
过表达和敲除NtabNAC087基因对烟草响应干旱胁迫的影响
2023-06-05 06:23王世泽杨志晓曹领改仪林小虎余世洲
核农学报 2023年7期
王世泽 刘 杰 杨志晓 曹领改 刘 勇, 宗 仪林小虎 余世洲,
(1河北科技师范学院农学与生物科技学院,河北 秦皇岛 066004; 2贵州省烟草科学研究院,贵州 贵阳 550081;3河南农业大学烟草学院,河南 郑州 450002; 4江苏中烟工业有限责任公司,江苏 南京 210000)
干旱是抑制作物生长、发育和产量的主要非生物胁迫之一[1]。作物遭受干旱胁迫后生长受到抑制,可导致作物减产乃至绝收[2]。前人通过对拟南芥(Arabidopsis thaliana)和水稻(Oryza sativaL.)NAC 基因家族进行综合分析发现,按照蛋白结构相似度和进化关系,可以将NAC 基因划分为两大类,共18 个亚组[3]。其中AtNAC3 和ATAF 亚组基因多被报道与干旱、盐害、冷害等逆境胁迫应答相关[4]。除了这两个亚组基因,NAP、NAM 和OsNAC3亚组也有少数基因具有响应逆境胁迫的功能[5]。
Tran 等[6]利用基因表达谱芯片发现,拟南芥AtNAC3 亚组成员基因ANAC019、ANAC055和ANAC072/RD26遭受干旱或盐胁迫后,基因表达显著上调,且构建的过表达植株在干旱胁迫下表现出抗旱性,证实了上述3 个基因能够响应干旱或盐胁迫。随后NAC 基因家族AtNAC3亚组成员基因或同源基因在多个物种中被证实具有抵御干旱胁迫的功能。如在玉米(Zea maysL.)中过表达ZmNAC49基因,能够影响气孔发育,降低气孔密度,从而提高玉米的耐旱性[7]。通过在拟南芥中异源表达玉米ZmSNAC13基因能够显著提高拟南芥植株的耐旱性[8]。在小麦(Triticum aestivumL.)中,通过正向遗传学和基因调控网络分析,发掘出小麦干旱响应基因TaSNAC8-6A,且证实了其具有抗旱功能[9]。在茄科物种NAC 基因家族相关研究中,证实了过表达马铃薯(Solanum tuberosum)StNAC053和番茄(Solanum lycopersicum)SlNAC6基因,可使植株对干旱胁迫处理的耐受性显著提高,同时维持较低的水分损失率和氧化损伤程度,以及较高的脯氨酸含量和抗氧化酶活性[10]。另外在木薯(Manihot esculentaCrantz)和芹菜(Apium graveliensL.)研究中,分别发现拟南芥ANAC072/RD26同源基因MeRd26[11]和AgNAC63[12]受干旱胁迫诱导后表达量显著上升,具有抗旱作用。
本研究前期通过生物信息学手段筛选到与拟南芥ANAC072/RD26基因的同源烟草(Nicotiana tabacumL.)基因NtabNAC087[13],且通过基因表达谱数据发现干旱胁迫下表达量上调。为了进一步鉴定NtabNAC087基因是否参与干旱胁迫应答,本研究以具有参考基因组序列的主要栽培烟草品种K326为试验材料,通过遗传转化、CRISPR/CAS9 技术及烟草生育期调控技术[14],构建NtabNAC087基因的过表达和基因敲除纯合阳性株系(T2),并在此基础上,观测不同烟草株系材料在遭受干旱胁迫后植株表型和基因表达情况,以期为进一步研究NtabNAC087的分子调控网络及育种利用提供帮助。
1 材料与方法
