
特异性敲除肠道miR-146a 对小鼠肠道菌群的影响
2023-02-22 19:19邢利鹏朱嘉豪胡芳馨陈婷罗君谊孙加节张永亮习欠云
华南农业大学学报 2023年2期
邢利鹏 朱嘉豪 胡芳馨 陈婷 罗君谊 孙加节 张永亮 习欠云


摘要: 【目的】 miR-146a 作为抑炎因子,仍然不清楚其是否参与宿主与微生物间的互作,进而影响肠道稳态,因此本文旨在研究miR-146a 对小鼠肠道菌群的影响。【方法】以肠道miR-146a 特异性敲除小鼠(CKO 鼠)及对照小鼠(Flox 鼠) 为研究对象,利用16S rRNA 高通量测序法检测2 组空肠段的微生物菌群分布。【结果】测序共获得1 134 个用于物种分类的OTUs,包括 37 门、80 纲、161 目、198 科、261 属、117 种的细菌;Flox 组和CKO 组小鼠的空肠微生物中共有46 个相同的OTUs;各组肠道微生物中厚壁菌门Firmicutes、拟杆菌门Bacteroidota、疣微菌门Verrucomicrobiota、变形菌门Proteobacteria 和脱硫杆菌门Desulfobacterota 是优势菌门;2 组肠道微生物群落组成整体相似,但CKO 组梭状芽孢杆菌纲Clostridia 的平均相对丰度高于Flox 组(P=0.067),毛螺菌目Lachnospirales 平均相对丰度显著高于Flox 组(P<0.05),其他层级组成无显著差异。【结论】 miR-146a 敲除可改变宿主肠道梭状芽孢杆菌纲和毛螺菌目微生物的含量,为研究miR-146a 通过改变宿主肠道微生物丰度来影响肠道健康状况提供参考。
关键词: miR-146a;小鼠;肠道;特异性敲除;微生物
中图分类号: S511;S502文献标志码: A 文章编号: 1001-411X(2023)02-0197-08
肠道微生物与动物的健康关系密切。动物肠道内微生物的种类多、数量大。这群微生物与宿主互利共生,因其组成和分布的不同,发挥的功能也不同。肠道微生物不仅可以在机体代谢过程中提供底物、酶和能量,同时其代谢产物如脂肪酸等对机体上皮细胞生长与分化有促进作用,此外还参与合成维生素以及各种离子吸收。肠道微生物与宿主在肠道黏膜表面的交流促进了免疫系统的形成,从而构建了重要的生物及免疫屏障[1]。随着相关研究的不断发展,肠道微生物不仅在维持机体健康中发挥作用,还参与了许多疾病的进程。Qin 等[2] 于2012 年進行了肠道微生物与Ⅱ型糖尿病的宏基因组关联分析,研究结果表明在糖尿病患者与非糖尿病患者的中国人群中,肠道微生物组成发生改变,Ⅱ型糖尿病患者产丁酸细菌种类减少。一项对人类和小鼠微生物菌群的研究表明肥胖个体具有更高的从饮食中获取能量的能力[3],其原因是肥胖与2 种主要细菌分支相对丰度的差异有关,肥胖个体厚壁菌门Firmicutes 的相对丰度增加,拟杆菌门Bacteroidota的丰度减少。此外,一项通过宏基因组分析比较同卵双胞胎和异卵双胞胎及其母亲的微生物群的研究,也支持了微生物多样性减少会促进热量收集的结论[4]。最近的研究同样表明,微生物基因计数低的个体有更多的全身炎症、肥胖、胰岛素抵抗和血脂异常[5]。
