
甘蔗野生种割手密杂交F1代SSR鉴定和遗传分析
2022-11-11 02:40田春艳董立华吴才文郎荣斌俞华先桃联安经艳芬
热带作物学报 2022年10期
田春艳,边 芯,董立华,吴才文,郎荣斌,俞华先,张 钰,桃联安,经艳芬*
甘蔗野生种割手密杂交F1代SSR鉴定和遗传分析
田春艳1,边 芯1,董立华1,吴才文2,郎荣斌1,俞华先1,张 钰1,桃联安1,经艳芬1*
1. 云南省农业科学院甘蔗研究所瑞丽育种站,云南瑞丽 678600;2. 云南省农业科学院甘蔗研究所/云南省甘蔗遗传改良重点实验室,云南开远 661699
割手密具有宿根性好、抗逆性强和适应性广等特性,是甘蔗育种中利用最多,育种成效最显著的野生种。进一步发掘和利用割手密优良抗逆基因资源对现代甘蔗品种改良具有重要意义。为获得更多优良割手密的真实杂交后代,本研究以2个地方种为母本、3个野生割手密为父本进行杂交,获得3个F1群体,共359份杂交后代。并从21对SSR标记中筛选出在双亲中具有多态性且扩增条带清晰的6对SSR标记,基于高通量的荧光毛细管电泳检测平台进行杂交后代真实性鉴定、亲本指纹图谱分析、遗传相似性分析和特异性条带的遗传分析。结果表明:共筛选出的6对SSR标记对5个亲本材料的分辨率较高,扩增多态性好,每个亲本都具有其特异的SSR指纹,可有效鉴定其杂交后代血缘的真实性;其中,359份F1个体中共鉴定出真杂种262份,3个组合的真杂种率分别为67.77%、75.51%、75.66%,平均值为72.98%。遗传相似性分析表明2个地方种的遗传相似性系数为0.70,3个割手密两两间的遗传相似性系数分别为0.53、0.60和0.70,地方种和割手密间为0.38~0.53,种间遗传相似性系数小于种内。亲本特异性SSR位点的遗传分析结果表明,3个组合的母本特异性条带遗传率分别为68.47%、80.96%、73.39%,平均值为74.27%,而父本特异性条带遗传率分别为58.90%、76.60%、61.45%,平均值为65.65%,杂交后代具有偏母本遗传倾向,因此,在甘蔗杂交育种中应选择综合农艺性状较好的材料作为母本。本研究结果为今后割手密杂交后代的鉴定提供了高效、可靠的SSR标记选择,同时鉴定出的真实割手密后代群体可为开展割手密优异性状的遗传研究提供种质材料,为选育超亲遗传株系提供科学依据。
甘蔗;割手密;SSR;杂种鉴定;遗传
有性杂交育种是甘蔗育种上最常用、成效最显著的方法,在我国育成的甘蔗品种中,通过有性杂交育成的甘蔗品种占98%以上[1]。而作物种质资源,尤其是优异种质的发掘和利用是作物育种取得突破的关键[2]。玉米和水稻的育种经验表明,每一次粮食的重大增产皆得益于少数几个关键种质的发掘和利用[3]。
早在20世纪初,印尼利用爪哇割手密(L)与热带种()杂交选育出‘POJ2878’,成为风靡全球的蔗王。印度利用印度割手密与热带种、印度种()杂交,选育出‘Co213’‘Co281’‘Co290’等全球性著名的亲本或品种,现今绝大多数甘蔗商业品种均是含有‘POJ’‘Co’系列品种的血缘[4-5]。割手密成为了甘蔗育种史上最重要的种质资源之一,是甘蔗属内利用最多,育种成效最显著的野生种。然而,甘蔗是典型的中日照作物,只有在特定光照时间段(12~12.5 h)才能开花[6]。在我国,除海南岛以外的大部分地区甘蔗都不能自然开花,或者虽有少量开花但亲本花期不遇。甘蔗地方果蔗属于难花亲本,即使在人工光周期诱导下开花率也不高且花期较晚,而割手密自然条件下就可开花,且花期较早。地方种开花诱导难、晚花,与割手密杂交花期不遇、杂交结实率低等问题,使其获得后代杂交群体的难度较大,进而影响了甘蔗遗传图谱构建、图位克隆、重要性状相关的QTL定位等工作的开展。因此,构建割手密杂交群体,鉴定出真实杂种可为开展甘蔗遗传研究提供种质基础。