1.1 试验材料
普通烟草品种K326(野生型/对照材料)种子,遗传转化过程中使用的大肠杆菌DH5α、Top10 感受态细胞、农杆菌EHA105菌株和载体pBWA(V)HS均由贵州省烟草科学研究院保存并提供,前期研究初步筛选得到的NtabNAC087基因(Nitab4.5_0000662g0120.1)[13]序列在茄科基因组网站(https://solgenomics.net/)下载得到。
1.2 试验方法
1.2.1 烟草NtabNAC087基因的克隆及转基因植株的构建 利用总RNA 小量制备试剂盒(爱思进,杭州)提取烟草总RNA,并用EasyScript One-Step gDNA Removal and cDNA Synthesis SuperMix 试剂盒(全式金生物,北京)进行RNA 反转录。使用Primer6 软件[15]对NtabNAC087基因进行PCR 扩增引物设计(引物信息见表1)。PCR体系为20 μL:DNA 3 μL,2× Taq Master Mix 10 μL,10 μmol·L-1正反引物各1 μL,蒸馏水补至20 μL。PCR反应条件:95 ℃预变性2 min;95 ℃变性15 s,60 ℃退火20 s,72 ℃延伸2 min,35个循环;72 ℃终延伸7 min。选择1%琼脂糖凝胶电泳,并用DNA 凝胶回收试剂盒(爱思进,杭州)回收目的片段,随后将回收的NtabNAC087基因片段与pBWA(V)HS 载体片段利用T4 连接酶进行连接。遗传转化流程参考文献[16]。
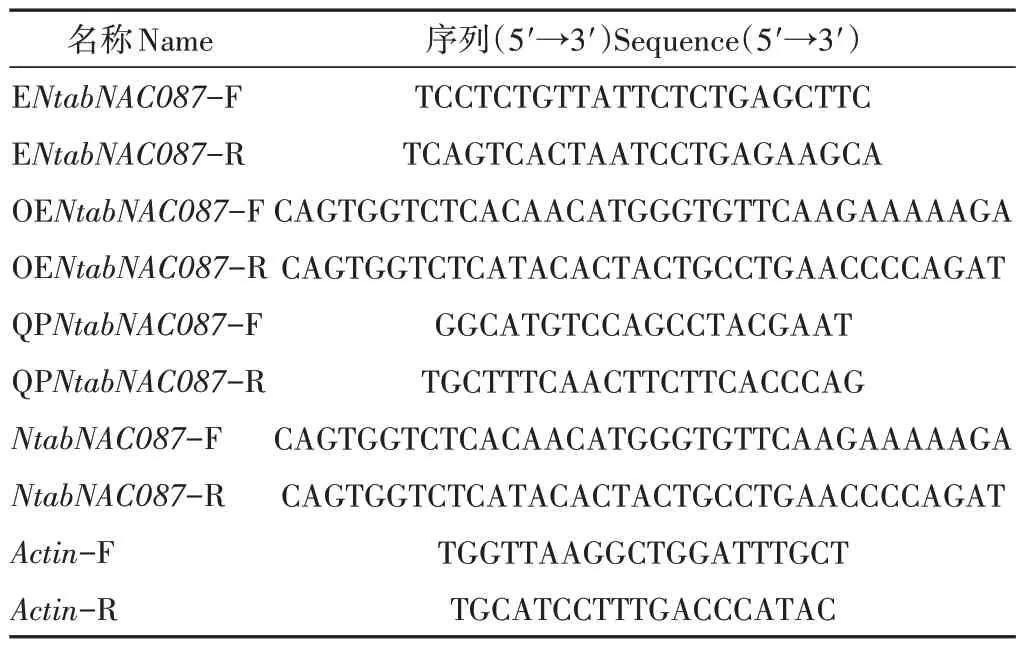
表1 所用引物序列Table 1 The sequence of the primers used in this experiment
通过CRISPR-P 2.0(http://crispr.hzau.edu.cn/CRISPR2/)在线网站设计CRISPR/Cas9 敲除sgRNA,基因敲除载体构建及遗传转化委托武汉伯远生物科技有限公司完成。利用琼脂糖凝胶电泳检测获得的T0代植株是否为阳性,对阳性植株进行早花处理[17],在贵州省烟草科学研究院人工气候室进行连续加代,得到T2代材料,挑选基因表达量较高的过表达植株及编辑位点纯合的编辑植株做为后续的试验材料。
1.2.2 干旱胁迫试验设计 将NtabNAC087基因过表达植株、编辑植株和野生型K326 种子放入含有20%PEG6000的MS固体培养基中进行种子萌发试验,观察种子萌发10 d后根的生长情况。
苗期试验处理流程:将种子放入含Hoagland 营养液的培养皿中进行催芽,待发芽后移栽到泥炭土中进行培养,在16 h 光照/8 h 黑暗、28 ℃/23 ℃培养箱内生长,设置干旱处理组,自然胁迫(移栽成活后不浇水)处理15 d,观察并记录植株的萎蔫程度,对照组烟草植株正常补充水分。
