
滇中矮杨梅种群结构与动态特征*
2022-10-26 01:11杨艳波蔡起航巩合德
西部林业科学 2022年5期
杨艳波,蔡起航,巩合德
(1.西南林业大学 地理与生态旅游学院,云南 昆明 650224;2.西南林业大学 园林与园艺学院,云南 昆明 650224)
矮杨梅(Myricanana)属杨梅科(Myricaceae)杨梅属(Myrica),广泛分布于云南中部、贵州西部等地,抗污染和耐旱特性显著。矮杨梅在保持水土和增强土地肥力方面表现突出,这源于其能与根瘤菌共生,具有较强的固氮能力。良好的生态效益使得其成为矿区摞荒地植被恢复的先锋树种以及原始林的优良保留树种;同时,其根茎皮均可入药,在治疗内外伤方面都有一定的作用,具有较高的食用、医药和轻工业等方面的经济价值。同许多野生植物资源一样,很多野生矮杨梅种群都遭到严重破坏,甚至在一些地区濒临灭绝,对野外矮杨梅种群的保护迫在眉睫[2]。
目前,国外学者针对矮杨梅研究较少,国内学者的研究主要集中在遗传多样性[3]、根茎叶化学成分[4-6]、生物量分配特征[7]、碳密度分配特征[8]等方面;近年来,植被动态的相关研究领域由从前的单一演替扩展到更新、动态、边缘效应及进化等方面[9]。国外学者对于群落动态发生变化的原因存在一些争议,有的学者认为自然干扰,如火灾、干旱、洪涝等自然灾害导致了森林动态及其组成的变化,有的学者认为群落动态变化是由环境变化所引起的,如长时间干旱会引起一定范围的物种大规模死亡,还有些研究者认为全球范围的诸多因素正改变着森林的现状[10-11]。国内学者在群落结构及动态特征方面的研究主要集中在高寒草甸[12]、次生针阔混交林[13]、温带落叶阔叶林[14]等群落,此外,解婷婷等[15]选择荒漠绿洲过渡带的沙拐枣种群,对其种群结构和动态进行研究,但对于滇中地区矮杨梅灌丛的种群结构和动态方面的研究还未见报道。
因此,通过绘制矮杨梅种群生命表、存活曲线、生存函数曲线,对各项重要参数进行详细分析,可以在一定程度上揭示矮杨梅种群年龄结构、存活率、死亡率等的变化。进而了解其生存现状,根据时间序列预测种群的变化趋势,探讨矮杨梅种群结构特征和动态变化,以及种群更新和维稳机制。以期实现矮杨梅生境维护和削弱干扰因素影响程度,制定合理有效的保护管理、种群恢复和增殖机制提供科学理论支持和参考。
1 研究区概况及研究方法
1.1 研究区概况