传统鉴定方法多是基于对甘蔗种质的形态特征描述,如根据茎形、芽形、曝光前后节间颜色、叶鞘、叶耳、蜡粉带等性状进行识别,为早期甘蔗种质的鉴定发挥了重要作用[7]。但传统鉴定方法由于涉及的性状众多、受环境条件影响大,耗时耗力且鉴定结果不准确[8-9]。而分子标记因不受环境因素限制,更能客观地反映DNA水平上的差异,已被广泛应用于杂交后代的真实性鉴定和遗传分析研究。其中,SSR(simple sequence repeat)标记因其具有共显性遗传、多态性丰富、分布广泛、重复性好等优点,是目前用于甘蔗杂交后代真假杂种鉴定最有效的标记之一[10-12]。
本研究利用前期从云南本土收集的甘蔗种质资源,以2个地方种作母本,3个割手密作父本进行杂交,创制了3个F1群体,共计359份杂交后代。以此359份F1个体及其亲本为研究材料,利用多态性SSR标记,并结合荧光毛细管电泳(fluorescence-capillary electrophoresis,CE)检测平台鉴定真假杂种,为后续甘蔗遗传图谱构建、重要性状QTL定位等提供作图群体材料,同时探讨地方种和割手密的种间遗传相似性、亲本的SSR位点遗传特性,为甘蔗开展分子标记辅助育种、选育超亲遗传种质奠定基础。
1 材料与方法
1.1 材料
参试材料共364份,包括3个割手密F1群体后代及其亲本。组合A:‘歪娥’ב云南82-114’,F1代有49个,组合B:‘南涧果蔗’ב云南2015-2’,F1代有121个,组合C:‘南涧果蔗’ב云南06-7-3’,共189个F1。其中,‘歪娥’和‘南涧果蔗’是地方种(‘南涧果蔗’也属于甘蔗属热带种资源),‘云南82-114’‘云南2015-2’和‘云南06-7-3’是前期采集的野生割手密。
1.2 方法
1.2.1 基因组DNA提取和质量检测 取200 mg幼嫩叶片,采用天根植物基因组DNA提取试剂盒(DP320-02)提取甘蔗基因组DNA,具体操作步骤参考试剂盒说明书。提取完成后,取2 μL用Thermo微量紫外分光光度计NanoDrop(型号ND1000)检测DNA浓度及质量。取4 μL在1%浓度的琼脂糖凝胶上进行电泳,检测DNA完整性,主带清晰且不大量拖尾的DNA用于后续SSR分型。
1.2.2 SSR分型 根据PAN[13]建立的甘蔗分子身份证数据库构建的方法,从中筛选出在父母本间具有特征性条带的引物6对,用于本研究杂交后代群体的真实性鉴定(表1)。PCR反应体系总体积为15 μL,其中,DNA模板1 μL,10×PCR buffer 1.5 μL,25 mmol/L MgCl21.5 μL,10 mmol/L的dNTPs 0.3 μL,正反向引物各0.15 μL(浓度10 µmol/L),DNA聚合酶(5 U/μL)0.3 μL,ddH2O 10.1 μL。PCR扩增程序设置为:94℃预变性3 min;94℃变性15 s,55℃退火15 s,72℃延伸30 s,这3个阶段循环35次;最后72℃延伸5 min,4℃保存。PCR产物进行琼脂糖凝胶电泳,根据电泳结果估计PCR产物浓度,并将产物稀释10倍后,与ROX 500内标(片段大小分别为50、75、100、139、150、160、200、300、350、400、450、490、500 bp)混匀,置于ABI3730XL基因测序仪样本架上进行毛细管电泳检测,完成后导出SSR基因分型文件。
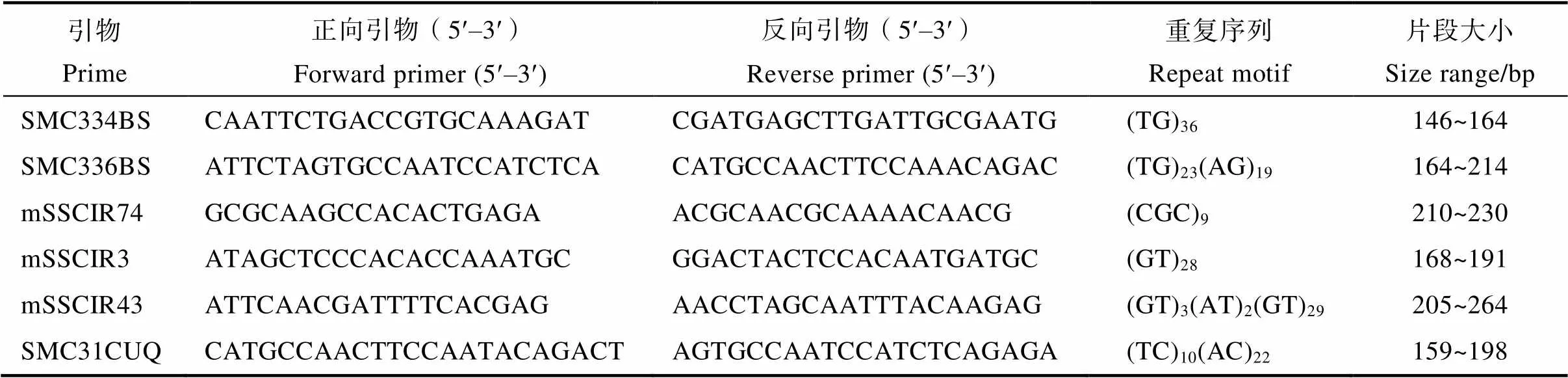
表1 本研究所用SSR引物信息
1.2.3 位点统计和杂交后代真实性鉴定 利用GeneMarker 2.7(Soft Genetics LLC,State College,Pennsylvania,USA)软件对毛细管电泳输出图谱进行扩增片段统计。首先设定每个SSR标记的等位基因panel,系统将根据所设定的panel进行条带统计,某一位点上有条带记为“1”,无条带记为“0”,最后进行人工校对,剔除影子峰和不规则峰。利用NTSYSpc 2.10计算遗传相似性系数。
