
川西亚高山林线岷江冷杉和红杉对气候变化的响应
2022-12-26 13:25薛盼盼张远东
生态学报 2022年23期
薛盼盼,缪 宁,*,王 东,张远东
1 四川大学生命科学学院,教育部生物资源和生态环境重点实验室, 成都 610065 2 中国林业科学研究院森林生态环境与自然保护研究所,国家林业和草原局森林生态环境重点实验室, 北京 100091
以气候变暖为主要特征的全球变化影响着森林的结构、功能和稳定性[1—2]。树木作为森林生态系统的基本结构单元,水热条件的改变将影响树木径向生长对气候因子的响应。由于气候变化的复杂性以及树木径向生长与环境因子之间耦合关联的关系,树木径向生长对气候因子的敏感性变化极其复杂[3],这使得气候变化对树木生长的影响以及树木的响应方面仍然存在很多不确定性。因此,只有综合评估树木径向生长对气候变化的响应,了解树木径向生长对气候变化响应的现状和规律,才能更好的应对气候变化给森林可持续经营所带来的挑战。
在林线区域,树木径向生长对气候变化尤为敏感。一般认为,海拔上限树木的生长主要受温度的限制。1995年,Jacoby和D′Arrigo[4]在阿拉斯加林线区域发现,随着温度的升高,白云杉(Piceaglauca)年轮宽度和密度对温度的敏感性趋于下降,径向生长表现出放缓甚至停滞,他们由此提出了树木径向生长与温度变化的“分异现象”[5]。分异现象动摇了树木年轮学基本的假设前提-均一性原理,对基于树木年轮重建历史气候方法体系提出了质疑[6—7]。温度阈值效应[5, 8]、大气CO2浓度的变化[9]、立地微环境的改变[10]和去生长趋势导致的尾端效应[11]等均有可能造成分异现象。然而,气候暖干化导致的干旱胁迫[4, 12—16]被认为是造成该现象的主要原因。
全球变暖导致极端干旱事件发生的频率和强度急剧增加[17],森林衰退事件频现,树木出现生长下降、停滞甚至死亡[18]。树木年轮可以记录树木径向生长滞缓或下降的过程,是研究干旱事件对树木径向生长影响的理想资料。判定树木生长衰退一般是通过计算干扰事件前后若干年的年轮指数平均值的比率,生长变化率持续处于负值表明树木的径向生长受到抑制,而长时间的抑制则会引发衰退事件[19]。Williams等[20]利用覆盖美国西南部335个采样点的数据建立了区域树木年轮干旱胁迫指数序列,发现该指数序列不仅很好的记录了发生在2000—2012年、1945—1964年和1899—1904年有文献记载的森林衰退事件,而且揭示了过去1000年无文字记录的森林衰退事件。此外,树木即使能够在极端干旱事件中存活,对干旱事件的敏感性也会发生变化[21]。Gao等[22]发现,树木生长对不同频度的干旱存在响应差异,干旱频率的增加和持续时间的延长,会导致树木抵抗干旱的能力下降。
20世纪90年代以来,青藏高原温度显著升高,而降水的变化则并不明显[23],气候趋于暖干化。气候暖干化通常被认为是导致干旱和半干旱地区树木生长下降的主要原因,因为这些地区的森林特别容易受到干旱胁迫的影响,但气候暖干化对半湿润地区森林生长的影响研究较为鲜见[24]。青藏高原东缘的川西地区,地形起伏大,地貌复杂,相对高差悬殊[25],其丰富的高山森林资源使得该地区成为国内年轮生态学研究的热点地区。