MicroRNAs(miRNAs) 是一类长度为18~25 核苷酸的非编码小RNA 分子,通过与靶基因mRNA结合,导致其翻译抑制或降解,在细胞增殖、分化以及其他生物过程中发挥关键作用[6]。其中miR-146a作为近些年来miRNAs 的研究热点之一,其研究最早由T a g a n o v 等[ 7 ] 报道, 通过鉴定脂多糖(Lipopolysaccharide,LPS) 诱导的人单核细胞系THP-1 中miRNA 表达谱发现,其中miR-146、miR-132 和miR-155 表达显著升高,且LPS 介导的miR-146a 表达的上调是以NF-κB 依赖的方式发生的。随着miR-146a 研究的深入,发现其还与癌症相关。结直肠癌(Colorectal cancer,CRC) 是世界上最常见的癌症之一。2018 中国癌症统计报告显示:我国结直肠癌发病率、病死率在全部恶性肿瘤中分别位居第3 位、第5 位[8]。研究发现miR-146a 作为结肠炎症和相关肿瘤发生的主要负调控因子,调节IL-17 反应,其高表达量与大概率延长结肠癌患者生存时间的表征相关,同时miR-146a 缺陷的小鼠易患结肠炎相关和散发的CRC,并呈现出IL-17 信号的增强[9-10]。早在1974 年Reddy 等[11] 对无菌大鼠与普通大鼠每周注射10 mg/kg 1,2?二甲基肼,连续注射20 周,发现无菌大鼠无一只患病,而常规大鼠有17% 患上结直肠癌,表明肠道菌群对结直肠癌有调控作用。微生物群可以通过炎性介质(如肿瘤坏死因子、NF-κB、白细胞介素和干扰素等)、微生物代谢物(如肠道微生物源脱氧胆酸、丁酸等) 以及环境因素(如饮食因素、身体不活动、环境污染物等) 等影响结直肠癌的进程[12]。
那么miR-146a 作为抑炎因子,是否参与宿主与微生物间的互作,进而影响肠道菌群的稳态?本文以miR-146a 条件性敲除小鼠为模型,通过采用16S rRNA 高通量测序技术,研究miR-146a 对小鼠肠道菌群分布以及种类等方面的影响,以期为miR-146a 与微生物互作的研究提供有益的数据支持。
1 研究方法
1.1 试验动物繁育
miR-146a 肠道特异性敲除小鼠(Conditionalknockout mice,CKO 鼠) 由赛业(广州) 生物科技有限公司制备,简单来说,应用CRISPR-Cas9 技术在miR-146a 基因外显子1 两端插入Flox 序列,经PCR 可鉴定出基因组插入Flox 序列的阳性鼠为对照组鼠(Flox 鼠);为繁育CKO 鼠,将Flox 鼠与villus-Cre 工具鼠进行杂交,其原理是villin-Cre 基因只在肠道中表达Cre 酶活性,Cre 酶可将2 个Flox 序列中间的片段剪切,从而达到敲除的目的,具体繁育方法见下文。小鼠为普通SPF 级,均购自广东省医学实验动物中心(广东佛山)。CKO 鼠的制备:首先将F0 代Flox+/+纯合小鼠与villin-Cre 小鼠杂交,获得F1 代Flox+/?Cre+小鼠。接着把F1 代Flox+/?Cre+小鼠再与F0 代Flox+/+纯合小鼠杂交,获得F2 代Flox+/+Cre+鼠,即CKO 鼠。试验开始前进行小鼠基因型鉴定,取同一只小鼠的少量耳朵或尾巴组织对Flox(使用Flox 引物) 和Cre(使用villus-Cre 引物)(表1)2 个基因位点的PCR 产物进行琼脂糖凝胶电泳,Flox 鼠的FloxPCR 产物为单一206 bp 条带,Cre PCR 产物无明显条带;CKO 鼠的Flox PCR 产物为单一206 bp 条带,Cre PCR 产物为单一120 bp 条带。将基因鉴定后的公鼠分为Flox 组与CKO 组各4 只,单笼饲养,保证充足的饮水与饲料。
1.2 样品采集
将6 周龄小鼠颈椎脱臼处死,用镊子配合手术剪沿腹中线刨开腹部,分离出空肠段,再用剪刀纵切铺平,用盖玻片刮出黏膜,转移至离心管用于后续16S rRNA 测序,组织部分转移至新的离心管用于后续RNA 提取。
1.3 RNA 提取和qRT-PCR
