
基于Landsat 8 OLI 的云南元江流域森林生物量光学遥感估测及其饱和点分析
2023-03-31 08:04李紫荆刘彦枫欧光龙
西南林业大学学报 2023年1期
李紫荆 刘彦枫 冉 娇 欧光龙 胥 辉
(西南林业大学林学院,西南林业大学西南地区生物多样性保育国家林业局重点实验室,云南 昆明 650233)
全球碳循环的相对重要组成部分是陆地生态系统循环,其中森林生态系统碳汇潜力评估的重要基础是森林地上生物量,森林地上生物量的重要性决定了对森林生物量的精准估测已然成为研究地球碳循环和全球气候变化的重要内容之一[1-3]。传统的生物量估算方法周期长、成本高、耗费大量人力,而目前森林生物量及碳贮量的估测则更多地依赖于发展得不仅快速并且稳定的遥感技术[4-5]。不同于传统生物量估测技术,遥感技术具有快速、准确、对森林无破坏性并能进行宏观监测的优势,成为获取森林地上生物量的主要途径[6-7]。常用的遥感估测模型有参数模型的多元线性回归模型和非参数模型的人工神经网络模型、支持向量机回归模型、随机森林回归模型等[8]。众多学者在利用现代遥感技术去估测生物量时发现其存在诸多不确定性问题,不确定性问题来源于遥感数据源获取时影像分辨率、野外实测样地数据采集、森林空间异质性以及生物量估测模型等[9]。其中,遥感估测中存在光饱和点的问题尤为突出,即当森林植被密度达到一定阈值时,遥感影像就会出现光饱和点问题,从而无法准确估测研究区高生物量分布区情况[1,10-11],造成森林植被密度高地区高值低估的问题。光饱和值不仅受到森林结构的影响,也会因传感器分辨率的高低而发生变化,因此,基于光饱和点估测森林生物量时,在生物量估测模型上寻求合适的遥感模型提高生物量估测精度已成为学者们的研究热点之一[12-14]。现代遥感技术估测森林生物量主要通过建立如多元线性逐步回归、偏最小二乘法、数量化法等参数模型,然而当针对森林生物量较高的研究区进行生物量估测时,不能很好地拟合模型,便出现了高值低估的情况,但非参数模型相较于参数模型能够有效克服这种缺陷。孙雪莲等[13]、吴娇娇等[14]利用随机森林模型以及BP 神经网络模型提高了思茅松生物量遥感估测精度;卢腾飞等[12]、徐小强等[15]以探索光饱和现象造成的估测精度问题,利用混合效应模型以及支持向量机回归模型构建不同非参数模型对森林生物量遥感进行估测并得到了较好实验结果。以上研究表明非参数模型对提高森林生物量估测精度具有较大的潜力。
元江流域贯穿云南省中部、东南部并流经广西壮族自治区西南部。流域内具有两种不同的地貌特征,大致以元江为界,其东北部多属滇东高原,高原上石灰岩广布,具有明显的喀斯特地貌;西南部为西北东南走向的哀牢山,山区地形复杂,具有非常重要的生态价值。国内众多学者已对元江流域进行了大量的研究[16-17],但是这些研究大多集中在干热河谷的景观格局、植物区系等方面[18-19],但对于该流域内森林生物量的遥感估测还未见报道。基于此,本研究拟以云南省元江流域11 种优势树种作为研究对象,以2016年的森林资源二类调查数据和同期Landsat 8 OLI遥感影像为数据源,通过构建参数模型与非参数模型估测研究区森林生物量,反演研究区11 种优势树种地上生物量,并探索森林生物量估测中光学遥感数据源的光饱和点确定方法,以期为其他地区森林生物量遥感估测以及光学遥感数据源光饱和点的确定提供方法参考,实现对森林生物量的精准估测。