然而,川西地区以往研究侧重探索不同树种、不同海拔树木径向生长对气候变化的响应差异[15,26—29],以及树木径向生长与增温的分异现象[30—32]。例如,张萌等[29]发现温度升高会促进川西道孚林线鳞皮冷杉(Abiessquamata)、红杉(Larixpotaninii)和川西云杉(Picealikiangensisvar.rubescens)的径向生长,但方枝柏(Juniperussaltuaria)的径向生长受到的影响相对较小。彭钟通等[33]通过比较川西道孚林线川西云杉和鳞皮冷杉径向生长在升温前后对气候因子的响应差异发现,升温突变后林线川西云杉径向生长依然受夏季温度控制,而鳞皮冷杉径向生长则受到春季干旱胁迫的影响。然而,在川西地区,对林线处生物学特性差异明显的树种,综合分异现象、衰退问题和生态弹性的评价研究仍然较少。
本研究选取川西林线优势树种岷江冷杉 (Abiesfargesiivar.faxoniana)和红杉为研究对象。岷江冷杉耐阴性强,喜冷湿,红杉则耐干寒,喜光照。在区域暖干化加剧背景下,生长环境相对干旱瘠薄的红杉是否面临更加严峻的生长问题。为了验证这一猜想,本文拟通过回答以下4个科学问题:(1)限制林线岷江冷杉和红杉的径向生长的主要气候因子是什么?(2)岷江冷杉和红杉与年均温的升高之间是否存在分异现象?(3)岷江冷杉和红杉是否存在较为明显的生长衰退,两树种有何差异?(4)岷江冷杉和红杉应对严重干旱事件的抵抗力和恢复力在时间尺度上有何变化?以期为预测该区域森林生态系统对气候变化的响应及适应提供依据,为林线树木对气候变化的响应和历史气候重建提供理论参考。
1 研究方法
1.1 研究区概况
研究区位于四川省阿坝羌族藏族自治州,属青藏高原东缘褶皱带外缘部分,地形以石质山地为主,地貌为典型的高山峡谷。该区域植被垂直地带性特征明显,海拔3000 m以上主要为亚高山暗针叶林[34]。岷江冷杉是川西亚高山暗针叶林主要建群树种之一,在高海拔区域常形成大面积的天然纯林[35]。红杉常在云、冷杉林上部边缘形成不连续的淡绿色落叶针叶林带,海拔分布在纬度上自南向北呈递减趋势,最高分布上限达4300 m,个体分布在康定大炮山有4800 m的分布记录[36]。
1.2 气象数据
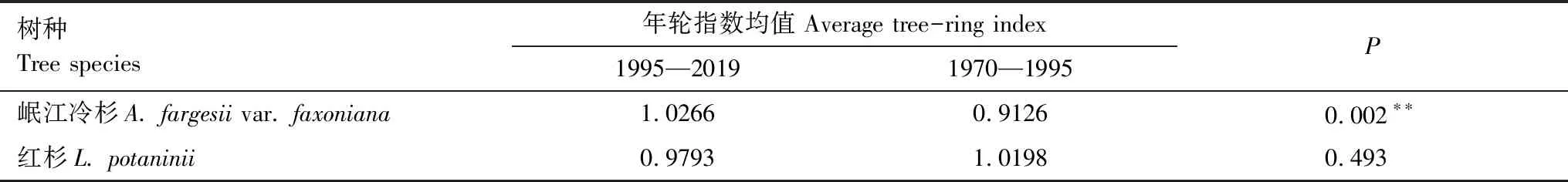
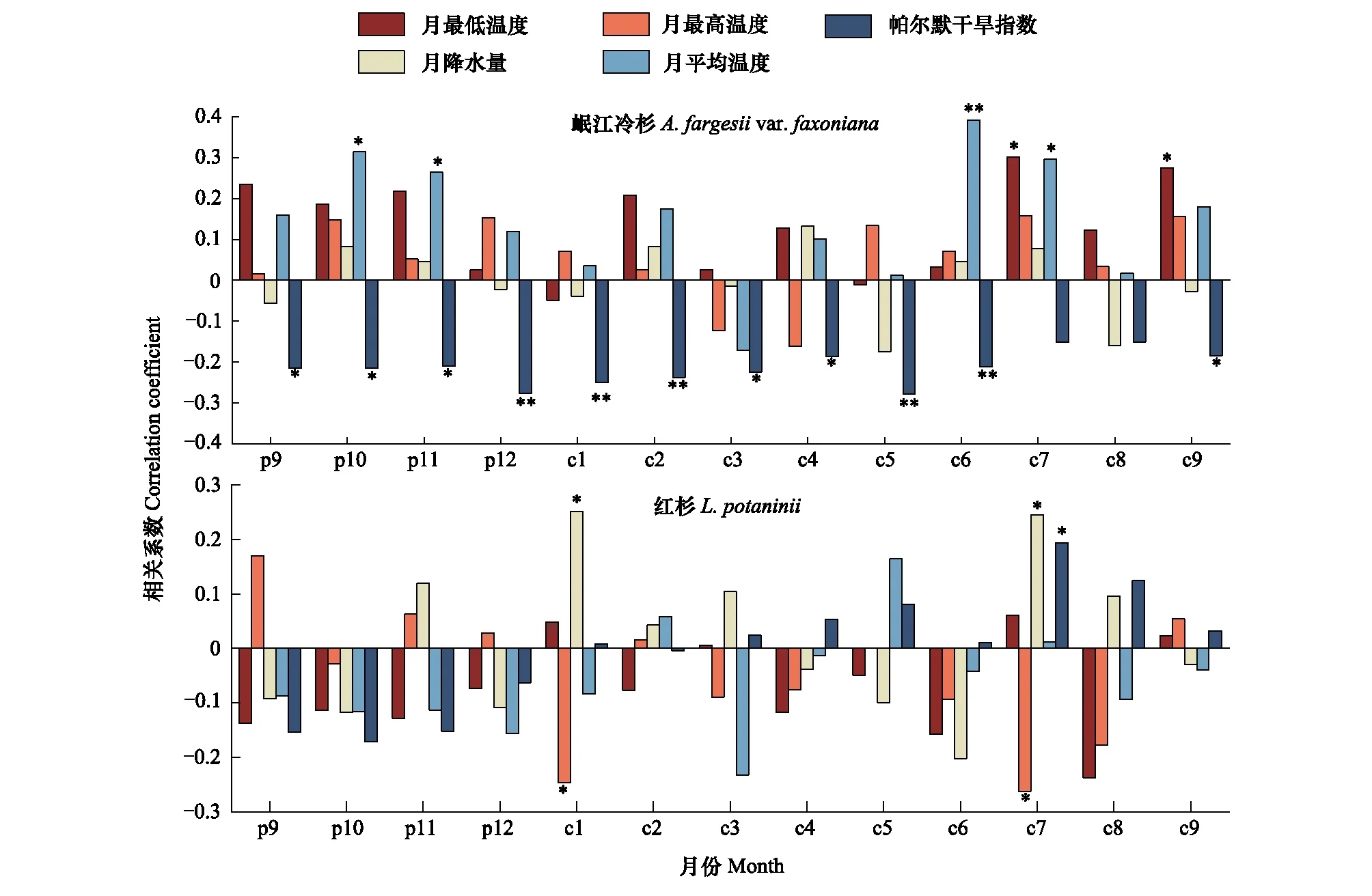
从中国气象数据网(http://data.cma.cn)获取距离岷江冷杉采样点25 km的国家标准气象站-马尔康气象站(31°54′N,102°14′E,海拔2664 m)的气候数据,使用该站1954—2019年的年平均温度(Ty)、月平均温度(Tm)、月最低温度(Tmin)、月最高温度(Tmax)和月总降水量(Pm)等5项气候数据与岷江冷杉和红杉标准年表年轮指数进行分析。帕尔默干旱指数(Palmer drought severity index)采用世界气象组织网站(World Meteorological Organization,https://climexp.knmi.nl/)的区域平均自校准数据(区域范围为102°—103°E,31.5°—32°N),数据时段为1901—2019年。帕尔默干旱指数(PDSI)用于衡量大气水分供应和土壤水分条件的累积效应,是考虑了温度、降水和蒸发的综合水分条件指标。