使用 EZB 通用型 RNA 提取试剂盒(海方生物)提取组织总 RNA,并使用分光光度计(Nanodrop 2000,ThermoFisher) 测定 RNA 浓度。取 1 μg total RNA,使用 EZB 4×EZscript Reverse Transcription Mix II逆转录试剂盒进行逆转录,mRNA 使用Oligo(dT)18引物进行逆转录,miR-146a-5p 使用特异性茎环引物(mmu-miR-146a-5p RT:GTCGTATCCAGTGCGTGTCGTGGAGTCGGCAATTGCACTGGATACGACAACCCA) 逆转录。得到的cDNA 稀释 5倍,保存在?30 ℃ 冰箱待用。
qRT-PCR 使用 EZB 2× Color SYBR GreenqPCR Master Mix 进行目的基因miR-146a-5p(使用mmu-miR-146a-5p 引物) 和内参基因U6(使用mmu-U6 引物)(表1) 表达水平检测,基因表达结果使用 2?ΔCt 方法统计。
1.4 16S rRNA 测序
1 . 4 . 1 基因组D NA 的提取 样品总基因组DNA 用CTAB/SDS 法提取后在10 g/L 的琼脂糖凝胶上电泳检测DNA 的质量浓度和纯度,最后用无菌水将DNA 质量浓度稀释至l μg/μL。
1.4.2 扩增子一代 不同区域的16S rRNA/18SrRNA/ITS 基因用特异性引物(16S V4、18S V4、ITS1,表1) 进行扩增。参照Phusion?High-FidelityPCR Master Mix (New England Biolabs) 说明书进行PCR 反应。
1.4.3 PCR 产物定量和鉴定 将等量的1×上样缓冲液与PCR 产物混合,在20 g/L 琼脂糖凝胶上进行电泳检测并用Qiagen Gel Extraction Kit (Qiagen)进行PCR 产物纯化。
1.4.4 文库制备和测序 按照TruSeq?DNA PCRFreeSample Preparation Kit(Illumina) 说明书生成测序文库,并添加索引代码。使用Qubit?2.0 荧光计(Thermo Science) 和安捷伦生物分析仪2 100 系统对文库质量进行评估。随后在Illumina NovaSeq 平台上对文库进行测序,获得了250 bp 的成对末端阅读序列。
1.5 数据统计分析
对原始数据进行处理,得到有效数据(Cleand a t a ), 基于有效数据进行O T U s ( O p e r a t i o n a ltaxonomic units) 聚类和物种分类分析。根据OTUs 聚类结果,对每个OTU 的代表序列做物种注释,同时对OTUs 进行丰度、Alpha 和Beta 多样性计算等分析。为进一步挖掘分组样本间的群落结构差异,选用t 检验等统计分析方法对分组样本的物种组成和群落结构进行差异显著性检验。用Tax4Fun 软件对生态样本中的微生物群落进行功能预测分析。
2 结果与分析
2.1 肠道miR-146a 敲除小鼠的鉴定
提取Flox 组和CKO 组小鼠空肠组织的RNA,对2 组空肠组织的miR-146a-5p 进行实时荧光定量PCR 检测,结果表明,CKO 组小鼠空肠组织的miR-146a-5p 表达与Flox 组相比显著降低(图1)。
2.2 小鼠空肠微生物群落OTUs 分析