1 研究区概况
元江流域地处100°06′~105°40′ E、22°27′~25°32′ N,该流域起始于中国云南省西部哀牢山东麓,其地势西北高东南地,形呈狭弓带状,海拔高度在300~1 200 m,平均宽度约120 km,元江流域平坝面积很小,绝大部分属于山区。元江流域土壤类型有10 种,其中红壤和紫色土分布广泛,在海拔800 m 以下的土壤主要是燥红土,较高的土壤丰富度具有极高的研究价值。元江流域属于亚热带高原季风气候,年均温15~24 ℃,因受季风气候影响,导致该地区降水量丰富,年降水量在1 100 mm 左右。该流域属国际河流,南流入南海,江水年出境水量为479 亿m3,除农业少量耗水外,占河口入海水量的四成以上,占中国入海总水量17 243 亿m3的2.8%。元江流域森林资源极其丰富,森林覆盖率达到66.67%,其中云南松(Pinus yunnanensis)、思茅松(Pinus kesiyavar.langbianensis)、华山松(Pinus armandii)、橡胶(Hevea brasiliensis)、杉木(Cunninghamia lanceolata)、桉树(Eucalyptus robusta)常绿阔叶树等作为优势树种在该区域分布[18-20]。
2 材料与方法
2.1 数据来源
2.1.1 森林资源二类调查数据收集与处理
从相关林业部门获取2016 年森林资源二类调查数据,根据二类调查数据利用蓄积量-生物量的转换公式完成生物量的计算,计算方法见表1。

表1 生物量模型Table 1 Biomass models
本研究为提高生物量估测的精度,选择研究树种平均胸径大于6 cm 的小班,再利用ArcGIS 10.5 分别对各树种样本进行筛选(按照7∶3 的比例确定建模样本数和检验样本数),采用3 倍标准差法筛选生物量数据以减小误差。小班尽可能的为近纯林,且保证小班内部不包括其他非植被像元。可通过目视解译的方法剔除含有其他地类的小班,从而提高数据的准确性,具体情况如表2。

表2 小班分布情况Table 2 The distribution of sample plots from subcompartment
2.1.2 遥感数据收集与整理
遥感数据源为地理空间数据云(http://www.gscloud.cn/)上下载的与森林资源二类调查数据同期的Landsat 8 OLI 遥感影像数据以及元江流域DEM 数据,数据分辨率为30 m,相关信息见表3。

表3 研究区 Landsat8 影像基本信息Table 3 The basic information of Landsat 8 images in study area
2.2 遥感特征因子提取及筛选
植被定量遥感将地形效应定义为传感器接收到的地表斜面与地表水平面辐射亮度的差异,是地物相对于光源和传感器位置的函数[21-22]。