杂交后代真实性判断方法:根据桃联安等[14]、陆鑫等[15]的方法进行真假杂种鉴定。即杂交后代扩增条带均来自父本和母本,且具有父本特征带的为真杂种;后代扩增条带来自母本和父母本共有带,而无父本特征带的为自交种,需根据其他引物进一步判断;后代出现新条带,即父母本都没有的条带,为假杂种。
2 结果与分析
2.1 SSR标记对3个组合亲本的鉴定效率
标记的选择是杂交后代鉴定的重要环节。从图1可看出,本研究所选用的6个SSR标记在亲本上具有多个特征性条带,可有效鉴别每一个亲本材料。2个母本材料中,标记SMC334BS扩增出2个‘南涧果蔗’特征带(位点17、位点18),SMC336BS扩增出3个‘歪娥’特征带(位点9、位点10和位点11)和2个‘南涧果蔗’特征带(位点4和位点12),mSSCIR74扩增出1个‘歪娥’特征带,mSSCIR3扩增出1个‘歪娥’特征带和3个‘南涧果蔗’特征带,mSSCIR43扩增出3个‘南涧果蔗’特征带,SMC31CUQ扩增出6个‘歪娥’特征带和1个‘南涧果蔗’特征带。每个标记都能较好地鉴定母本‘歪娥’和‘南涧果蔗’。
3个割手密在6对SSR引物上也扩增出了各自的特征带。如‘云南82-114’在SMC334BS、SMC336BS、mSSCIR74、mSSCIR3、mSSCIR43、SMC31CUQ上分别具有特征带6个、1个、3个、2个、2个、3个,可区别于‘云南2015-2’和‘云南06-7-3’的特征带。‘云南2015-2’在标记SMC334BS和SMC31CUQ上具有1个可区别于‘云南82-114’和‘云南06-7-3’的特征带。‘云南06-7-3’在标记SMC336BS、mSSCIR3、SMC31CUQ上分别有特征带2个、4个和2个,可区别于‘云南82-114’和‘云南2015-2’的特征带。此外,从图1可看出,2个母本和3个父本间遗传差异较大,特征带较多,同时也表明本研究所选用的SSR引物可有效鉴定甘蔗地方种和割手密杂交后代的真实性。

黑色框表示该位点有条带;白色框表示该位点无条带。
2.2 杂交后代F1群体真假杂种鉴定
2.2.1 ‘歪娥’与云南82-114杂交后代鉴定结果 由图2可知,6对引物均能扩增出组合A(‘歪娥’ב云南82-114’)父本‘云南82-114’(编号P2)特征带。引物mSSCIR3在176、179、180、182 bp 处扩增出条带,其中176 bp和179 bp为父本(编号P2)与母本(编号P1)共有,180 bp和182 bp为父本特有,无母本‘歪娥’特征带。而其余5对引物均具有1个或多个双亲特征带,如SMC334BS共扩增出14个条带,其中4个为母本特有,10个为父本特有;mSSCIR74共扩增出8个条带,其中3个为母本特有,3个为父本特有,2个为父母本共有。根据杂交后代血缘真实性判断方法和6对SSR引物的扩增结果,组合A的49个F1中,37个为真杂种,12个为假杂种,无自交种,真杂种率为75.51%。
2.2.2 ‘南涧果蔗’与‘云南2015-2’杂交后代鉴定结果 由表2可知,6对SSR引物均能扩增出组合B双亲的特征带。其中,SMC334BS在双亲上共扩增出10个条带,包括5个母本特异带(160、161、162、163、164 bp),4个父本特异带(145、146、149、150 bp),1个父本和母本共有带(159 bp)。SMC336BS在双亲上共扩增出6个条带,3个母本特异带,3个父本特异带;mSSCIR74 扩增出5个条带,3个母本特异带,1个父本特异带,1个父母本共有带(219 bp);mSSCIR3扩增出3个条带,1个母本特异带,1个父本特异带,1个父母本共有带;mSSCIR43扩增出9个条带,5个母本特异带,4个父本特异带;SMC31CUQ扩增出4个条带,1个母本特征带,3个父本特征带。根据杂种鉴定标准和6对SSR引物对组合B后代群体的扩增结果,121个F1中,82个为真杂种,39个为假杂种,无自交种,真杂种率为67.77%。