以97% 的一致性将序列聚類成为OTUs,共得到1 134 个OTUs,Flox 组、CKO 组分析共得到46 个相同的OTUs,但由于CKO 组间偏差较大,2 组之间并不存在显著差异。根据注释到的结果,所有样本在不同等级上的种类数量依次为界(Kingdom):2;门(Phylum):37;纲(Class):80;目(Order):161;科(Family):198;属(Genus):261;种(Species):117。
2.3 小鼠空肠微生物Alpha 多样性分析
样本内的微生物群落的丰富度和多样性用Alpha 多样性分析,分析结果如表2 所示。由表2 可知,各组的Good coverage 指数均为0.999,说明测序数据覆盖99.9% 的微生物,覆盖度较好。CKO 组的Shannon 指数低于Flox 组,Ace、Chao1、Observed_species 指数高于Flox 组,但差异均不显著(P>0.05)。
2.4 小鼠空肠微生物Beta 多样性分析
通过分析图2 中的不同样本点的距离来判断个体或群体间的差异,样本的群落组成越相似,则它们在主成分分析(Principal component analysis,PCA) 图中的距离越接近。
由图2 可知,PCA 显示Flox 组和CKO 组中各样本的组内偏差较大,其微生物群落组成相似。
2.5 小鼠空肠微生物门水平物种分布以及有差异的物种分析
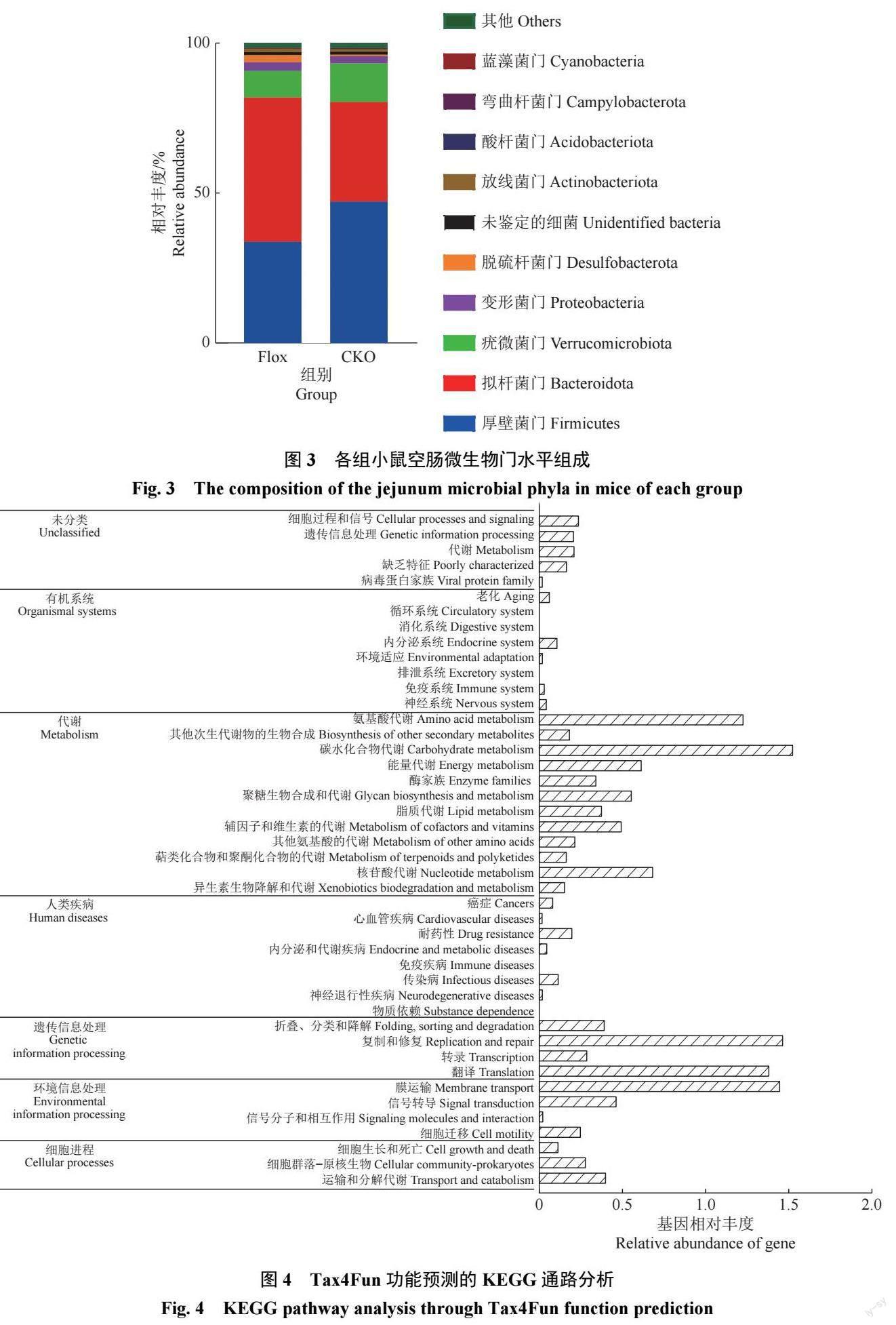
各组小鼠肠道微生物在门水平上,厚壁菌门Firmicutes、拟杆菌门Bacteroidota、疣微菌门Verrucomicrobiota、变形菌门Proteobacteria、脱硫杆菌门Desulfobacterota 丰度大于1%(图3)。
CKO 组厚壁菌门、疣微菌门相对丰度与Flox 组相比分别高38.24%、44.44%,而拟杆菌门相对丰度比Flox 组低31.25%,但均差异不显著(P>0.05)。从不同层级的物种丰度出发,通过常规的t 检验可以得到差异物种。在纲水平上,CKO 组和Flox组中梭状芽孢杆菌纲Clostridia 的平均相对丰度分别为7.32% 和2.23%,2 组间差异不显著(P=0.067);在目水平上,毛螺菌目Lachnospirales 在CKO 组和Flox 组中的平均相对丰度分别为1.40% 和0.50%,该物种在2 组间的差异显著(P=0.013 5)。
2.6 Tax4Fun 功能预测的KEGG 通路分析
各组样本Tax4Fun 预测到的功能主要集中在细胞进程、环境信息处理、遗传信息处理、人类疾病、代谢、有机体系统以及其他未分类的功能,其中与膜运输、遗传信息翻译、复制和修复、碳水化合物代谢、氨基酸代谢以及核苷酸代谢方面相关的基因相对丰度较高(图4)。
2.7 KEGG 通路Level 1 相对丰度分析
以Level 1 相对丰度柱形图(图5) 为例,2 组间的预测功能无显著差异,主要集中在代谢、遗传信息处理与环境信息处理上。
3 讨论与结论
微生物与宿主健康息息相关,例如肠道微生物多样性与长期体质量增加呈负相关,而与纤维摄入量呈正相关[13]。随着生猪养殖规模的不断扩大,封闭式猪舍越来越多,存在通风不顺畅的因素,导致封闭式猪舍内部存在大量的微生物、粉尘、内毒素等,会形成微生物气溶胶。微生物气溶胶浓度过高,就会影响生猪健康[14]。近来李喜阳等[15] 通过对腹泻和健康断奶仔猪肠道微生物比较发现,在门水平上,腹泻组厚壁菌门、变形菌门、放线菌门和Myxococcota 的相对丰度显著高于健康组,而拟杆菌门的相对丰度显著低于健康组;在属水平上,腹泻组中罗伊氏乳杆菌L a c t o b a c i l l u s r e u t e r i 、M a r v i n b r y a n t i a 、消化球菌属P e p t o c o c c u s 、Subgroup_10 和Bryobacter 的相对丰度显著高于健康组,说明仔猪肠道菌群的结构改变可能是仔猪发生腹泻的重要原因。