以云南省云江流域森林资源二类调查数据中11 种优势树种小班面状矢量数据为单位,利用ArcGIS 10.5 软件其中一个功能“分区统计”,在地理坐标相同的情况下将遥感特征因子数据垂直投影小班面状矢量数据上层相叠加,光学遥感估测森林生物量的关键在于采用统计检验的方法来确定影响因子。本研究提取了3 类遥感因子,分别为原始单波段因子、植被指数因子、信息增强因子(进行穗帽变换(K-T)和主成分分析(PCA)),具体见表4。皮尔逊相关性分析是针对森林生物量与遥感因子之间的,选择相关性显著(P<0.05)的遥感特征因子作为建模备选变量。

表4 遥感特征因子基本信息Table 4 Basic information of remote sensing characteristic factors
2.3 生物量遥感估测饱和点的确定方法
根据地上生物量与各波段光谱反射率相关性(P< 0.05),选择生物量(X 轴)与相关性最为显著的波段反射率(Y 轴)做散点图,发现波段反射率会随着生物量的增大逐步减小,当生物量达到一定水平时波段反射率不再发生变化,该水平值即为光学遥感估测生物量时的饱和点。参考赵盼盼等[29]和孙雪莲等[30]的方法,利用参数估计方对比9 种模型建模效果,得出二次项、三次项模型能较好的拟合曲线方程,如公式(1)。本研究结合最值的求解方法,构建单波段信息与生物量之间的曲线关系,选用立方项模型求解光学遥感生物量的饱和点。
式中:h表示模型极值(光学遥感生物量饱和点);a、b、c、d分别表示模型参数。
2.4 生物量光学遥感估测模型的构建
2.4.1 模型选择
本研究选择2 种数学模型估测元江流域优势树种的地上生物量,其中,一种为参数模型多元线性回归(MLR),另一种为非参数模型支持向量机(SVM)。
多元线性回归生物量遥感模型见式(2)。
式中:Y为生物量, β0为 常数项, β1,β2,···,βn为模型系数,x1,x2,···,xn为相关遥感因子, ε为模型满足随机正态分布下的随机残差,n为自变量个数。
支持向量机方法在回归问题上的推广为支持向量回归(SVR),在很多方面都表现出较好的性能。支持向量机方法不仅实现了结构风险最小化原则并且保证即使在有限的样本情况下时仍具有较好推广能力,SVR 在解决回归问题上也具有相同的性能。因此,本研究将用SVR 来建立森林生物量遥感模型,以解决森林生物量遥感估测中的遥感因子与预测的森林生物量的非线性关系。SVR 算法的具体实现步骤:
1)获取训练样本集:(x1,y1), (x2,y2), ···,(xl,yl),xi∈Rn,yi∈Rn,i=1, 2, ···,l;
2)确定特征空间—选择径向基核函数;
3)对自由参数ϵ 的选择,确定经验风险;
4)将原二次规划问题转化为凸优化问题进行求解;
5)确定最优超平面,获得SVR 模型;
6)通过获得的模型对测试样本集进行预测,输出结果。
2.4.2 模型评价与独立性检验
模型评估是检验模型参考价值高低的重要组成部分,本研究评价模型精度主要使用以下2 种指标模型的决定系数(R2)、均方根误差(RMSE),模型的独立性检验指标选用相对均方根误差(rRMSE)、平均相对绝对误差值(MARE)和平均绝对误差(MAE),计算方法见式(3)~(7)。