2.2.3 ‘南涧果蔗’与云南06-7-3杂交后代鉴定结果 组合C的母本为‘南涧果蔗’,父本为割手密‘云南06-7-3’,扩增结果表明,父本和母本的扩增条带差异较大(表2),可有效鉴定其杂交后代血缘的真实性。根据6对SSR引物扩增结果和杂交后代真实性鉴定方法,组合C的189个F1中,143个为真杂种,46个为假杂种,无自交种,真杂种率为75.66%。

P1为母本‘歪娥’,P2为父本‘云南82-114’。

表2 3个组合亲本的扩增条带统计情况
2.3 母本和父本遗传相似性分析
由表3可知,2个母本地方种间遗传相似性系数为0.70,3个父本割手密间遗传相似性系数为0.53~0.70,其中‘云南2015-2’和‘云南06-7-3’遗传相似性系数最大为0.70,‘云南82-114’和‘云南06-7-3’间遗传相似性系数最小,为0.53,‘云南82-114’和‘云南2015-2’间遗传相似性系数为0.60。地方种和割手密种间遗传相似性系数范围为0.38~0.53,小于种内的遗传相似性系数。表明本研究所选用的6个SSR标记对甘蔗地方种和割手密种的分辨率较高,可高效鉴定其杂交后代的血缘真实性。
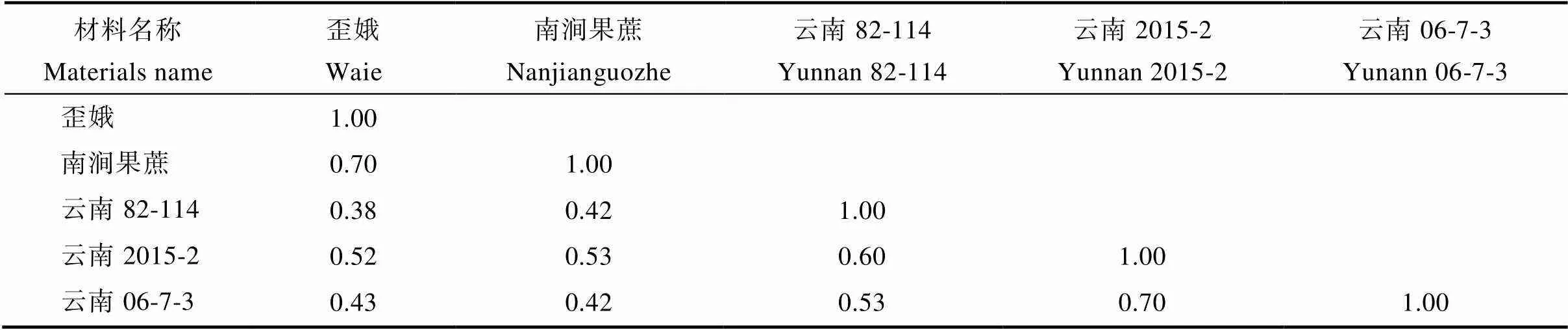
表3 5个亲本材料的遗传相似性系数
2.4 亲本的SSR位点遗传分析
由表4可知,6对SSR 引物在组合A的37个真杂种中共扩增出52个条带,其中18个为母本特有,29个为父本特有,其余为父母本共有带。母本特异性条带遗传率为68.47%,父本特异性条带遗传率为58.90%。组合B有真杂种82个,共扩增出37个条带,其中18个为母本特异性条带,16个为父本特异性条带,其余为父母本共有带;母本特异性条带遗传率为80.96%,父本特异性条带遗传率为76.60%。6对SSR引物在组合C的真实后代群体中扩增出46个条带,20个为母本特异性条带,23个为父本特异性条带,其余为父母本共有带;双亲的特异性条带中,母本条带遗传率为73.39%,父本条带遗传率为61.45%。3个组合的杂交后代中,母本条带遗传率均高于父本条带的遗传率,具有偏母本遗传倾向。
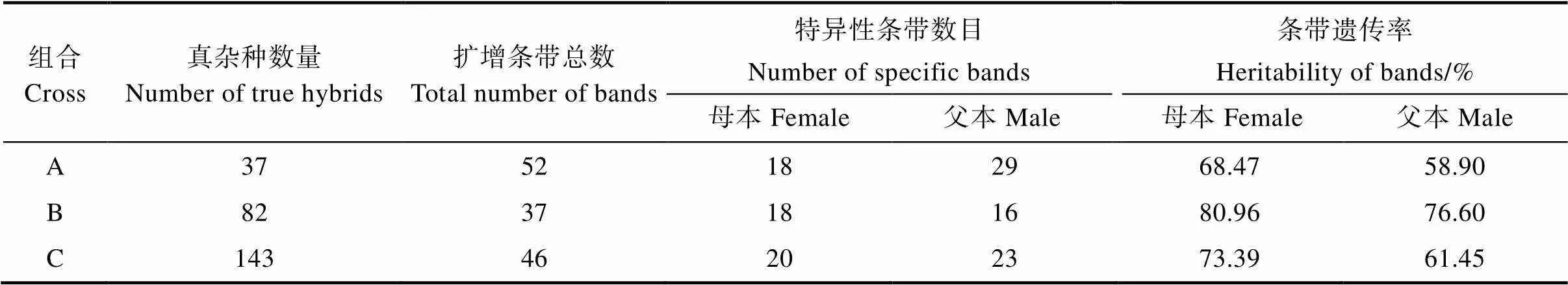
表4 6个SSR标记的亲本特异性条带数目及遗传率
3 讨论
杂交是作物产生遗传变异的重要途径,是新品种选育、群体构建和重要性状QTL定位的前提和基础[16]。在甘蔗杂交过程中,由于人工操作不规范、母本未进行温汤杀雄等导致的未知来源花粉混入和母本自交结实易造成后代的混杂。对杂交后代进行早期筛选,淘汰假杂种是种质创新工作中的重要环节。由于甘蔗是高度复杂的多倍体和非整倍体作物(2n=8x or 10x=100–130)[17-18],杂交后代遗传基础复杂,性状分离广泛,从形态学上很难辨别杂种的真实性。SSR为共显性标记,可区分纯合和杂合,且重复性好、易操作,已被广泛应用于杂种的真实性鉴定[19-20]。