越来越多的研究表明,不仅肠道微生物群的组成能调控宿主的代谢活动,单个微生物的代谢产物也可以作为信号分子在宿主的代谢中发挥重要作用。微生物可以通过分泌代谢产物向宿主的上皮细胞发出信号,如食物中不能被消化吸收的多糖在微生物糖苷酶的作用下生成短链脂肪酸(如乙酸、丙酸和丁酸),其可作为胃肠道上皮细胞的直接能源,还可通过体循环来调控宿主代谢[16];丙酸盐和醋酸盐可运输到肝脏来调控糖代谢和脂代谢进而抑制糖尿病与肥胖的发生,且醋酸盐还可以通过激活白色脂肪细胞中的G 蛋白偶联受体41 来调节能量平衡[17];常规传统方法与分子鉴定结果表明太子参内生细菌RPB-32 为芽孢杆菌Bacillus sp.。与空白对照组相比,给予内生菌代谢提取物的乙酸乙酯中剂量组、乙酸乙酯高剂量组、正丁醇低剂量组、正丁醇中剂量组及正丁醇高剂量组小鼠肠道乳酸菌数量显著增加,正丁醇中剂量组的小鼠肠道肠球菌和正丁醇高剂量组的小鼠肠道肠杆菌数量显著减少。初步判断太子参内生芽孢杆菌 RPB-32 代谢物(BM) 对肠道微生物组成有显著作用,并可能对机体糖脂降解、抗感染和抗炎等方面有改善作用[18];大豆异黄酮的保健功能主要归功于其肠道代谢產物 S?雌马酚。S?雌马酚是豆类食品在肠道内经特定微生物代谢后产生的一种高度稳定小分子,其与雌激素的结构和功能高度相似,能与β?雌激素 (ER-β) 受体结合,具有防治更年期综合征、心血管疾病和多种雌素依赖性癌症的功能[19]。
此外miRNA 可通过特有的作用方式影响宿主肠道微生物。有研究[20] 报道,生姜外泌体样纳米颗粒(Ginger exosome-like nanoparticle,GELN) 优先被乳酸杆菌科以GELN 脂质依赖性方式吸收,并含有靶向鼠李糖乳杆菌(Lactobacillus rhamnosus GG,LGG) 中各种基因的miRNA。其中,GELN mdomiR-7267-3p 靶向LGG 单加氧酶ycnE 产生更多的吲哚?3?甲醛(Indole-3-carbaldehyde,I3A),GELNRNAs或I3A 是芳烃受体的配体,诱导IL-22 的产生,而IL-22 与屏障功能改善有关。GELN-RNA 可以通过IL-22 依赖性机制改善小鼠结肠炎。Guo 等[21]发现,口服miRNA-10a-5p 可显著防止体质量增加,并改善高脂日粮喂养小鼠的葡萄糖耐量和胰岛素敏感性,但破坏了颤螺菌属Oscillospira、瘤胃球菌属Ruminococcus 和毛螺菌科Lachnospiraceae 的昼夜波动。miRNA 不仅存在于细胞外囊泡中,在小鼠和人类粪便样本中含量也丰富。宿主肠道上皮细胞(Intestinal epithelial cells,IEC) 和Hopx+ 细胞是粪便 miRNA 的主要来源,这些 miRNA 可以进入细菌,如具核梭杆菌Fusobacterium nucleatum 和大肠埃希菌Escherichia coli,特异性调节细菌基因转录,并影响细菌生长。 IEC-miRNA 缺陷 (Dicer1△IEC) 小鼠表现出肠道微生物群紊乱和结肠炎加剧,而野生型小鼠粪便 miRNA 移植可恢复缺陷小鼠粪便微生物结构并改善结肠炎。这些发现既确定了粪便miRNA 塑造肠道微生物群的生理作用,也确定了操纵微生物组的潜在策略[22]。