式中:yi是实测值;是预测值;是样本平均值;n是样本总数。
2.4.3 “刀切法”残差分析
对各植被类型在森林生物量估测中存在的光饱和点进行确定时,能更好体现检验模型对高值和低值的处理情况。本研究采用“刀切法”残差检验,选取平均残差(ME)、相对平均残差(MRE)对各个模型在不同林组下的预估能力进行分析,RMSE 作为残差检验的评价指标。
式 中:yi是实测值;是预测值;n是样本总数。
3 结果与分析
3.1 生物量遥感因子相关性分析
本研究选取森林生物量估测的基础变量为45 个特征因子的均值。并利用统计检验的方法,在SPSS 中分析比较了各特征因子与地上单位面积生物量的相关性,选取显著相关性(P< 0.05)且独立性好的因子,其中海拔、坡度和坡向的相关性都未通过检验,故模型将不采用地形因子。
为确定不同植被的生物量饱和值,将云南省元江流域11 中优势树种森林生物量与Landsat 8 OLI 影像各波段的光谱反射率进行皮尔逊相关性分析,结果见表5。由表5 可知,B3、B6、B7 波段与11 中优势树种生物量均呈极显著相关(P<0.01),因此本研究基于B3、B6、B7 波段进行分析和确定云南省元江流域11 中优势树种森林生物量饱和值。

表5 光谱波段与生物量相关系数Table 5 Relationships between AGB and spectral band
结果表明:各类型植被波段与生物量均呈负相关,研究选择与各类型植被相关性最强的波段来确定其生物量饱和值。各类型植被生物量与相关性最强的波段反射率散点图均表现出随着生物量的增加反射率逐渐减小,直至趋于稳定。波段反射率趋于稳定时的值所对应的生物量为遥感估测生物量的饱和点。
3.2 生物量遥感估测光饱和值的确定
本研究采用三次项模型拟合曲线并求得其拐点的值,其拐点所对应的值即为11 种优势树种遥感估测森林生物量的饱和值。生物量采用立方项模型的主要原因是因为在波段变换区间内,它能够较好地反映极小范围内波段光谱值与生物量的关系,各植被类型的饱和点模型见表6。
为了进一步确定不同植被类型在光学遥感下的生物量饱和点,研究采用立方项模型求解不同植被类型与对应波段之间的函数关系,函数在区间内的极值所对应的自变量值为光学遥感生物量估测的光饱和值。通过各植被类型单波段的分析比较,最终选择决定系数R2最大的波段作为光学遥感估测的饱和点模型并绘制相关的饱和点曲线图,饱和点曲线见图1,得出各植被类型饱和值见表6。

图1 立方项模型模拟的不同植被生物量饱和曲线Fig. 1 Saturation curves of different vegetation biomass model
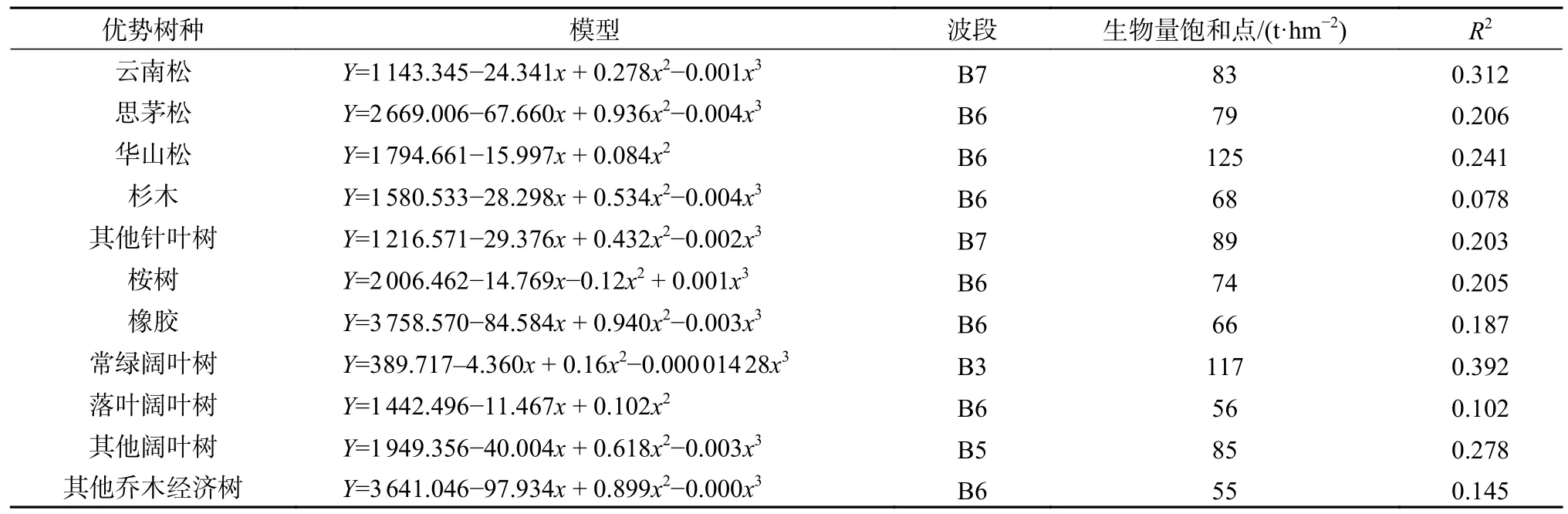
表6 不同植被光学遥感生物量估测模型以及饱和值Table 6 Biomass estimation model and saturation value of different vegetation by optical remote sensing
3.3 森林生物量遥感估测模型
3.3.1 多元线性回归模型结果