在杂种鉴定中,SSR引物的选择至关重要,不同的引物鉴定效率不同,理论上纯合显性标记只需一个即可鉴定出全部的杂种后代,而杂合的显性标记至少要用5个父本特异标记才能鉴定出97%的真杂种[21-22]。如本研究中6对引物对组合A父母本的扩增结果所示,mSSCIR3扩增出2个父本特异带而无母本特异带,mSSCIR43扩增出1个母本特异带和4个父本特异带,相比其他4对在父母本上均扩增出多个特异带的引物而言,鉴定效率较低。综合6对SSR引物的扩增结果,3个F1群体的真杂种率分别为75.51%、67.77%、75.66%,平均真杂种率为72.98%,与陆鑫等[15]、郭育强等[23]和高轶静等[24]报道的真杂种率为100%相差较大,这主要是由所选用的引物多态性及其数量造成的。周宁宁等[25]研究认为引物的多态性越高,后代扩增出现新条带的几率就越大。本研究选用的6个SSR标记,多态性较高,扩增出了多条父母本特征带。因此,在后代扩增出父母本没有的条带即视为假杂种的情况下,假杂种出现的概率就会提高。然而,吴学蔚等[26]、韩国辉等[22]研究认为,杂交后代扩增出现新条带,能否作为判断杂种的依据,需要根据这些新条带的确切来源判断,并推测新条带的出现可能是由于配子在形成过程中染色体的不等价交换产生。但是,基于甘蔗基因组的复杂程度和杂交后代染色体遗传方式的多样化,目前的技术还很难追踪新条带的确切来源。因此,在甘蔗杂种后代鉴定中,真假杂种的判断和新条带出现的原因还需开展更加深入的研究,这对了解甘蔗杂交后代的遗传具有重要意义。
种质资源是作物遗传育种的基础,开发利用优异的种质资源是进行种质创新、选育突破性品种的关键。同时,亲本间遗传基础的差异与后代群体的遗传变异水平及后期能否获得优良株系密切相关。本研究遗传相似性分析结果表明,3个组合父本和母本的遗传相似性分别为0.38、0.42和0.53,双亲间遗传差异较大,杂交后代更有利于获得广泛的分离和变异。选用的3个父本割手密间遗传相似性分别为0.53、0.60和0.70,也具有一定的多样性和遗传差异。据前人研究报道,甘蔗和割手密杂交,其染色体遗传有2n+n和n+n 两种方式[27],亲本间较大的遗传差异和杂交后代染色体遗传方式的多样性进一步增加了后代的遗传变异水平,提高获得优良育种中间材料和优异株系的可能性,提高育种效应。此外,本研究的SSR位点遗传分析表明,3个组合的后代群体中,其母本条带的遗传率均高于父本条带的遗传率,后代具有偏母本遗传倾向。因此,在今后开展高产高糖新品种选育时,应注重选择具有高产高糖的材料为母本,以提高杂交后代高产高糖性状出现的频率。
[1] 吴才文, 赵培方, 夏红明. 现代甘蔗杂交育种及选择技术[M]. 北京: 科学出版社, 2014: 1-2.
WU C W, ZHAO P F, XIA H M. Modern cross breeding and selection techniques in sugarcane[M]. Beijing: Science Press, 2014: 1-2. (in Chinese)
[2] 刘家勇, 邓祖湖, 吴才文, 桃联安, 陆 鑫, 赵培方, 张跃彬. 甘蔗属野生种割手密的育种利用进展与探讨[J]. 植物遗传资源学报, 2021, 22(6): 1491-1497.
LIU J Y, DENG Z H, WU C W, TAO L A, LU X, ZHAO P F, ZHANG Y B. Progress and discussion of sugarcane breeding usingL[J]. Journal of Plant Genetic Resources, 2021, 22(6): 1491-1497. (in Chinese)
[3] WANG Z T, PAN, Y B, LUO J, YOU Q, XU L P, ZHANG H, QUE Y X. SSR-based genetic identity of sugarcane clones and its potential application in breeding and variety extension[J]. Sugar Tech, 2020, 22: 367-378.
[4] 邓海华. 我国本土割手密育成品种的亲缘关系分析[J]. 广东农业科学, 2012, 39(8): 167-170.