帕尔默干旱指数的评定标准:-0.5 青藏高原在20世纪80年代呈现出变暖的趋势,1995年左右发生增温突变[23],郭明明等[30]在研究升温对马尔康地区树木的影响时,综合分析后认为1995年为该地区增温的突变年份,因此本研究将1995年作为增温突变年。 图1 月平均帕尔默干旱指数所指示的严重干旱年份Fig.1 Severe dry years as indicated by the monthly mean Palmer Drought Index 2020年7月采集了岷江冷杉和红杉年轮树芯。采样时,选取生境相似且生长状况良好的大径级树木(DBH ≥ 25 cm),垂直于胸径(约1.3 m)处沿等高线方向用生长锥钻取树芯(个别生长位置特殊的树木,限于采样条件,钻取高度和方向有所差别)。每株树木采集1个树芯,共采集95个树芯,其中,岷江冷杉45个,红杉50个(表1)。 表1 树芯采样信息 将采集的树芯样品带回实验室,待其自然风干后固定于U型木槽,先后采用300目、800目和1200目砂纸进行打磨和抛光,直至年轮清晰可见。在显微镜(型号OLYMPUS SZ51)下初步标记伪年轮,使用万深LA-S植物图像分析仪测量年轮宽度(精度0.001 mm),利用COFECHA程序对交叉定年的结果进行检验。鉴于林线区域树木稀疏,树木之间的相互作用微弱,主要受气候因素的影响,因此通过ARSTAN程序中的负指数函数消除树木生长趋势,以保留更多的气候信号,建立岷江冷杉和红杉的标准年表(STD)。 分析岷江冷杉和红杉标准年表序列与温度变化趋势的一致性[31,37]。分别将年平均温度及标准年表序列进行曲线拟合,判断拟合曲线在1995年后变化趋势。此外,以1995年为温度变暖的分界点,使用配对样本t检验的方法,分别检验岷江冷杉和红杉1970—1994年与1995—2019年岷江冷杉和红杉年轮指数的差异显著性。 采用树木径向生长变化百分率(Growth change,GC(%))分析树木生长衰退,即树木发生生长衰退前后若干年树木年轮指数的变化率[38]。为了降低个别特殊年份年轮指数对计算对果的影响,采用5年滑动平均计算岷江冷杉和红杉树木径向生长变化率。树木径向生长变化率的计算公式为: GC=(Mi-5-Mi+5)/Mi+5×100% 式中,Mi-5和Mi+5分别为第i年的前5年(包括第i年)和后5年的年轮指数的平均值。若GC为负值,说明树木径向生长受到抑制,若GC小于-25%,则判定树木生长发生衰退。 依据Lloret等[39]提出的抵抗力和恢复力公式公式计算岷江冷杉和红杉应对严重干旱事件的抵抗力(Resistance)和恢复力(Recovery)指标: Rt=InGr/PreGr 式中,Rt和Rc分别表示树木的抵抗力和恢复力。InGr指发生严重干旱事件当年的树木年轮指数值,PreGr和PostGr分别表示干旱事件前后4年的平均年轮指数值。若Rt>0.75,则表明对严重干旱事件具有高抵抗力,若Rc>1.25,则表明在严重干旱事件之后具有高恢复力。 考虑到树木径向生长对气候变化的反馈存在滞后效应,本研究选取前一年9月到当年9月的气候数据与岷江冷杉和红杉标准年表序列进行皮尔逊(Pearson)相关分析。