
荔波樟江野生斑鳜线粒体Cyt b基因序列多态性分析
2023-02-02 13:06麻智芳潘秋芝何孝宽
水产科学 2023年1期
麻智芳,余 科,安 苗,黄 胜,潘秋芝,李 珊,何孝宽
( 1.贵州大学 动物科学学院,贵州 贵阳550025; 2.贵州农业职业学院,贵州 贵阳 551499; 3.荔波县农业农村局,贵州 黔南 558400 )

樟江系珠江流域西江水系柳江支流龙江的上游河段,是贵州省荔波县境内最大的河流。樟江流域地处云贵高原东南过渡的斜面上,喀斯特地貌特征明显,且地处亚热带季风湿润气候区,其径流的季节变化和年际变化都比较大。其特殊的地貌和适宜的气候造成了该流域复杂多样的生态环境,为鱼类的繁衍分化提供了优渥的自然条件。自20世纪80年代至今,野生斑鳜一直是樟江流域的主要经济鱼类,但是产量不高[7-9]。在我国野生鳜类资源下降的大趋势下[10],了解该水域野生斑鳜的遗传变异特征及其多样性水平,对开展野生斑鳜资源保护和开发利用具有重要意义。
当前,关于斑鳜的研究涉及形态学、个体生态学、繁殖生物学特性以及营养与饲料学等领域[11-16],但主要集中在斑鳜的分子系统发育、遗传多样性和种群结构等方面[17-20]。遗传多样性研究是评估物种资源现状、制定物种保护和管理策略的重要前提。王伟伟等[21-23]认为,我国斑鳜的不同地理群体大体能划分成以秦岭山脉为界的南北两个群体;但Cao等[19]研究发现,鸭绿江斑鳜群体和长江中上游的斑鳜群体具有较高的亲缘性,不能简单地按照地理屏障分为南北群体;而Chu等[24]则将斑鳜分成珠江群体和长江群体。贵州省境内河流被苗岭山脉划为珠江水系和长江水系,丰富的水资源和发达的喀斯特地貌孕育了种类多样的鱼类资源。贵州境内长江流域中一些支流的鳜类遗传多样性研究已有报道[6,20,25],而贵州境内珠江流域中柳江水系的一些主要支流的鳜类遗传学评估尚未见报道。为填补贵州境内珠江水域柳江水系上游斑鳜野生遗传多样性研究的空白,合理开发和利用野生斑鳜资源,并为保护野生斑鳜种质资源提供科学依据,笔者采用Cyt b基因作为分子标记,对柳江水系上游贵州省荔波县境内的樟江野生斑鳜的遗传多样性进行初步研究。
1 材料与方法
1.1 材料
2015年10月至2019年9月间,在贵州省樟江荔波县河段的捞村乡、小七孔景区和水春河景区3个点采集野生斑鳜共75尾(表1)。每尾样本取背部肌肉组织3~5 g放入离心管中,加入无水乙醇固定,-20 ℃保存备用。

表1 荔波野生斑鳜采样信息
1.2 方法
1.2.1 DNA提取
采用(肌肉、血液、组织)基因组DNA提取试剂盒(北京天根)提取野生斑鳜基因组DNA。用微量紫外分光光度计检测其浓度及吸光值,1.0%琼脂糖凝胶(1×TBE)电泳检验其完整性,将成功提取的DNA置于-20 ℃保存。
1.2.2 PCR扩增及测序