DENG H H. Pedigree analysis of the native spontaneum-derived varieties of sugarcane in mainland China[J]. Guangdong Agricultural Sciences, 2012, 39(8): 167-170. (in Chinese)
[5] SREENIVASAN T V, AHHOWWALIA B S, HEINZ D J. Sugarcane improvement through breeding[J]. Netherlands: Elsevier Science Publishers, 1987, 12: 211-254.
[6] MOORE P H, BOTHA F C. Sugarcane: physiology, biochemistry, and functional biology[M]. New Jersey: John Wiley & Sons Inc, 2014: 384-385.
[7] 毛 钧, 蔡 青, 陆 鑫, 刘新龙, 马 丽, 应雄美. 甘蔗杂交后代真实性鉴定技术发展与应用[J]. 中国糖料, 2008(4): 62-65.
MAO J, CAI Q, LU X, LIU X L, MA L, YING X M. Development and application of sugarcane hybrid reliability identification[J]. Sugar Crops of China, 2008(4): 62-65. (in Chinese)
[8] CORDEIRO G M, PAN Y B, HENRY R J. Sugarcane microsatellites for the assessment of genetic diversity in sugarcane germplasm[J]. Plant Science, 2003, 165: 181-189.
[9] DOS SANTOS J M, DUARTE FILHO L S, SORIANO M L, DA SILVA P P, NASCIMENTO V X, SOUZA BARBOSA G V, TODARO A R, RAMALHO NETO C E, ALMEIDA C. Genetic diversity of the main progenitors of sugarcane from the RIDESA germplasm bank using SSR markers[J]. Industrial Crops and Products, 2012, 40: 145-150.
[10] 刘昔辉, 方锋学, 高轶静, 张荣华, 宋焕忠, 杨荣仲, 方位宽, 段维兴, 罗 霆, 张革民, 李杨瑞. 斑茅割手密杂种后代真实性鉴定及遗传分析[J]. 作物学报, 2012, 38(5): 914-920.
LIU X H, FANG F X, GAO Y J, ZHANG R H, SONG H Z, YANG R Z, FANG W K, DUAN W X, LUO T, ZHANG G M, LI Y R. Identification and genetic analysis of hybrid from cross between(Retz.) Jesws. andL[J]. Acta Agronomica Sinica, 2012, 38(5): 914-920. (in Chinese)
[11] 黄玉新, 罗 霆, 高轶静, 张保青, 周 珊, 杨翠芳, 段维兴, 雷敬超, 韦金菊, 张革民, 李杨瑞. 甘蔗与斑茅割手密复合体的回交后代真实性鉴定及染色体遗传分析[J]. 南方农业学报, 2016, 47(10): 1642-1647.
HUANG Y X, LUO T, GAO Y J, ZHANG B Q, ZHOU S, YANG C F, DUAN W X, LEI J C, WEI J J, ZHANG G M, LI Y R. Authenticity identification and chromosomes genetic analysis of backcross progeny between sugarcane () and intergenetic hybrid complex (×)[J]. Journal of Southern Agriculture, 2016, 47(10): 1642-1647. (in Chinese)
[12] 赖小群, 黄帝媛, 姚 潇, 陈悦佳, 姚姿婷, 邹承武, 陈保善. 应用SSR分子标记鉴定新台糖25号×云蔗89-7 F1代大规模遗传群体的杂种真实性[J]. 中国糖料, 2021, 43(1): 7-12.
LAI X Q, HUANG D Y, YAO X, CHEN Y J, YAO Z T, ZOU C W, CHEN B S. Identification of the F1hybrids facticity from cross between ROC 25 and Yunzhe 89-7 using SSR molecular markers[J]. Sugar Crops of China, 2021, 43(1): 7-12. (in Chinese)
[13] PAN Y B. Databasing molecular identities of sugarcane (spp.) clones constructed with microsatellite (SSR) DNA markers[J]. American Journal of Plant Sciences, 2010(1): 87-94.
[14] 桃联安, 楚连璧, 经艳芬, 刘新龙, 安汝东, 董立华, 杨李和, 周清明, 段惠芬. 云南割手密82-114种间杂交后代SSR分子标记鉴定[J]. 植物遗传资源学报, 2009, 10(1): 132-135.
TAO L A, CHU L B, JING Y F, LIU X L, AN R D, DONG L H, YANG L H, ZHOU Q M, DUAN H F. Identification of genuine hybrids from the cross of chewing cane andin Yunnan by SSR markers[J]. Journal of Plant Genetic Resources, 2009, 10(1): 132-135. (in Chinese)
[15] 陆 鑫, 毛 钧, 刘洪博, 刘新龙, 马 丽, 苏火生, 蔡 青.甘蔗野生种滇蔗茅种质创新利研究Ⅰ.甘蔗与滇蔗茅远缘杂交F1群体构建与SSR分子标记鉴定[J]. 植物遗传资源学报, 2012, 13(2): 321-324.
