,说明本研究所用马铃薯上的PMTV有可能通过云南与河北之间的种薯调运而来,但还需进一步研究确定。值得注意的是,系统进化分析发现该分离物的RNA3与来自瑞典的Rn23关系最近而与云南马铃薯分离物的进化关系较远,说明本试验克隆的PMTV的RNA3曾与其他分离物的RNA3发生过重配,但是重配发生的时间及地点未知。
PMTV侵染性克隆之前已有报道[15]。本试验构建的PMTV侵染性克隆与之相比,具有不需要依赖于体外转录的优点。此外,本侵染性克隆来自于中国本土发生的带病马铃薯,而不是PMTV代表株系Sw。侵染性试验表明本分离在本氏烟上造成花叶的症状,而Sw造成黄花叶的症状[15],说明两者在致病性方面存在明显差异。马铃薯侵染试验表明,在本试验的条件下,该侵染性克隆能侵染‘大西洋’,但不能引起明显的症状,也不能侵染‘中薯5号’与‘费乌瑞它’,说明本侵染性克隆在不同的马铃薯品种上侵染力不同。温度在PMTV致病过程中具有重要的作用,低温有利于病症的发展和显现[16]。本研究未能在‘大西洋’上观察到症状,可能是由于温度等条件所限。而不能侵染‘中薯5号’与‘费乌瑞它’的原因未知,还需进一步试验验证。
PMTV为三分体病毒,成功侵染需要3条RNA在同一细胞表达。本试验所构建的侵染性克隆虽然在本氏烟上实现100%的侵染效率,但侵染‘大西洋’时只有66.67%(4/6)的侵染率,后续可考虑将病毒的两条RNA整合到一个质粒中,以提高马铃薯的侵染效率。本试验构建的侵染性克隆成功在烟草与马铃薯建立侵染,为进一步开展PMTV与寄主互作的研究,揭示其分子致病机制奠定基础。
猜你喜欢
当代水产(2022年1期)2022-04-26
黑龙江大学自然科学学报(2022年1期)2022-03-29
中国农业科学(2021年23期)2022-01-14
计算机系统应用(2021年10期)2022-01-06
中国瓜菜(2020年8期)2020-09-26
中国瓜菜(2019年8期)2019-09-19
学生天地(2019年28期)2019-08-25
名人传记·财富人物(2017年9期)2017-11-02
名人传记·财富人物(2017年9期)2017-11-02
Coco薇(2016年8期)2016-10-09