以上统计分析均采用SPSS 25.0软件进行,显著性水平α=0.05,制图使用Origin 2018软件完成。 由岷江冷杉和红杉的标准年表可以看出,20世纪以来,岷江冷杉的年轮指数在1900年后上升,50年代末趋于下降,70年代中期达到最低,1980年后持续增长。然而,红杉的年轮指数变化比较稳定,仅在1995后呈现出下降的趋势(图2) 图2 岷江冷杉和红杉的标准年表Fig.2 Standard chronology of A. fargesii var. faxoniana and L. potaninii 岷江冷杉和红杉的样本总体代表性分别为0.92和0.90,大于树木年轮气候分析0.85的阈值要求。第一特征方差解释量达到36.06%以上,信噪比分别为11.96和9.30,平均敏感度分别为0.13和0.20(表2),这说明岷江冷杉和红杉标准年表包含了大量的气候信息,符合年轮气候学的分析要求。 表2 标准年表的主要统计特征 20世纪90年代中期,岷江冷杉年轮指数变化趋势相对稳定,呈不断增长的趋势;红杉年轮指数呈现下降的趋势,与年平均温度的增长趋势表现出分异现象(图3)。 图3 1954—2019年平均温度和岷江冷杉与红杉的年轮指数之间的变化趋势Fig.3 Trends of annual mean temperature and tree-ring index of A. fargesii var. faxoniana and L. potaninii from 1954 to 2019 岷江冷杉1995—2019年的年轮指数极显著大于1970—1994年间的年轮指数(P< 0.01),而红杉1995—2019年间的年轮指数与其1970—1994年间的年轮指数之间无显著差异(表3)。 树木径向生长变化百分率小于25%,则表明这些年份及之后5年内树木生长发生了衰退。即,岷江冷杉在1811—1818年间和1835—1841年间,红杉在1661—1668年间、1711—1720年间、1749—1754年间、1783—1789年间、1812—1818年间、1836—1845年间、1857—1865年间、1903—1910年间和2011—2019年间发生了生长衰退(图4)。90年代温度升高以来,岷江冷杉的生长状况良好,未出现生长衰退。红杉在2011—2019年间出现了连续9年的生长衰退。在1800—2019年间,岷江冷杉出现生长衰退2次,红杉出现生长衰退5次,相同 时间内,红杉发生衰退的频率高于岷江冷杉。 表3 岷江冷杉和红杉年轮在升温前后的对比 图4 岷江冷杉和红杉径向生长变化百分率Fig.4 Percentage of radial growth change of A. fargesii var. faxoniana and L. potaninii 岷江冷杉的恢复力弹性均小于1.25,红杉的恢复力弹性在不同年份的差异较大,在1939—1943年间和1959年大于1.25,在1928—1931年、1987年和2003年则小于1.25;岷江冷杉和红杉的抵抗力均大于0.75,呈现出高抵抗力(图5)。总体来讲,红杉的恢复力高于岷江冷杉,岷江冷杉在1928—1931年间、1939—1943年间和1959年的抵抗力高于红杉,在1987年和2003年的抵抗力低于红杉(图5)。