本研究采用SPSS 优选出自变量与森林生物量构建多元线性逐步回归模型。多元线性逐步回归模型中,自变量选择了显著性水平值为P< 0.05的,变量剔除选择水平设置为P> 0.1 的;同时调用VIF 函数对模型中各个自变量做多重共线性检验并选取VIF < 10 的遥感特征因子建立多元线性逐步回归模型。对各植被类型的线性回归分析,模型和精度评价见表7。

表7 多元线性回归模型精度评价结果和独立检验结果Table 7 Multiple linear regression model accuracy evaluation results and independent test results
通过模型的决定系数(R2)和均方根误差(RMSE)的计算发现,常绿阔叶树和其他阔叶树的模型决定系数最大,分别为0.423 和0.652。其他阔叶树的R2最高,云南松、华山松、其他针叶树、桉树和橡胶的R2均在0.2 以上但小于0.4,思茅松、杉木、落叶阔叶树和其他乔木经济树的R2均小于0.2。综上说明其他阔叶树的生物量模型最好,而常绿阔叶树次之,其他各植被类型的模型模型精度和独立性检验结果较差,原因需要进一步探索。
3.3.2 支持向量机回归建模结果
根据前文提到的模型精度评价方法,对支持向量机模型进行了泛化处理,提高了模型估测的稳定性。各树种的生物量估测模型拟合精度结果见表8。

表8 支持向量机模型精度评价结果和独立检验结果Table 8 Support vector machine model accuracy evaluation results and independent test results
由表8 可知,其他阔叶树的生物量模型精度最高,达到了0.841,其次是橡胶和常绿阔叶树生物量估测模型,其精度也达到了0.650 和0.524,其他各植被类型的模型精度和独立性检验结果较差。结合表7 和表8 可看出同一优势树种在2种模型下,支持向量机回归模型的模型精度以及检验指标均高于多元线性逐步回归模型精度以及检验指标,支持向量机模型在一定程度上提高了生物量的遥感估测精度,通过使用支持向量机模型,能够很大程度上提高生物量估测的能力。
3.3.3 “刀切法”残差分析
本研究将不同植被类型的生物量进行分段处理,对比在不同植被类型生物量的各个分段下模型的估测精度,结合表9 可知,2 种模型在不同生物量分段处其预估能力有所差异,根据ME 值的变化,发现各树种所对应的模型都存在着一定程度上的低值高估和高值低估的现象。ME < 0,表示低值高估;ME > 0,表示高值低估。
由表9 可知,11 种优势树种多元线性逐步回归模型在低生物量段(0~50 t/hm2)时,多元线性回归模型下其他乔木经济树ME(2.801)、MRE(-0.067)与支持向量机回归模型下其他乔木经济树ME(0.358)、MRE(-0.008)最低,与其他树种的模型比较,ME、MRE 绝对值降低了。多元线性回归模型下其他阔叶树在生物量段(50~100 t/hm2)有最好的表现,支持向量机模型下橡胶在生物量段(50~100 t/hm2)有最好的表现。橡胶对于2 种模型高生物量段(>150 t/hm2)的估测误差值最小。由此可见,2 个模型对各个植被类型不同生物量段的生物量估测能力的程度是不同的。