调查于2020年6—9月进行,综合考虑样地地域特点、灌丛类型等是否具有代表性、可操作性后,在云南省中部7个市(县)共布设9个主要灌丛采样地(24.16°~25.48° N、101.01°~104.37° E)。采样地地势平缓,盆地、山地与平地相间,海拔1 936 ~ 2 737 m,常年受西南季风的影响,气候温暖湿润,呈典型的亚热带季风气候,年均气温15.6 ~ 16.2 ℃,年均降水量930.5 mm[16],土壤类型以紫色土、红壤、赤红壤为主。滇中地区植被类型主要以亚热带常绿阔叶林、暖温性落叶阔叶林和温性针阔混交林为主。半湿润常绿阔叶林作为亚热带常绿阔叶林的一个植被亚型,广泛分布于滇中高原的山地(海拔1 300 ~ 2 400 m),是当地的特有植被,即地带性代表植被,但目前大部分已被破坏,仅在一些自然保护区仍有分布,残存的多是萌生灌丛状次生植被和次生林,它们与半湿润常绿阔叶林具有相同的乔木树种组成,保护这些灌丛与次生林已成为保护半湿润常绿阔叶林的关键[17]。采样地优势植物种群主要有:铁仔(Myrsineafricana)、矮杨梅、亮毛杜鹃(Rhododendronmicrophyton)、毛脉高山栎(Quercusrehderiana)、金丝桃(Hypericummonogynum)等。植被受干扰类型及程度见表1。

表1 样地概况Tab.1 Sample plot survey
1.2 研究方法
1.2.1 样地设置与调查方法
在滇中地区设置9块调查样地,分别为Q1、Q2、Q3、Q4、Q5、Q6、Q7、Q8、Q9,每块样地大小为25 m×25 m,根据样地特征,在样地内随机选择3个5 m×5 m的样方进行调查,逐一记录样方内所有矮杨梅个体的株数、基径、高度、胸径、冠幅等指标,并记录样地经纬度、海拔、坡度等数据。
1.2.2 大小级划分
生长在同一生境中的同一树种,环境对其径级和龄级结构具有同样的影响作用,因此,在对数量稀少的濒危树种以及其他获取年龄较困难的树种进行研究时,径级结构代替龄级结构的划分方法是大多数研究者所选用[18]。基于本次矮杨梅的实际调查情况,依据其生物学特性使用矮杨梅个体体积的立方根进行大小级划分,该划分方法可以更好地反映矮杨梅灌丛盖度、生物量及发展趋势等特性,更好的表达矮杨梅种群结构特征[19]。计算样地内每株矮杨梅的冠面积A(长冠幅z ×短冠幅Y)、体积V(冠面积A×高度H)、体积的立方根(d)。通过矮杨梅体积立方根(d)的分布状况,共分为7个等级:I级(d<90 cm)、Ⅱ级(90 cm≤d<110 cm)、Ⅲ级(110 cm≤d<130 cm)、IV级(130 cm≤d<150 cm),Ⅴ级(150 cm≤d<170 cm)、Ⅵ(170 cm≤d<190 cm)、Ⅶ(d≥190 cm)。将不同生境下各大小级的矮杨梅个体数进行统计,绘制矮杨梅种群大小级结构图。