miR-146a 作为一个抑炎因子,在脊髓损伤大鼠中的表达低于对照组,而TLR/NF-κB 信号通路相关基因和炎性细胞因子表达上调,在注射agomiR-146 后,TLR/NF-κB 信号通路中的炎症基因和细胞因子(IL-6 和IL-8) 明显下调[23]。与此类似,在急性痛风性关节炎大鼠模型中,miR-146a 高表达组能显著改善关节肿胀指数、关节功能障碍指数和关节炎症指数。同时,miR-146a 的高表达明显抑制滑膜组织中TLR4、MyD88、相关炎症因子和NF-κB 的表达[24]。新生儿坏死性小肠结肠炎(Neonatal necrotizingenterocolitis,NEC) 是新生儿重症监护病房最常见的胃肠道急症,病死率达30%。近年来,多项研究表明,早在NEC 发病前数天至数周,NEC 病儿已经发生肠道菌群失调且革兰阴性杆菌丰富度增多,即生态失调早于 NEC 发病[25]。以上证据说明肠道微生物菌群失调会导致肠道炎症,而miR-146a 作为抑炎因子在许多炎症进程中发挥调控作用。
本研究中,肠道miR-146a 特异性敲除能显著提高毛螺菌目Lachnospirales 菌群的表达丰度。Zeng 等[ 2 6 ] 为了验证长期饲喂高脂料(High-fat,HF) 会加速炎症过程和改变肠道菌群组成的假说,将C57BL/6 小鼠分别饲喂高脂(45% 能量) 和低脂(Low-fat,LF,10% 能量) 饲料36 周。研究结束时,HF 组的体质量比LF 组重35%,喂食 HF 饮食的小鼠血浆瘦素、IL6 和TNF-α浓度也升高,表明存在慢性炎症,提取粪便 DNA 进行16S rRNA 测序,发现与 LF 组相比,HF 组厚壁菌门、毛螺菌科以及毛螺菌科/链球菌科比例显著升高,而这些细菌与代谢紊乱、糖尿病和结肠癌的发展有关。在他们的另一项有关非酒精性脂肪性肝病(Non-alcoholic fatty liverdisease,NAFLD) 跟肥胖饮食关联的研究发现,晚期脂肪肝发展与喂养HF 饮食小鼠的肝脏炎症、结肠继发性胆汁酸及其相关细菌水平升高同时发生,其中包括梭状芽孢杆菌纲和毛螺菌科的丰度增加[27]。
关于早期肠道微生物与特应性皮炎( A t o p i cdermatitis,AD) 之间的关系的研究,缺乏机制的了解和确定的结果, 研究者调查了1 2 例6 月龄AD 婴儿和12 例健康婴儿肠道微生物区系的多样性和组成,发现AD 患儿梭状芽孢杆菌的相对丰度与AD 发病年龄相关,与血液中嗜酸性粒细胞百分比呈负相关[ 2 8 ]。也有报道证明与健康组相比,从NEC 早产儿粪便样本中分离到更多的酪酸梭菌和新生儿梭状芽胞杆菌,并且这些菌具有更高的存活率与耐气性[29]。上述资料提示,miR-146a 可能作用于肠道微生物,进而调节肠道的发育与功能,但其具体机制还需作进一步研究探讨。
综上所述,本研究利用miR-146a 肠道特异性敲除模型,首次研究了宿主miR-146a 敲除对肠道微生物的影响,结果表明miR-146a 敲除可改变宿主肠道微生物的含量, 即显著增加毛螺菌目Lachnospirales 的平均相对丰度(P<0.05),同时梭状芽孢杆菌纲Clostridia 的平均相对丰度有增加趋势(P=0.067),为进一步研究miR-146a 通过影响宿主肠道微生物来调节肠道炎症进程提供了参考。
参考文献:
OLSZAK T, AN D, ZEISSIG S, et al. Microbial exposureduring early life has persistent effects on natural killerT cell function[J]. Science, 2012, 336(6080): 489-493.