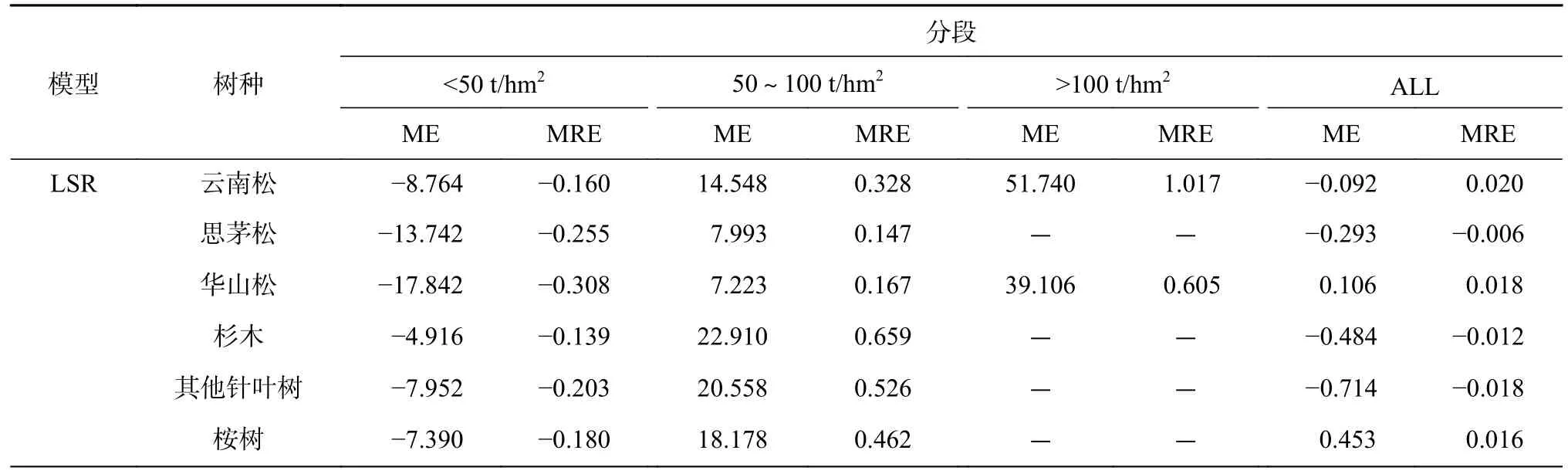
表9 “刀切法”残差检验结果Table 9 Jack knife error results

续表9
3.4 森林生物量反演
由图2 可知,多元线性回归模型下云南省元江流域中云南松林生物量集中在30~100 t/hm2,少数分布在100 t/hm2以上。思茅松林生物量主要在0~60 t/hm2,少数分布在60~100 t/hm2;华山松林生物量主要在30~70 t/hm2,少数分布在70~150 t/hm2;杉木林生物量主要集中在0~100 t/hm2,少数分布在100 t/hm2以上;其他针叶树林生物量主要集中在20~100 t/hm2,少数分布在100 t/hm2以上。桉树林生物量估测主要在0~100 t/hm2,少数分布在100 t/hm2以上;橡胶林生物量分布在0~100 t/hm2,少数分布在100 t/hm2以上。常绿阔叶林生物量主要在30~100 t/hm2之间,少数分布在30 t/hm2以下,100 t/hm2以上;落叶阔叶林生物量主要在60~100 t/hm2之间,少数分布在60 t/hm2以下,100 t/hm2以上;其他阔叶林生物量主要在0~100 t/hm2之间,少数分布在100 t/hm2以上。乔木经济树林生物量主要在0~50 t/hm2之间,少数分布在50 t/hm2以上。
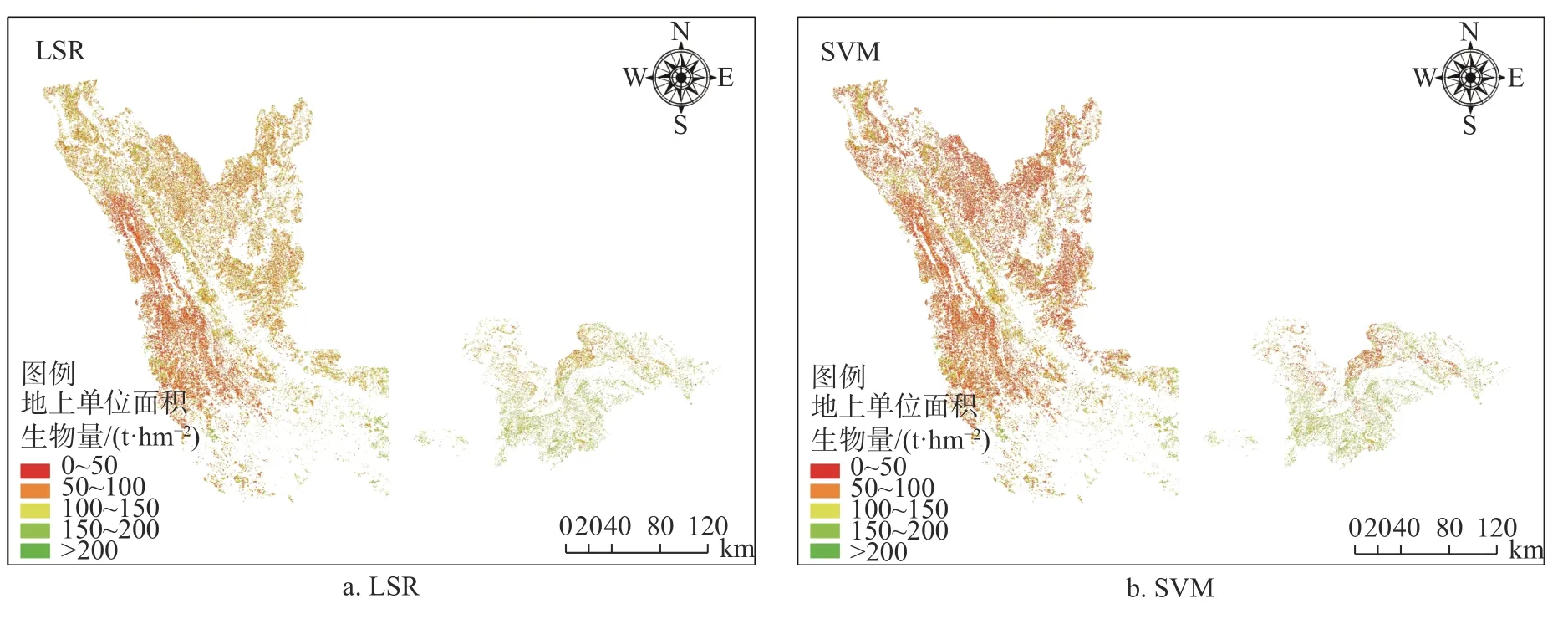
图2 2 种模型下11 种优势树种生物量反演Fig. 2 Biomass inversion of 11 dominant tree species based on support vector machine model
支持向量机回归模型下云南省元江流域中云南松林生物量集中在0~70 t/hm2,少数分布在70 t/hm2以上;思茅松林生物量主要在30~100 t/hm2,少数分布在30 t/hm2以下,100 t/hm2以上;华山松林生物量主要在0~100 t/hm2,少数分布在100 t/hm2以上;杉木林生物量主要集中在0~150 t/hm2,少数分布在150 t/hm2以上;其他针叶树林生物量主要集中在70~100 t/hm2,少数分布在70 t/hm2以下,100 t/hm2以上。桉树林生物量估测主要在0~100 t/hm2之间,少数分布在100 t/hm2以上;橡胶林生物量分布在0~100 t/hm2,少数分布在100 t/hm2以上。常绿阔叶林生物量主要在50~100 t/hm2,少数分布在50 t/hm2以下,100 t/hm2以上;落叶阔叶林生物量主要在0~100 t/hm2之间,少数分布在100 t/hm2以上;其他阔叶林生物量主要在0~100 t/hm2之间,少数分布在100 t/hm2以上。乔木经济树林生物量主要在0~50 t/hm2之间,少数分布在50 t/hm2以上。
4 结论与讨论
4.1 讨论
4.1.1 数据饱和