LU X, MAO J, LIU H B, LIU X L, MA L, SU H S, CAI Q. Innovation germplasm ofI. establishment of F1 intergeneric distant hybridization ofL. andand the identification of F1by SSR molecular markers[J]. Journal of Plant Genetic Resources, 2012, 13(2): 321-324. (in Chinese)
[16] 雷 雨, 段继华, 黄飞毅, 康彦凯, 罗 意, 陈宇宏, 丁 玎, 姚利娜, 董丽娟, 李赛君. 茶树杂交F1代真假杂种的SSR鉴定及遗传多样性分析[J]. 植物遗传资源学报, 2021, 22(3): 748-757.
LEI Y, DUAN J H, HUANG F Y, KANG Y K, LUO Y, CHEN Y H, DING D, YAO L N, DONG L J, LI S J. Identification and genetic diversity of tea F1 hybrids based on SSR markers[J]. Journal of Plant Genetic Resources, 2021, 22(3): 748-757. (in Chinese)
[17] SINGH R K, MISHRA S K, SINGH S P, Mishra N, SHARMA M L. Evaluation of microsatellite markers for genetic diversity analysis among sugarcane species and commercial hybrids[J]. Australian Journal of Crop Science, 2010, 4(2): 116-125.
[18] GRIVET L, ARRUDA P. Sugarcane genomics: depicting the complex genome of an important tropical crop[J]. Genome Studies and Molecular Genetics, 2002, 5(2): 122-126.
[19] 王一帆, 郑 赟, 荣二花, 吴玉香. 亚洲棉与拟似棉远缘杂种的合成与鉴定[J]. 生物技术通报, 2020, 36(8): 35-40.
WANG Y F, ZHENG Y, RONG E H, WU Y X. Synthesis and identification of distant hybrids betweenand[J]. Biotechenology Bulletin, 2020, 36(8): 35-40.(in Chinese)
[20] 杨丽娟, 饶庆琳, 李洪有,王 艳, 冉 盼, 崔娅松, 陈庆富. 荞麦属植物多年生种间杂种的鉴定[J]. 植物遗传资源学报, 2019, 20(2): 387-395.
YANG L J, RAO Q L, LI H Y, WANG Y, RAN P, CUI Y S, CHEN Q F. Identification of perennial interspecific hybrids on genus[J]. Journal of Plant Genetic Resources, 2019, 20(2): 387-395. (in Chinese)
[21] 张开春, 李荣旗, 毕晓颖, 尹淑萍, 吴禄平, 景士西. RAPD技术鉴定无融合生殖型平邑甜茶的有性后代[J]. 农业生物技术学报, 1997, 5(4): 392-396.
ZHANG K C, LI R Q, BI X Y, YIN S P, WU L P, JING S X. Sexual hybrid identification in apomictic “Pingyi Tiancha” seedlings using RAPD markers[J]. Journal of Agricultural Biotechnology, 1997, 5(4): 392-396. (in Chinese)
[22] 韩国辉, 向素琼, 汪卫星, 魏 旭, 何 波, 李晓林, 梁国鲁. 沙田柚杂交后代群体的SSR鉴定与遗传多样性分析[J]. 中国农业科学, 2010, 43(22): 4678-4686.
HAN G H, XIANG S Q, WANG W X, WEI X, HE B, LI X L, LIANG G L. Identification and genetic diversity of hybrid progenies from Shatian pummelo by SSR[J]. Scientia Agricultura Sinica, 2010, 43(22): 4678-4686. (in Chinese)
[23] 郭育强, 符 成,许环映, 常海龙, 吴建涛, 邱永生, 王勤南. 割手密与甘蔗杂交F1群体构建及遗传多样性分子检测[J]. 分子植物育种, 2021, 19(1): 234-239.
GUO Y Q, FU C, XU H Y, CHANG H L, WU J T, QIU Y S, WANG Q N. Construction of F1 populations ofL. and sugarcane and molecular detection of genetic diversity[J]. Molecular Plant Breeding, 2021, 19(1): 234-239. (in Chinese)
[24] 高轶静, 方锋学, 刘昔辉, 张荣华, 宋焕忠, 杨荣仲, 罗 霆, 段维兴, 游建华, 张革民. 甘蔗与斑茅割手密复合体杂交后代的分子标记鉴定[J]. 植物遗传资源学报, 2012, 13(5): 912-916.
GAO Y J, FANG F X, LIU X H, ZHANG R H, SONG H Z, YANG R Z, LUO T, DUAN W W, YOU J H, ZHANG G M. Identification of progeny from crosses between sugarcane (spp.) and intergeneric hybrid complex (×) with molecular markers[J]. Journal of Plant Genetic Resources, 2012, 13(5): 912-916. (in Chinese)
[25] 周宁宁, 李淑斌, 李远波, 蹇洪英, 晏慧君, 王其刚, 陈 敏, 邱显钦, 张 颢, 唐开学. 二倍体月季F1群体的SSR鉴定与遗传分析[J]. 园艺学报, 2017, 44(1): 151-160.