1.2.3 种群动态数量化分析
种群动态需要进行定量描述,本研究采用陈晓德[20]的量化方法。
①
②
式中:Vn为该种群从n到n+1级的个体数量变化值;Vpi为整个种群结构的数量变化动态指数;Sn、Sn+1为第n和n+1级的种群个体数;k为种群大小级数量。不考虑未来外部环境干扰对种群动态进行比较时式②适用,当考虑未来外部环境对种群具有干扰时,种群结构动态与大小级数量(k)及各大小级个体数(S)存在相关性,故将式②修正为:
③
④
式中:Vn、Vpi和Vpi依次代表着种群个体数量增长、衰退和稳定的动态关系。
1.2.4 种群静态生命表编制和生存分析
数据处理要选择同一种群在同一时间内的所有个体的年龄数据进行分析,并基于此编制特定时间生命表,也称静态生命表。假设不同大小径级的矮杨梅个体每年呈等量生长,综合样地获取数据,以20 cm为级差,绘制矮杨梅静态生命表,并对表中数据进行分析,详细编制方法参考谢宗强等[21]。
种群生存环境不变是编制静态生命表的前提条件,但在现实中环境变化的不可控性导致生命表存在一定局限性,为了解矮杨梅种群生存规律,引用4个函数对该种群动态特征进行分析,并依次绘制相应函数曲线,函数公式参考杨凤翔等[22]。
生存率函数:S(i)=S1×S2×…×Si
⑤
累积死亡率函数:F(i)=1-S(i)
⑥
⑦
⑧
式中:hi为大小级宽度,qi为死亡频率。
1.2.5 种群数量动态时间序列分析
对矮杨梅种群采用时间序列分析的一次移动平均法[23],以预测和模拟其未来的大小级结构。
⑨
式中:n为需要预测的未来时间年限;t为大小级,是近期n个值在t时刻的平均值,称为第n周期的移动平均,表示未来n年时t大小级的种群大小;Xk为当前k大小级的种群大小。本研究选择对个体在未来2、4、6个大小级生长时间后的数量进行预测。
2 结果与分析
2.1 矮杨梅种群结构及动态特征
矮杨梅种群大小级结构呈单峰型(图1)。
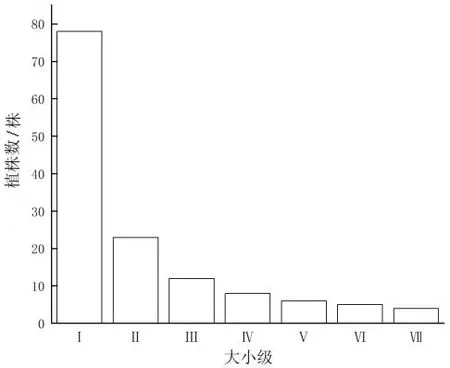
图1 矮杨梅种群大小级结构Fig.1 Size class structure of M.nana population