由于光谱影像受光谱反射率、辐射分辨率、空间分辨率等制约,光谱信息对生物量的敏感度会随着生物量的增大而降低,从而出现数据饱和现象[31]。单位面积生物量数据饱和情况是无法避免的,诸多森林生物量估测研究都发现了影响生物量估测精度的重要原因为数据饱和[28,32]。研究学者们采用了各种各样不同的方法来提高生物量的估测精度,大多以结合不同数据源及利用高光谱影像估测生物量来减少生物量的饱和、提高生物估测精度[33-36]。由于样地数据的不足,针对不同植被类型进行生物量饱和的研究也很少。本研究在庞大的二调数据支撑下,通过对比Landsat 8 OLI 影像的各个波段与不同植被生物量间的相关性,发现各植被类型均有与生物量呈相关性的波段。因此,研究利用不同类型植被最强波段来确定生物量的饱和值,并采用立方项模型对Landsat 8 OLI 影像估测元江流域不同森林类型的饱和值进行了定量的研究。结果表明常绿阔叶树具有最高的生物量饱和值,华山松、其他针叶树、其他阔叶树次之,云南松、思茅松、杉木、桉树、橡胶、落叶阔叶树、其他乔木经济树的饱和值均不高。
本研究采用植被分类的方法定量研究遥感估测森林生物量数据饱和的问题。采用不同的估测方法,可以减少生物量遥感估测的数据饱和,从而提高生物量模型的估测精度。其中阔叶树的估测精度基本高于针叶树种,在估测模型中针叶树数据饱和也较为严重。因此,仅使用光学遥感信息并不能解决数据饱和的问题,可以向自然环境和多源数据方面进行进一步探索。
4.1.2 建模方法
本研究采用相关性分析的方法从大量的遥感特征因子中选取与单位面积生物量密切相关且独立性强的自变量建立森林生物量遥感估测模型,这种特征的筛选的方法是依据线性相关的大小选取的。然而很多研究表明遥感特征因子与单位面积生物量间多存在明显的非线性特征。某些与生物量相关性较低特征与相关性较高的特征一起进入模型时,低特征会在很大程度上影响模型精度,所以利用线性相关性大小进行特征选取的合理性存在质疑。而且多元逐步回归模型必须满足一定的假设条件(正态、独立、线性、等方差),往往使得能够进入模型的变量较少(通常2~3 个),导致了遥感信息不充分利用,制约了模型的参考性[37]。
本研究采用了多元线性逐步回归和支持向量机回归2 种建模方法进行对比研究。通过对比发现,多元线性逐步回归在生物量估测中不能很好地反映实地生物量的分布情况,其基本参数模型的决定系数R²总体偏低,其结果与其他研究类似。而支持向量机回归模型有效描述森林生物量和遥感特征数据之间复杂的非线性关系。利用该模型各树种估测模型确定系数显著提高,较充分地利用了各类遥感特征信息,说明非参数模型比参数模型有更好的解释能力。
4.1.3 模型精度
本研究采用2 种建模方法得出的结果表明,支持向量机回归模型的精度较好。导致模型精度的原因有很多方面。本研究所用的建模样本为2016 年的森林资源二类调查数据,样地调查采用角规控制检尺的方法,样地大小不一,还受调查者经验及调查时天气影响等,导致调查精度不高,加之遥感影像在高山峡谷处易变形,导致与地面实际样地的空间位置匹配度降低,影响模型的精度。其次,建模样本数据中的生物量并非专业人士实地获取的实测生物量,而是用相似地区的蓄积量-生物量的转换公式,实际跟研究区实测样地数据存在差异,导致样地生物量的不准确性,从而影响模型精度。此外,Landsat 8 OLI 数据主要包括的是森林植被表层的光谱数据信息,缺少垂直结构的信息,并且在一定程度上还会出现遥感信息饱和,导致了生物量估测精度低。最后,由于地形、大气和传感器的不同等造成了遥感数据与地物间的对应关系极其复杂。