扩增野生斑鳜线粒体Cyt b基因序列的引物参考文献[18](表2),由生工生物工程(上海)股份有限公司合成。PCR反应总体积为25 μL,其中模板DNA 2 μL,2×Taq PCR Master Mix 12.5 μL,正、反引物各1 μL,用超纯水补足体积。在PTC-200PCR扩增仪(AB 公司)进行扩增,PCR反应程序为:94 ℃预变性5 min;94 ℃变性40 s,56.5 ℃退火50 s,72 ℃延伸50 s ,重复35个循环;再72 ℃延伸10 min,反应终产物4 ℃保存。
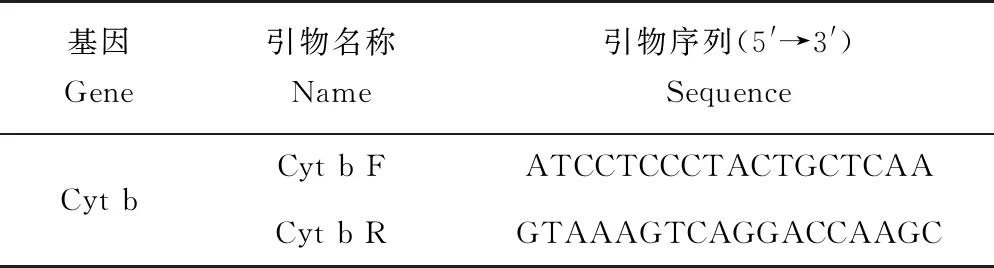
表2 引物名称及序列
PCR产物经1.0%琼脂糖凝胶(1×TBE)电泳检测,将目的条带清晰的产物送至生工生物工程(上海)股份有限公司纯化并双向测序,测序引物为扩增引物。
1.3 数据处理
利用DNAstar软件[26]对测序结果进行序列编辑、比对和排序,并进行人工校对。采用MEGA 6.0软件计算碱基组成、氨基酸含量、遗传距离,并以花鲈(Lateolabraxmaculatus)为外群,运用邻接法构建单倍型分子系统进化树。使用DnaSP 5.0 软件[27]计算单倍型数目、多态位点数目、单倍型多样性、核苷酸多样性、变异位点、单一突变位点、简约信息位点等。利用Tajima′sD中性检验和核苷酸不配对分析斑鳜种群历史上是否发生过扩张。利用软件Network 10.2按照荔波野生斑鳜单倍型频率绘制神经网络图,对单倍型间的进化关系进行分析。
2 结果与分析
2.1 荔波野生斑鳜Cyt b基因碱基组成和变异分析
经校对和拼接,得到75条长度为1141 bp的Cyt b基因同源序列。序列中的碱基A、T、G和C的平均含量分别为24.4%、27.2%、15.5%和 32.9%,A+T的含量(51.6%)略高于G+C(48.4%)的含量,呈碱基偏倚性(表3)。75条Cyt b基因序列中,共检测到1124个保守位点和17个变异位点,变异位点是分析位点总数的1.40%。其中:有8个为简约位点,分别位于147、228、294、363、711、798、897和975位点;9个为单一多态位点,分别位于42、120、189、321、519、687、795、828和1078位点。这些突变位点中有14个转换位点,分别位于120、189、228、294、321、363、519、687、711、795、798、897、975和1078位点;有3个颠换位点,分别位于42、147和828位点;未有插入和缺失位点(表4)。转换/颠换比为5.34。

表3 荔波野生斑鳜群体线粒体DNA Cyt b基因单倍型碱基组成

表4 荔波野生斑鳜群体线粒体DNA Cyt b基因单倍型多态位点分布
2.2 荔波野生斑鳜种群的遗传多样性