经统计,矮杨梅种群个体总数为136株,Ⅰ大小级为幼龄期,个体数最丰富,有78株,占总个体数的57.35%;Ⅱ~Ⅳ大小级为成年期,占31.62%,Ⅴ~Ⅶ大小级仅占11.03%,种群大小级结构只有一个峰值(Ⅰ大小级时),为单峰型结构,个体从幼龄期过渡到成年期时数量明显减少,即幼体大量死亡,减少趋势贯穿矮杨梅整个生长期,但总体趋于稳定。

表2 矮杨梅种群大小级结构动态变化指数Tab.2 Dynamic change index of size-class structure of M.nana population
2.2 矮杨梅种群静态生命表及存活曲线
静态生命表的编制是以调查种群内所有个体的具体数据为基础,本文研究的矮杨梅灌丛常年生长在人为干扰较轻微的野外环境下,采集的数据可能存在一定的系统抽样误差,但不影响记录其生态学特性。由表3可知,矮杨梅个体存活数lx会随着大小级的增加而逐渐减少。个体期望生命值ex总体呈先增后减的趋势,在Ⅲ级时期望值最大(ex=2.250),Ⅵ级时最小(ex=0.900),说明Ⅲ级矮杨梅个体的平均生存能力最强,Ⅵ级最弱。
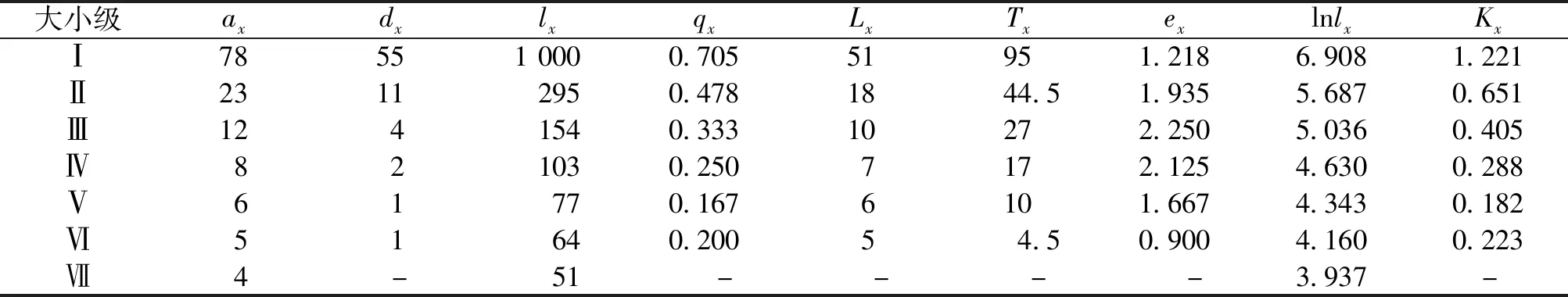
表3 矮杨梅种群静态生命表Tab.3 Static life table of M.nana population
存活曲线的绘制需要通过种群存活个体数对特定年龄死亡率的描述来进行,本研究分别以大小级和标准化存活数lx为横、纵轴作图(图2a)。存活曲线共有3种类型:Deevey-Ⅰ型种群幼龄期个体死亡率低、生存率最高,老年期相反,该型曲线表现为凸曲线;Deevey-Ⅱ型种群各龄级死亡率基本相同,曲线呈对角线;Deevey-Ⅲ型种群幼龄期个体死亡率最高,该型曲线呈凹曲线。本研究采用Hett和Loucks[24]的数学模型进行验证,以确定种群存活曲线属于曲线还是Deevey-Ⅲ型曲线。使用指数方程式Nx=Noe-bx描述Deevey-Ⅱ型曲线,使用幂函数式Nx=Nox-b描述Deevey-Ⅲ型曲线(两式中Nx为lnlx的值,x为大小级,No、b为常数),将数据导入SPSS 19.0进行拟合,建立相应模型后对结果进行比较分析。
矮杨梅种群幼龄期个体数较多,但环境筛强度大,大部分幼龄期个体在生长过程中存在着较高的死亡率。后期死亡率不断下降,说明快速生长过程后,矮杨梅种群的环境适应能力不断增强。对2种生存曲线模型进行检验(表4),可知2种模型均达到极显著水平,但较指数函数模型而言,幂函数模型R2值较大、P值较小,故可认为矮杨梅种群存活曲线趋于Deevey-Ⅲ型,即矮杨梅种群幼龄期个体死亡率高于其他龄级。原因可能是矮杨梅Ⅰ级到Ⅱ级生长过程中个体所需生存资源较少,但随着个体对光热、养分等必需资源的需求不断增长,个体间竞争渐强,以及受周边植被的影响加剧,死亡率维持一个较高水平,使得较少的矮杨梅个体进入到Ⅲ级,进而导致Ⅲ~Ⅶ级个体数量较少。