以90年代快速升温为界限的时间尺度来看,岷江冷杉的恢复力有所增强,而抵抗力相比升温前下降明显;红杉抵抗力和恢复力相比升温前均有所减弱(图5)。 图5 岷江冷杉和红杉的在严重干旱年份的抵抗力和恢复力Fig.5 Resistance and recovery of A. fargesii var. faxoniana and L. potaninii in severe drought years 岷江冷杉年轮指数与前一年10月和11月、当年7月平均温度呈显著正相关(P<0.05),与当年6月平均温度呈极显著正相关(P<0.01),与当年7月和9月最低温度呈显著正相关(P<0.05),与前一年9—11月,当年3月、4月、6月和9月帕尔默干旱指数呈显著负相关(P<0.05),与前一年12月,1月、2月和5月帕尔默干旱指数呈极显著负相关(P<0.01)(图6)。红杉与当年1月和7月最高温度呈显著负相关(P<0.05),与当年1月和7月的降水量呈显著正相关(P<0.05),与当年7月的帕尔默干旱指数呈显著正相关(P<0.05)(图6)。岷江冷杉的径向生长与温度表现出更多的正相关,在前一年冬季和生长季相关性均达到了显著;红杉则与生长季前和生长季的高温呈显著负相关,生长季前和生长季降水呈显著正相关。这表明温度是限制林线岷江冷杉径向生长的主要气候因素,而限制红杉径向生长的主要气候因素是降水。 图6 年轮指数与月气候因子的相关性Fig.6 Correlations between tree-ring indices and monthly climate factors*表示显著相关(P<0.05);**表示极显著相关(P<0.01);p表示前一年;c表示当年 岷江冷杉年轮指数与前一年10月和11月的月平均气温呈显著正相关(P<0.05)(图6),该研究结果进一步证实了温度是限制林线树木径向生长的主要因素。前一生长季末适宜的温度有利于光合产物积累,为下一年树木生长创造出充足的物质条件[26]。温暖的冬季则有利于树木完成正常的生理代谢活动,避免活性组织遭受冻伤(形成层的初始细胞、叶组织等),保证来年树木生长的潜力[40—41]。当冬季温度偏低时,植物叶细胞组织可能会脱水,土壤长时间冻结可能会导致植物根系部分组织死亡,造成来年树木光合作用减弱和生长季缩短[42]。岷江冷杉的径向生长与当年7月平均温度和7月最低温度呈显著正相关(P<0.05),与当年6月平均温度呈极显著正相关(P<0.01)(图6)。岷江冷杉径向生长与生长季温度呈显著正相关的结果与李宗善等[43]在川西卧龙,郭滨德等[44]在川西九寨沟,以及赵志江等[31]在王朗自然保护区的研究结论一致。在川西林线区域,6月和7月是岷江冷杉的主要生长季,温度的升高可以促进树木的光合作用,进而可以为树木提供充足的养分,从而有利于其生长[45]。岷江冷杉的径向生长与9月最低温度呈显著正相关(P<0.05)(图6),9月末已接近其生长季末期,生长季末最低温度的升高会改变植物物候,延长生长季的时间[46]。 与温度升高促进岷江冷杉径向生长不同的是,红杉的径向生长则受到增温的抑制作用。树木的径向生长与温度的显著负相关,月降水量的正相关,则认为树木径向生长受干旱胁迫的影响[47—48]。