QIN J, LI Y, CAI Z, et al. A metagenome-wide associationstudy of gut microbiota in type 2 diabetes[J].Nature, 2012, 490(7418): 55-60.
TURNBAUGH P J, LEY R E, MAHOWALD M A, et al.An obesity-associated gut microbiome with increased capacityfor energy harvest[J]. Nature, 2006, 444(7122):1027-1031.
TURNBAUGH P J, HAMADY M, YATSUNENKO T,et al. A core gut microbiome in obese and lean twins[J].Nature, 2009, 457(7228): 480-484.
LE CHATELIER E, NIELSEN T, QIN J, et al. Richnessof human gut microbiome correlates with metabolicmarkers[J]. Nature, 2013, 500(7464): 541-546.
BARTEL D P. MicroRNAs: Genomics, biogenesis,mechanism, and function[J]. Cell, 2004, 116(2): 281-297.
TAGANOV K D, BOLDIN M P, CHANG K, et al. NF-κB-dependent induction of microRNA miR-146, an inhibitortargeted to signaling proteins of innate immuneresponses[J]. Proceedings of the National Academy ofSciences of the United States of America, 2006, 103(33):12481-12486.
中華人民共和国国家卫生健康委员会. 中国结直肠癌诊疗规范2020 版[J]. 中华消化外科杂志, 2020, 19(6):563-588.
曾长青, 黄良祥, 郑羽, 等. miR-146a 在结肠癌中的表达及意义[J]. 南方医科大学学报, 2014, 34(3): 396-400.
REDDY B S, WEISBURGER J H, NARISAWA T, et al.Colon carcinogenesis in germ-free rats with 1, 2-dimethylhydrazineand N-methyl-N'-nitro-N-nitrosoguanidine[J]. Cancer Research, 1974, 34(9): 2368-2372.
孔垂秀. 综述微生物群与结直肠癌的关系研究[J]. 中国药物与临床, 2021, 21(23): 3839-3842.
MENNI C, JACKSON M A, PALLISTER T, et al. Gutmicrobiome diversity and high-fibre intake are related tolower long-term weight gain[J]. International Journal ofObesity, 2017, 41(7): 1099-1105.