4.2 结论
本研究以元江流域为研究区,采用线性(多元线性逐步回归模型)和非线性(支持向量机回归模型)遥感估测方法,通过相关性分析确定最佳的生物量估测波段,采用立方项模型定量研究了元江流域11 种不同类型植被森林生物量的饱和值,研究表明:
1)各树种地上单位面积生物量与遥感特征变量的相关性均极为显著(P< 0.01),其中常绿阔叶最高达到0.615,杉木最低仅0.287,这是由于受到林分结构和树种间差异所致;
2)各个树种则具有不同的饱和阈值,其中华山松(125 t/hm2) > 常绿阔叶(117 t/hm2) > 其他针叶树(89 t/hm2) > 其他阔叶树(85 t/hm2) >云南松(83 t/hm2) > 思茅松(79 t/hm2) > 桉树(74 t/hm2) > 杉 木(69 t/hm2) > 橡 胶(66 t/hm2) > 落叶阔叶树(56 t/hm2) > 其他乔木经济树(55 t/hm2);
3)检验结果表明,多元线性逐步回归模型的精度较低,支持向量机回归模型精度高;在保证了足够样本数量的情况下,基于方法的不同建立多个生物量估测模型进行对比,能有效地提高生物量估测的精度。
本研究旨在通过改变建模方法,充分考虑林分异质性,提高遥感估测森林生物量的精度,减小数据饱和带来的不确定性,但其仍然存在一些不足,需要未来的研究加以重视。首先,本研究使用的多元线性回归模型,公式简单且便于计算,但线性关系很难反映描述出各个参数间的复杂非线性关系,模型泛化能力较差。因此开发出更具有适用性,同时能够减小因为数据饱和的存在造成的遥感估算误差的非线性和非参数建模方法是今后重要的研究课题。另外,本研究仅仅基于Landsat 8 OLI 遥感数据源对云南省元江流域的优势树种进行生物量的估测,遥感数据源单一,缺少多源遥感获取的方法及利用。最后,遥感估测森林生物量普遍存在低值高估与高值低估现象,减小数据饱和问题的影响主要是解决高值低估带来的误差,同样低值高估带来的误差也应引起重视,选择更合适的建模方法,以及充分利用遥感数据信息对提高整体森林生物量的遥感估测精度具有很重要的帮助,这一方面需要进一步的研究与实践。
猜你喜欢
数学物理学报(2022年4期)2022-08-22
中学生数理化·高一版(2021年2期)2021-03-19
今日农业(2020年19期)2020-12-14
江苏农业科学(2020年12期)2020-07-20
西安科技大学学报(社会科学版)(2019年1期)2019-09-10
中央民族大学学报(自然科学版)(2018年3期)2018-11-09
中国科技信息(2015年2期)2015-11-16
新课程·中旬(2014年10期)2014-12-26
植物营养与肥料学报(2014年1期)2014-03-11
中国烟草学报(2012年3期)2012-04-10