本研究中红杉的径向生长受1月和7月干旱胁迫的不利影响,说明其径向生长受春季和生长季降水的限制。7月光照辐射强烈,温度的升高会加剧林线环境下土壤水分蒸发和植被的蒸腾作用,造成树木的水分供应不足,进而会抑制树木的径向生长[49]。 干旱胁迫造成树木径向生长与增温发生分异这一现象,广泛的分布于北半球的高纬度、高海拔区域,以气候寒冷,区域气候暖干化趋势明显的区域最为突出[6]。例如,Lloyd等[50]对阿拉斯加地区的8个林线样点研究发现,除湿润地区外,其余样点的树木径向生长在1950年均出现了下降,从而推断升温引起的干旱胁迫是导致树木生长分异的主要原因。在本研究中,年平均温度与岷江冷杉和红杉的年轮指数间共变趋势的分析表明:岷江冷杉年轮指数在20世纪90年代中期之后随年平均温度的逐渐升高呈增长趋势,但红杉年轮指数与年平均温度之间未表现出共变趋势,其径向生长与增温表现出相反趋势(图2)。此外,岷江冷杉1995—2019年与1954—1994年标准年表年轮指数的比较中,岷江冷杉的径向生长显著增大(表3)。高海拔树木径向生长随温度的升高而加快的结论在青藏高原的一些区域上也得到了印证[24,51—53]。结合红杉年轮指数与月气候因子的相关性分析结果,红杉与年均温的分异现象可能是由于干旱胁迫抑制了其径向生长。干旱胁迫会通过降低细胞分裂速率从而限制树木生长,通过减少光合作用和可用于次生生长的可溶性糖而间接限制树木生长[54]。随着水分有效性减少,缺水引起的干旱胁迫会导致树木径向生长提前停止,细胞大小和管腔直径减小,细胞产量降低[55],从而导致窄轮的出现。 岷江冷杉和红杉的衰退事件可以从径向生长的历史变化中判定。岷江冷杉在1811—1818年间和1835—1841年间,红杉在1812—1818年间和1836—1845年间的发生了2次生长衰退(图3)。喻树龙等[56]重建了川西马尔康7月平均气温,在1787—1850年间,马尔康地区的7月平均气温明显偏低。李宗善等[25]利用川西林线区域高山柏(Sabinasquamata)年轮资料重建过去200年的夏季温度,表明1810—1850年之间为该区的极寒冷时期。青藏高原东部普若岗日冰芯亦表明了1810—1850年间为极寒冷时期[57]。本研究中,两个树种的衰退发生时期与以往气候重建的寒冷时期是一致的,其生长衰退应都归因于该寒冷时期的低温。除了共同的衰退年份时期,红杉在1661—1668、1711—1720、1749—1754、1783—1789、1857—1865、1903—1910和2011—2019等年份间亦发生了生长衰退(图3)。马尔康地区7月份气候重建研究结果发现在1624—1675、1705—1757、1773—1786、1851—1868、1893—1919和2011—2014等年份间,气候明显偏暖[56],该重建结果的偏暖时间与本研究中红杉发生生长衰退的时间基本吻合。结合气候因子与红杉年轮指数相关性分析的结果,林线区域发生的干旱胁迫可能是红杉生长衰退发生的主要原因。 川西马尔康地区的严重干旱年份为1928—1932、1939—1943、1959、1987和2003年,这些年份中岷江冷杉和红杉的年轮指数均保持相对稳定,且两个树种的抵抗力均大于0.75(图5),这表明其径向生长并未受到严重干旱事件干扰。