温新水. 封闭式猪舍内微生物气溶胶对生猪的影响[J].畜牧兽医科技信息, 2019(12): 56.
李喜阳, 王中齐, 梁璐, 等. 腹泻和健康断奶仔猪肠道微生物比较分析[J]. 黑龙江畜牧兽医, 2021(17): 83-88.
CLARKE G, STILLING R M, KENNEDY P J, et al.Minireview: Gut microbiota: The neglected endocrine organ[J]. Molecular Endocrinology, 2014, 28(8): 1221-1238.
BAOTHMAN O A, ZAMZAMI M A, TAHER I, et al.The role of gut microbiota in the development of obesityand diabetes[J]. Lipids in Health and Disease, 2016, 15:108.
王俊丽, 欧阳湖, 谢虹雅, 等. 太子参内生细菌RPB-32 的分类鉴定及其代谢物对小鼠肠道微生物群落的影响[J]. 微生物学通报, 2021, 48(2): 502-515.
李海亮, 邓颖, 王欣, 等. 肠道微生物代谢产物: S?雌马酚与人类健康关系研究进展[J]. 中国微生态学杂志,2018, 30(3): 362-367.
TENG Y, REN Y, SAYED M, et al. Plant-derived exosomalmicroRNAs shape the gut microbiota[J]. Cell Host &Microbe, 2018, 24(5): 637-652.
GUO Y, ZHU X, ZENG S, et al. miRNA-10a-5p alleviatesinsulin resistance and maintains diurnal patterns oftriglycerides and gut microbiota in high-fat diet-fedmice[J]. Mediators of Inflammation, 2020, 2020:8192187. doi: 10.1155/2020/8192187.
LIU S, CUNHA A P D, REZENDE R M, et al. The hostshapes the gut microbiota via fecal microRNA[J]. CellHost & Microbe, 2016, 19(1): 32-43.
LV Z, CAO X, GUO Y, et al. Effects of miR-146a on repairand inflammation in rats with spinal cord injurythrough the TLR/NF-κB signaling pathway[J]. EuropeanReview for Medical and Pharmacological Sciences, 2019,23(11): 4558-4563.
CHEN X, GAO Q, ZHOU L, et al. MiR-146a alleviatesinflammation of acute gouty arthritis rats throughTLR4/MyD88 signal transduction pathway[J]. EuropeanReview for Medical and Pharmacological Sciences, 2019,23(21): 9230-9237.
王英豪, 戴立英. 新生儿坏死性小肠结肠炎与肠道微生物群落关系的研究进展[J]. 安徽医药, 2021, 25(12):2346-2350.
ZENG H, ISHAQ S L, ZHAO F, et al. Colonic inflammationaccompanies an increase of β-catenin signalingand Lachnospiraceae/Streptococcaceae bacteria in thehind gut of high-fat diet-fed mice[J]. The Journal of NutritionalBiochemistry, 2016, 35: 30-36.
ZENG H, LARSON K J, CHENG W, et al. Advancedliver steatosis accompanies an increase in hepatic inflammation,colonic, secondary bile acids and Lactobacillaceae/Lachnospiraceae bacteria in C57BL/6 mice fed ahigh-fat diet[J]. The Journal of Nutritional Biochemistry,2020, 78: 108336.
LEE E, LEE S, KANG M, et al. Clostridia in the gut andonset of atopic dermatitis via eosinophilicinflammation[J]. Annals of Allergy, Asthma & Immunology,2016, 117(1): 91-92.
SCHONHERR-HELLEC S, KLEIN G L, DELANNOYJ, et al. Clostridial strain-specific characteristics associatedwith necrotizing enterocolitis[J]. Applied and EnvironmentalMicrobiology, 2018, 84(7): e02428-17.
【責任编辑 庄 延】
猜你喜欢
新民周刊(2022年27期)2022-08-01
传染病信息(2021年6期)2021-02-12
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
食品与发酵工业(2020年16期)2020-01-07
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年8期)2019-11-23
实用皮肤病学杂志(2015年4期)2015-12-22
医学研究杂志(2015年12期)2015-06-10