75尾荔波野生斑鳜个体共定义了15个单倍型(Hap1~Hap15),其中单倍型Hap15的个体数量最多,共46尾。荔波野生斑鳜群体的单倍体多样性指数和核苷酸多样性指数分别为0.613±0.064和0.00172±0.00021,平均核苷酸差异数为1.968,中性检验Tajima′sD值为-1.37917,且差异不显著(P>0.10)(表5)。另外整体核苷酸不配对分析结果呈现双峰趋势(图1)。

表5 荔波野生斑鳜的遗传多样性参数
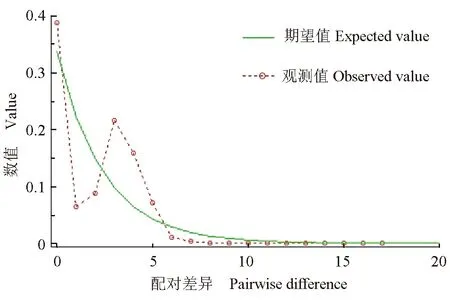
图1 荔波野生斑鳜线粒体DNA Cyt b序列核苷酸不配对分析
2.3 荔波野生斑鳜Cyt b单倍型聚类分析与简约网络图
15个单倍型经美国国家生物技术信息中心数据库中BLAST比对后发现:Hap15与珠江流域中西江水系的支流桂江斑鳜(登录号:DQ648645)、长江流域洞庭湖水系湘江斑鳜(登录号:JQ010988)和沅江斑鳜(登录号:DQ274043)的Cyt b序列相似性为100%,说明该单倍型跨水系分布;Hap9、Hap13与长江流域洞庭湖水系的支流澧水斑鳜(登录号:KC441451)和沅江斑鳜(登录号:KC441448)的Cyt b序列相似性为100%;Hap10与长江中游右岸的支流陆水河斑鳜(登录号:KC441435)和洞庭湖斑鳜(登录号:JQ010986)的Cyt b序列相似性为100%;其他的单倍型与长江流域洞庭湖水系的斑鳜的Cyt b 序列相似性均超过99%。总的来说,荔波野生斑鳜群体的Cyt b与长江流域中上游斑鳜群体具有更高的相似性。从GenBank下载16条鳜亚科的线粒体Cyt b基因同源序列(表6),以花鲈(Lateolabraxjaponicus)(登录号:DQ351530)为外群,与本研究中的15种斑鳜单倍型一起,采用邻接法构建分子系统发育树(图2)。由图2可见,斑鳜Cyt b单倍型与麻鳜(S.fortis)、波纹鳜(S.undulata)、暗鳜(S.obscura)、大眼鳜(S.knerigarman)及鳜(S.chuatsi)聚为一支,中国少鳞鳜(Coreopercawhiteheadi)则单独聚为一支。其中荔波野生斑鳜的15个单倍型和长江流域中的洞庭湖水系及上游的支流陆水河斑鳜群体以100%的置信度单独成支,与珠江流域中西江水系的斑鳜群体存在较大程度的分化。本研究中的15个荔波野生斑鳜Cyt b单倍型间的遗传距离较小,最大值为0.006,最小值为0.001,平均遗传距离为0.003(表7)。
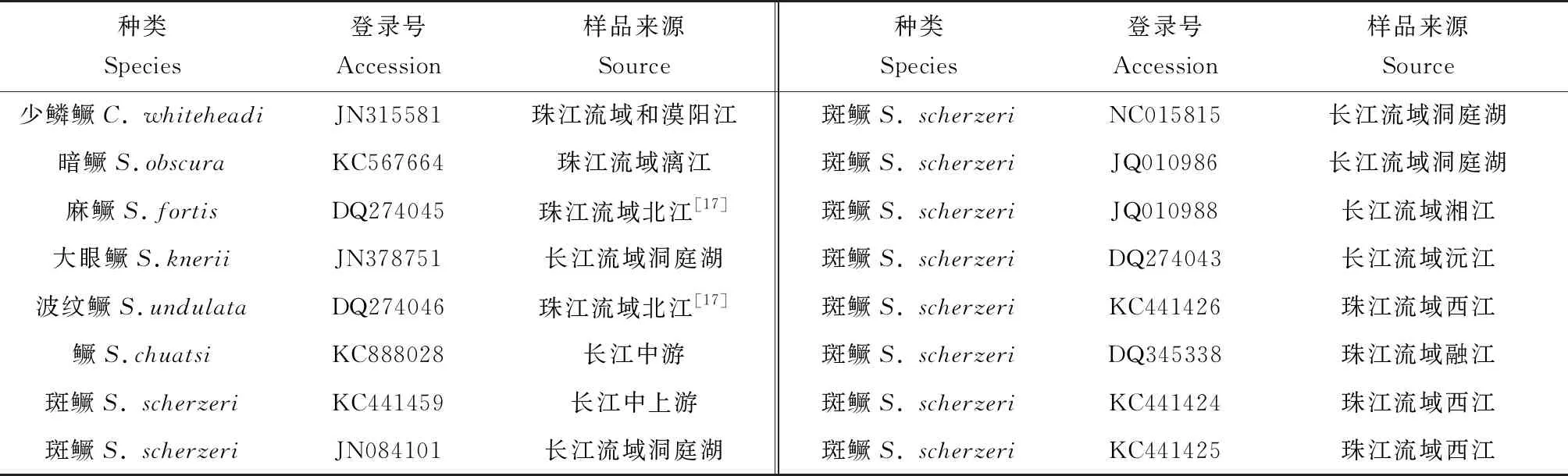
表6 鳜类登录号信息
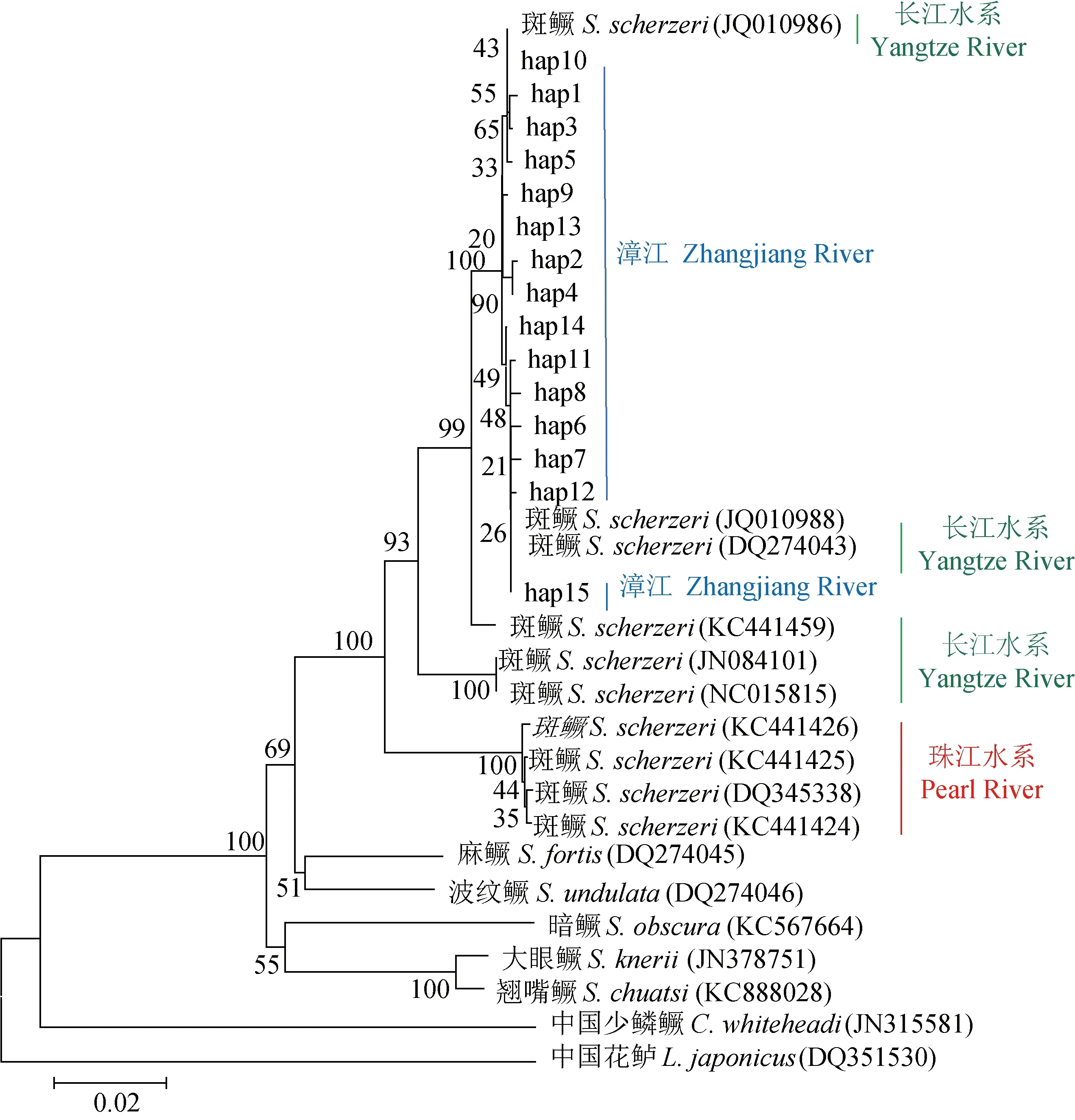
图2 基于Cyt b基因片段荔波野生斑鳜单倍型的邻接法系统发生树

表7 荔波野生斑鳜Cyt b基因序列15个单倍间及鳜类间的型遗传距离(Kimura-2-parameter法)