随着20世纪90年代的持续增温,岷江冷杉的恢复力有所增强,而抵抗力相比升温前下降明显;红杉的抵抗力和恢复力相比升温前均呈下降的趋势(图5)。升温促进林线了岷江冷杉径向生长,但加速生长引起的过度消耗可能是导致岷江冷杉抵抗力下降的主要原因,而升温带来的适宜的生境可能使得岷江冷杉恢复力趋于增强。 红杉的抵抗力和恢复力相比升温前均呈下降趋势,这可能是由于升温使得位于冷杉林线之上红杉的生境愈加干燥,其抵抗严重干旱事件及在严重干旱事件后的恢复力趋于下降。Longo等[58]研究发现,较干燥地区的森林对极端干旱的抵抗力较低。红杉在1928—1932、1939—1943和1959等年份的抵抗力均低于岷江冷杉的结果与Longo等的结论一致,但不同的观点则认为长期生长在干旱环境中的树木具备维持树木生理生长的适应能力,相对比湿润地区的树木,其抗旱能力更强[59]。在1987年和2003年,岷江冷杉的抵抗力低于同时期红杉的抵抗力。可能有两方面原因,一方面,岷江冷杉快速生长可能会带来生理消耗;另一方面,红杉喜光、耐干寒的生物学特性可能使其抗逆性更强,这也可能是红杉的恢复力在严重干旱事件中均优于岷江冷杉的原因。 林线岷江冷杉的径向生长主要受温度条件限制,红杉径向生长的主要限制因素是降水。近几十年的升温明显的促进了岷江冷杉的径向生长,使得岷江冷杉应对严重干旱事件的恢复力有所增强,而抵抗力相比升温前下降明显。与之相反,红杉的径向生长则呈现出下降的趋势,与年均温的升高表现出明显的分异,对严重干旱事件的抵抗力和恢复力相比升温前均呈现出下降的趋势。从长时间尺度来看,1800—2019年间岷江冷杉出现生长衰退2次,红杉出现生长衰退5次,相同的时间内,红杉的生长衰退频率高于岷江冷杉。综上所述,在未来气候变化的背景下,温度适度的升高可能会促进林线处岷江冷杉的径向生长,但会给红杉的径向生长带来更多的不利因素。本研究从气候对树木径向生长影响的角度推断了未来增温可能会对林线岷江冷杉和红杉带来的影响,但未考虑到气候变化的复杂性及树木的适应机制等因素可能带来的影响,因此,大范围和多角度的探索其背后的响应机制有待深入研究。
1.3 树芯采集与定年

1.4 数据分析方法
Rc=PostGr/lnGr2 结果与分析
2.1 标准年表统计特征


2.2 年轮指数与温度变化的分异

2.3 岷江冷杉和红杉的生长衰退

2.4 岷江冷杉和红杉的应对严重干旱事件抵抗力和恢复力差异

2.5 年轮指数与气候因子的相关关系
3 讨论
3.1 影响林线岷江冷杉和红杉径向生长的主要气候因子
3.2 岷江冷杉和红杉的生长分异现象
3.3 岷江冷杉和红杉的生长衰退
3.4 岷江冷杉和红杉应对干旱的抵抗力和恢复力差异
4 结论
猜你喜欢
生物化工(2021年3期)2021-07-10
农业工程学报(2020年3期)2020-04-09
科学导报(2019年73期)2019-12-20
花火A(2019年1期)2019-03-22
语文世界(小学版)(2018年9期)2018-09-18
灾害学(2018年2期)2018-04-12
科学之谜(2018年3期)2018-04-09
现代养生·上半月(2017年7期)2017-07-07
振动与冲击(2014年16期)2014-09-08
中南林业科技大学学报(2012年9期)2012-12-29