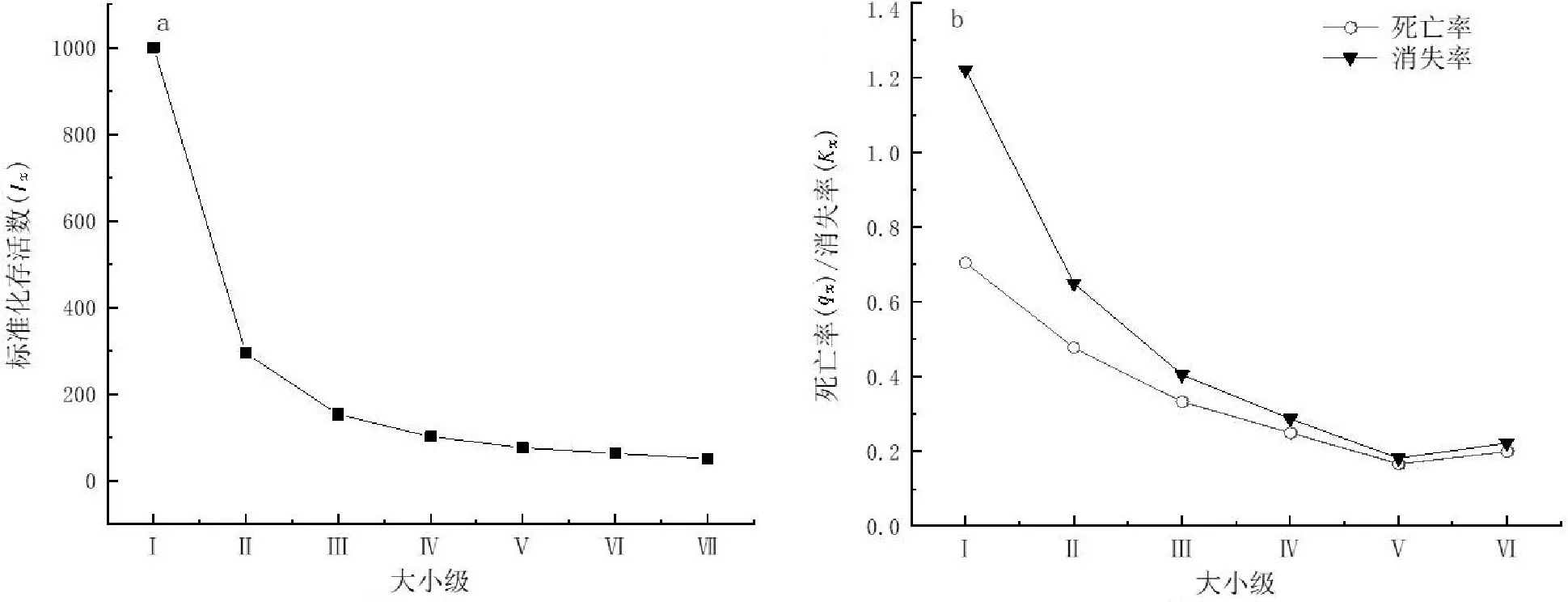
图2 矮杨梅种群存活曲线、死亡率及消失率曲线Fig.2 Curve of survival,mortality and disappearance rate of M.nana population

表4 矮杨梅存活曲线检验模型Tab.4 Test model of survival curve of M.nana population
由图2b可知,矮杨梅种群死亡率(qx)与消失率(Kx)呈相同趋势,峰值都出现在Ⅰ级(qx=0.705,Kx=1.221),二者在Ⅰ~Ⅴ级时呈现出明显下降趋势,与存活曲线表现出的趋势相一致,但在Ⅴ级后二者又出现了小幅增加,由Ⅴ级的0.167、0.182分别增加到Ⅵ级的0.2、0.223,这与矮杨梅个体达到寿命上限有关。
2.3 生存分析
矮杨梅种群各项生存函数估计值见表5、曲线见图3。
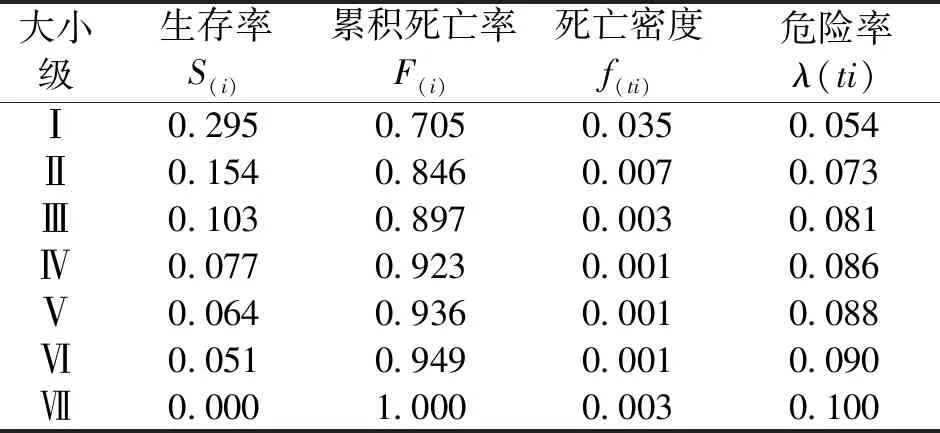
表5 矮杨梅种群生存函数估计值Tab.5 Estimated values of survival functions of M.nana population
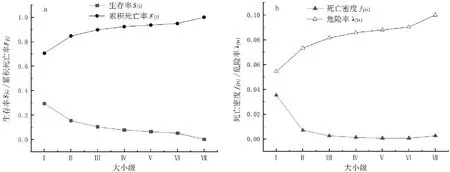
图3 矮杨梅种群生存率S(i)和累计死亡率 F(i)、死亡密度 (ti)和危险率 λ(ti)曲线 Fig.3 Survival rate S(i) ,cumulative mortality rate F(i) ,mortality density (ti) and hazard rate λ(ti) curves of M.nana population
矮杨梅种群生存率S(i)逐级下降,Ⅰ级时值最大〔S(i)=0.295〕,说明Ⅰ级时存活个体最多;累积死亡率F(i)呈上升趋势,Ⅰ~Ⅱ级时死亡率明显上升,表明幼龄期矮杨梅个体大量死亡,只有少部分个体顺利过渡到下一生长期,Ⅱ~Ⅲ级时S(i)和F(i)达到平衡;死亡密度f(ti)呈先降后升的趋势,峰值出现在Ⅰ级〔f(ti)=0.035〕,Ⅵ-Ⅶ级时出现了小幅增加,这与矮杨梅个体达到寿命上限有关;危险率λ(ti)呈缓慢上升趋势,最大值在Ⅶ级〔λ(ti)=0.100〕。