由基于荔波野生斑鳜单倍型频率绘制出的神经网络图(图3)可以看出,Hap10和Hap3位于网络的中心区域,其他单倍型均由其衍生而来,其在进化出Hap1的连接节点单倍型未检测到或已经灭绝。中心区域进化出Hap5、Hap13,Hap13又发展出Hap4、Hap9、Hap14,再由Hap14进化出荔波的优势单倍型Hap15。

图3 基于NETWORK软件的荔波野生斑鳜单倍型简约网络
3 讨 论
3.1 荔波野生斑鳜群体Cyt b基因片段的基因组成
线粒体DNA为母系遗传,且具有基因保守、序列较小和进化速率快等特点,因此被广泛应用于鱼类系统发育、种群遗传、种类鉴定和适应性进化研究中[28]。线粒体DNA中Cyt b基因是其重要的蛋白质编码基因,具有丰富的系统发育信息,进化速率适中,且易于使用通用引物扩增,是研究鱼类系统发育最常用的分子标记[29]。本研究中,获得了75条荔波野生斑鳜Cyt b基因同源序列,其中碱基G(15.5%)的含量最低,具有明显的反G偏倚,符合一般硬骨鱼类Cyt b基因的特征[30]。在这75条Cyt b基因同源序列中共检测到17个变异位点,其中有14个转换位点,3个颠换位点,没有插入或丢失位点,结果符合同种鱼类中线粒体DNA基因转换在数量上远超过颠换的特征[31]。
3.2 荔波野生斑鳜群体遗传多样性分析
遗传多样性通常是物种长期进化的结果,是物种或其群体持续生存并适应不断变化的环境而不断进化的前提[32]。本研究中,荔波斑鳜线粒体DNA Cyt b序列的Tajima′sD值为负,且核苷酸不配对分析图呈双峰,表明荔波野生斑鳜种群比较稳定,未发生过明显的种群扩张史[33]。单倍型多样性指数和核苷酸多样性指数作为衡量一个物种或者种群遗传变异度的重要指标,可以根据其数值的大小来判断所研究的目标群体的遗传多样性的高低。荔波野生斑鳜群体的单倍型多样性指数和核酸多样性指数分别为0.613±0.064和0.00172±0.00021,呈现出高单倍型多样性和低核苷酸多样性的特征,与李珊等[20,34]的研究结果相同。在Grant等[35]的4种单倍型多样性和核苷酸多样性间的组合类型中,第2种“高单倍型多样性、低核苷酸多样性”的遗传多样性类型与本研究结果相吻合,即在种群扩张过程中,种群数量的增加会导致单倍型多样性的提高,但在较短时间内无法积累核苷酸的变异,从而可能产生单倍型多样性较高、核苷酸多样性较低的遗传模式。荔波野生斑鳜群体遗传多样性呈现这种特征的原因推测有两方面:一方面,冰川时期生境的剧变,使斑鳜种群数量急剧下降,遗传多样性降低,即受到了遗传瓶颈效益的影响,斑鳜在“瓶颈效应”之后, 种群经过短期快速扩张,种群数量迅速扩大,有利于积累更多突变[5,36];另一方面,近年来,除过度捕捞和栖息环境恶化的原因外,随着斑鳜人工繁殖技术取得突破,人工繁殖群体的扩大、异地引种及管理不善等可能也是造成野生斑鳜的种质混杂、遗传多样性降低和本地种群的适应性下降的原因[37]。