ZHOU N N, LI S B, LI Y B, JIAN H Y, YAN H J, WANG Q G, CHEN M, QIU X Q, ZHANG H, TANG K X. Hybrids identification and genetic analysis in diploid roses population (F1) using SSR markers[J]. Acta Horticulturae Sinica, 2017, 44(1): 151-160. (in Chinese)
[26] 吴学尉, 崔光芬, 吴丽芳, 张艺萍, 明 军, 王 杰, 王继华. 百合杂交后代ISRR鉴定[J]. 园艺学报, 2009, 36(5): 749-754.
WU X W, CUI G F, WU L F, ZHANG Y P, MING J, WANG J, WANG J H. Identification of ISSR in lily hybrids[J]. Acta Horticulturae Sinica, 2009, 36(5): 749-754. (in Chinese)
[27] 邓祖湖. 甘蔗与斑茅杂交的染色体遗传及育性相关基因的筛选[D]. 福州: 福建农林大学, 2010.
DENG Z H. The chromosome inheritance for the hybrid progeny ofL. andand the screening of fecundity-related genes[D]. Fuzhou: Fujian Agricultural and Forestry University, 2010. (in Chinese)
Identification and Genetic Analysis of F1Hybrids from WildL. of Sugarcane Using SSR Markers
TIAN Chunyan1, BIAN Xin1, DONG Lihua1, WU Caiwen2, LANG Rongbin1, YU Huaxian1, ZHANG Yu1, TAO Lianan1, JING Yanfen1*
1. Ruili Breeding Station, Sugarcane Research Institute, Yunnan Academy of Agricultural Sciences, Ruili, Yunnan 678600, China; 2. Sugarcane Research Institute, Yunnan Academy of Agricultural Sciences / Yunnan Key Laboratory of Sugarcane Genetic Improvement, Kaiyuan, Yunnan 661699, China
is the most widely used and the most successful wild species in sugarcane breeding due to its good ratoon ability, strong stress resistance and wide adaptability. It has a great significance for sugarcane cultivars improvement by further exploring and using excellent stress resistance genes resources of widein modern sugarcane breeding programs. To obtain more true hybrids of, three segregation populations of three hundred and fifty-nine F1hybridswere developed from the inter-specific hybridization between two landrace (female) clones and three(male) genotypes in this study. Six SSR primer pairs with polymorphism and clear amplification fragments within parents were selected from twenty-one SSR primer pairs and the high-throughput fluorescence-capillary electrophoresis detection platform were used for SSR genotyping. Subsequently, hybrid identification, SSR fingerprints analysis of five parents, genetic similarity analysis within each two parents, and genetic analysis of parental-specific fragments were performed. The results showed that 6 SSR markers screened out in this study had high resolution and good amplification polymorphism within 5 parents. The map of five parents indicated that each parent had a unique SSR fingerprint, which could effectively identify the blood authenticity of their hybrid offspring. 262 of 359 progenies from three crosses were identified to be true hybrids, and the percentage of true hybrids were 67.77%, 75.51% and 75.66%, respectively, with a mean of 72.98%. Pairwise genetic similarity analysis showed that the coefficients of genetic similarity (SC) between two landraces was 0.70. The SC value of each twogenotypes were 0.53, 0.60 and 0.70, respectively. Whereas, the SC value between landrace andgenotypes ranged from 0.38 to 0.53, suggesting a smaller SC value was existed in inter-species than intra-species. Genetic analysis of SSR fragments revealed that heritability of female specific fragments from three crosses were 68.47%, 80.96% and 73.39%, respectively, with an average of 74.27%. While the heritability of male specific fragments was 58.90%, 76.60% and 61.45%, respectively, with a mean of 65.65%. Collectively, F1generations from inter-specific hybridization between landrace andgenotypes had the genetic tendency to female parent. Therefore, the clones with good comprehensive traits should be used as female preferentially in sugarcane crossing program. The results could provide effective and reliable SSR markers for hybrid identification ofprogenySimultaneously, the true hybrid populations identified of this study could offer good germplasm resources for genetic research of good agronomic traits and provide scientific basis for superparental clones selection.
sugarcane;n; SSR; hybrid identification; genetic
S566.1
A
10.3969/j.issn.1000-2561.2022.10.007
2021-10-27;
2022-03-15
国家自然科学基金项目(No. 31860406);国家重点研发计划项目子课题(No. 2018YFD1000503);云南省基础研究面上项目(No. 2019FB053)。
田春艳(1989—),女,硕士,助理研究员,研究方向:甘蔗分子育种。*通信作者(Corresponding author):经艳芬(JING Yanfen),E-mail:jyf@yaas.org.cn。
猜你喜欢
计算机应用与软件(2022年6期)2022-07-12
油气地质与采收率(2022年3期)2022-05-20
自然灾害学报(2022年2期)2022-05-10
中国饲料(2022年5期)2022-04-26
中国学校体育(2021年10期)2021-04-26
散文诗世界(2019年10期)2019-09-10
农业科技与装备(2017年9期)2018-03-02
江苏农业科学(2015年5期)2015-10-20
科技资讯(2014年26期)2014-12-03
现代农业科技(2009年1期)2009-04-09