2.4 时间序列预测
以矮杨梅各大小级原始株数数据为基础,对2、4、6个大小级生长时间后各级个体数进行时间序列预测。通过表6可以看出,各矮杨梅种群个体数自Ⅱ大小级开始均呈现出小幅增加的趋势,不考虑外界干扰的情况下,Ⅱ级个体数量在经过2个大小级生长时间后,由原始数据的23、12、8、6、5、4分别增加到51、18、10、7、6、5,分别增长119.57%、45.83%、25%、16.67%、10%、12.5%,表明矮杨梅幼体在总数中占比高,为种群生长中后期提供一定的数量补充。但各大小级到生长后期,个体数量的增长率不断下降,说明在后续生长过程中存在一定损耗,需加强对矮杨梅种群生长的抚育措施,以增加矮杨梅大小级中后期的个体数量,维持其种群的稳定性。
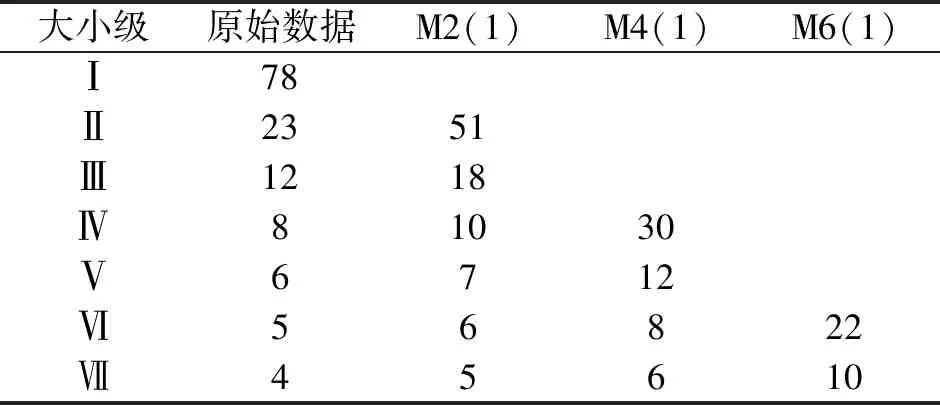
表6 矮杨梅种群数量动态的时间序列预测Tab.6 Time series prediction of population dynamics of M.nana population
3 讨论与结论
分析植物种群结构可以得到种群动态及其发展趋势,并反映不同生境与种群之间相互作用关系。本研究中,矮杨梅种群大小级结构呈明显的单峰型,Ⅰ级时种群个体数量达到峰值,生长后期数量明显减少,但趋于稳定,有向稳定化转化的趋势,作为当地的特有种,矮杨梅喜阴耐旱的特性体现出对本研究区很强的适应性。矮杨梅种子体积较大,不易扩散,大部分种子散布在母体附近,在生长早期同种群间竞争弱,Ⅰ、Ⅱ级个体数量明显多于其他大小级个体数,但幼龄个体对环境胁迫抗性弱,易受到人类、兽类采食以及生境变化的影响,数量不断减少。由生存函数分析可知,大部分Ⅰ级至Ⅲ级个体无法顺利过渡到生长中后期,种群中龄级得不到充足的数量补充,导致中老龄级个体数量明显少于幼龄期个体。待个体生长到中期时对生存资源的需求不断增加,且需要更多的生存空间,因此与同种个体以及其他种个体间产生竞争激烈,保持着较高的死亡率,个体数量较少,矮杨梅种群结构呈现出衰退趋势,种群的更新繁衍受到明显限制,但总体趋于稳定。当少量个体生长到后期,一部分Ⅵ级个体已达寿命上限,出现自然死亡,死亡率上升,因此矮杨梅老龄个体数量较少。