从遗传多样性水平看,荔波县境内的樟江河段野生斑鳜群体与清水江群体(单倍型多样性0.8165,核苷酸多样性0.0021)[20],鸭绿江(单倍型多样性0.735±0.085,核苷酸多样性0.00120±0.00061)、浙江千岛湖(单倍型多样性0.0858±0.063,核苷酸多样性0.00237±0.00007)、长江中游洞庭湖(单倍型多样性0.800±0.044,核苷酸多样性0.0128±0.00052)、长江下游秋浦河(单倍型多样性0.686±0.080,核苷酸多样性0.00076±0.00044)、闽江(单倍型多样性0.773±0.0690,核苷酸多样性0.0010±0.00050)、西江(单倍型多样性0.914±0.079,核苷酸多样性0.00164±0.00065)、北江(单倍型多样性0.111±0.096,核苷酸多样性0.00029±0.00044)、韩江(单倍型多样性0.268±0.113,核苷酸多样性0.00047±0.00037)等群体[34],钱塘江的江山(单倍型多样性0.570±0.078,核苷酸多样性0.0018±0.0003)、乌溪江(单倍型多样性0.668±0.067,核苷酸多样性0.0019±0.0002)群体,瓯江的云和群体(单倍型多样性0.667±0.032,核苷酸多样性0.0043±0.0006)[38]等野生斑鳜群体相比,其多样性处于中等偏下水平。因此,保护漳江野生斑鳜种质资源对于保护中国斑鳜种质资源的基因遗传多样性具有十分重要的意义。
3.3 荔波野生斑鳜群体的遗传分化
本研究中,15个斑鳜单倍型与其他鳜属的麻鳜、波纹鳜、暗鳜、大眼鳜及鳜聚为单独的一支,置信度为99%,少鳞鳜属则以99%的置信度单独成支,符合刘焕章等[39]的将鳜类分为鳜属和少鳞鳜属二个属分类处理。从鳜亚科的系统发育树看,斑鳜群体Cyt b单倍型以99%的置信度最先被分化出来聚在一起,剩下的鳜属聚成两大支,一支是麻鳜、波纹鳜,另一支包括大眼鳜、鳜和暗鳜,本研究结果支持斑鳜是鳜属中最先分化出来的物种的观点[40-41]。从长江群体和珠江群体亲缘关系上来看,樟江虽然是珠江流域西江水系柳江支流龙江的上游河段,但是15个樟江野生斑鳜单倍型与长江中上流的支流陆水江斑鳜群体和洞庭湖水系斑鳜群体表现出更近的亲缘性,历史原因是:在早更新世时期,湘黔桂低山区未分异时[42],这两个区域的祖先斑鳜可能生活在一起,随后因苗岭山脉形成地理屏障后分开,其在不同河流水系虽然进化速度各异,但母系祖先单倍型仍得以保留。