对种群生命表及存活曲线进行分析,可以获得种群实际生存状况,同时找到该种群与环境间的相互关系,对保护与利用珍稀濒危植物以及地方特有种资源有重要意义。本研究发现矮杨梅种群存活曲线为Deevey-Ⅲ型,即种群早期死亡率较高,与王电杰等[25]对浙江省大鹿岛木麻黄(Casuarinaequisetifolia)种群的研究结果相一致,但与赫红敏等[26]在水蚀风蚀交错带对黑沙蒿(Artemisiaordosica)灌丛斑块种群的研究结果不同。矮杨梅的Ⅰ、Ⅱ级个体大多生长在大灌木四周,对资源需求量少,与周围其他个体共享资源,竞争较弱,此时个体数量最丰富,但其应对环境阻力的适应能力较弱,不利气候与细微环境变化就有可能导致大量个体死亡,使得种群动态易波动,呈现出较高死亡率。当矮杨梅个体生长至中期时,经过环境筛选后,个体生存能力增强,代谢旺盛,地下根系生态位重叠,对空间与资源的需求也不断增加,种间竞争激烈,个体间出现分化,且本阶段的个体最易遭到人为樵采,Ⅲ~Ⅴ大小级个体数量明显减少。此时的矮杨梅个体正处于整个生长期中最主要的繁殖阶段,个体数量的减少严重影响种群的更新繁殖,种群虽逐渐趋于稳定,但极易受到外界因素影响,种群仍显现出衰退趋势。生长后期Ⅵ~Ⅶ级个体由于达到寿命期望值,已接近生长极限,各项生理机能逐渐下降,出现死亡情况,这与矮杨梅种群结构分析结果一致。
时间序列预测结果与生命表、存活曲线、动态变化的分析结果一致,在2、4、6个大小级后矮杨梅个体数呈小幅增长的趋势,说明矮杨梅种群存在一定的个体恢复能力,若不进行人工干预,维持种群现有生境稳定,矮杨梅种群可保持数量稳定,但人工管理和育种手段介入更利于矮杨梅种群的长期发展,使其生态、经济效益达到最大化。这与秦岭中段巴山冷杉(Abiesfargesii)的研究结果相似[27],但与秦岭冷杉(A.chensiensis)不同[28]。加强对矮杨梅种群及其他伴生种生境的修复、管理,推动天然状态下矮杨梅种子传播与萌发,适当使用人工辅助更新的方式,增强其繁殖速率和分布范围,以维持种群稳定性和自我恢复机制。
猜你喜欢
林业资源管理(2022年6期)2023-01-18
公民与法治(2022年6期)2022-07-26
现代园艺(2021年23期)2021-12-01
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
文苑(2019年20期)2019-11-16
西江月(2018年5期)2018-06-08
宝藏(2018年1期)2018-04-18
运动(2017年1期)2017-09-03
湖北农业科学(2017年11期)2017-07-13