从荔波野生斑鳜的单倍型简约网络图可以推测,Hap10和Hap3为樟江流域荔波河段斑鳜群体的祖先单倍型;Hap3和Hap10分别经美国国家生物技术信息中心数据库中BLAST比对发现,与长江流域的支流陆水河(KC441435)及洞庭湖斑鳜(JQ010986)的相似性为99.9%以上,表明荔波樟江野生斑鳜群体与长江流域斑鳜群体有较高的亲缘性,其结果与邻接系统进化树聚类结果相吻合。从单倍型间的遗传距离和邻接系统进化树看,荔波野生斑鳜单倍型间的遗传距离(0.001~0.006)非常低,所有单倍型聚为一支,表明荔波野生斑鳜种群内没有明显的遗传分化。本研究中的Hap15是跨珠江水系和长江水系分布的单倍型,与周文漪等[34,36]的研究中发现的斑鳜线粒体DNA Cyt b单倍型的有跨水系分布的情况相同。值得注意的是,不同的Cyt b单倍型的出现频率有非常大的差异,其中优势单倍型Hap15的比例高达61.33%,而Hap1、Hap3、Hap6、Hap7、Hap8、Hap11、Hap12、Hap13、Hap14等9个单倍型分别仅为1.33%,需要警惕的是,野生斑鳜种群一旦失去这些数量较少的单倍型,将会导致斑鳜的遗传多样性降低。因此,对于荔波野生斑鳜种质资源的保护除了设立种质资源保护区禁捕和改善水域栖息环境外,还应科学预防人为因素可能带来的不利影响。
4 结 论
物种的遗传多样性或变异性越丰富,则表明该物种的进化潜力越大,对环境改变响应的进化能力就越强[43]。笔者基于线粒体DNA Cyt b基因序列首次调查了樟江荔波县河段的野生斑鳜的遗传多样性水平,结果显示,该水域野生斑鳜群体呈现高单倍型多样性和低核苷酸多样性,与国内其他水系的野生斑鳜群体相比,其多样性处于中等偏下水平,且无种群扩张史。此外,对15个单倍型进行美国国家生物技术信息中心数据库中BLAST比对和构建系统发育树的结果表明,虽然樟江是珠江流域西江水系中柳江的支流,但相对于珠江水系,其野生斑鳜群体与长江水系的斑鳜群体拥有更近的亲缘关系。这些都为樟江野生斑鳜种质资源的保护和合理开发利用提供了参考依据。但是,本研究中荔波野生斑鳜进化出Hap1的连接节点单倍型出现缺失,是样本量不够大而未能检测到,还是该单倍型已经灭绝,需进一步调查研究。最后,为维持该流域野生斑鳜群体的可持续发展和提高其遗传多样性水平,建议可以采取设立野生斑鳜种质资源保护区、有选择地人工引种和增殖放流等措施。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
水利水电快报(2022年7期)2022-07-18
肝博士(2022年3期)2022-06-30
Asian Herpetological Research(2021年4期)2021-12-30
公民与法治(2020年18期)2020-10-28
水利规划与设计(2020年1期)2020-05-25
学苑创造·B版(2019年3期)2019-04-24
夜郎文学(2017年2期)2017-09-26
乡村地理(2017年4期)2017-09-18