1.2.3 数据调查 待对照组烟草植株播种后45 d时,利用SU8010扫描电镜(日立,日本)对其叶片组织气孔形态进行观察,利用Nano Measure 1.2 软件统计气孔密度、气孔长度和气孔宽度,每个样品设置5 个生物学重复。待烟草植株播种后45 d时,将其移入Hoagland营养液中进行水培,60 d时进行20% PEG6000模拟干旱处理。在干旱处理0、1、3、6 和12 h 时分别对植株叶片取样,液氮速冻,-80 ℃保存。采用脯氨酸(proline,Pro)、过氧化氢酶(catalase,CAT)、超氧化物歧化酶(superoxide dismutase,SOD)和丙二醛(malondialdehyde,MDA)含量试剂盒(苏州格锐思生物科技有限公司)测定不同材料中各成分的酶活及含量。以烟草Actin基因作内参基因,使用TB Green®Premix DimerEraserTM(Perfect Real Time)试剂盒(TaKaRa,大连)进行qRTPCR(quantitative real-time PCR)测定基因的表达量(所用引物见表1)。反应体系为20 μL:cDNA 2 μL,TB Green Premix DimerEraser 10 μL,RoxⅡ0.4 μL,10 μmol·L-1正反引物各0.6 μL,用蒸馏水补至20 μL。反应条件为:95 ℃预变性30 s;95 ℃变性10 s,60 ℃退火20 s,40个循环。设置3次生物学重复。
1.3 数据分析
使用Microsoft Excel 2007 软件进行数据计算和图表绘制,使用SPSS 19.0软件进行数据分析。
2 结果与分析
2.1 NtabNAC087的基因过表达及基因敲除
NtabNAC087基因的开放阅读框长度为1 766 bp,共编码342 个氨基酸,分子量为39.046×103,含有NAC超家族结构域,属于AtNAC3 亚族成员。共获得24 株T0代过表达株系材料,结果表明96%的过表达植株为阳性(图1-A)。对阳性植株进行连续早花处理后得到T2代材料,筛选得到5 个稳定株系(oeNAC087-1~oeNAC087-5)。
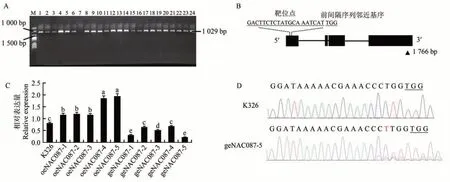
图1 NtabNAC087的基因过表达及基因敲除检测结果Fig.1 Results of gene overexpression and gene knockout detection of NtabNAC087
对NtabNAC087基因结构进行分析可知含有4 段编码序列(coding sequences,CDS)区(图1-B),将第一个外显子区5′→3′方向118~137 bp 区域的20 bp 碱基序列为靶点编辑位点进行NtabNAC087基因敲除,序列(5′→3′)为GACTTCTCTATGCAAATCATTGG。共得到10 株T0代编辑株系材料,进行加代后得到5 个稳定株系(geNAC087-1~geNAC087-5)。
分别对不同株系的基因表达量进行测定。待T2代材料生长45 d左右时,对NtabNAC087基因表达量进行检测,过表达植株中NtabNAC087基因表达量均显著高于对照K326,编辑植株中有3个株系NtabNAC087基因表达量与对照组存在显著差异(图1-C)。其中过表达株系oeNAC087-5的基因表达量最高是对照的14倍左右;基因编辑株系geNAC087-5 的基因表达量最低,是对照的0.27 倍,选择这两个株系做后续的干旱胁迫及自然干旱相关试验。同时对基因编辑株系geNAC087-5提取DNA后进行测序,发现其编辑位点存在胸腺嘧啶T碱基的插入,导致NtabNAC087基因移码突变(图1-D)。
2.2 苗期植株表型鉴定
由图2-A 可知,在PEG6000 干旱胁迫处理下,种子在萌发阶段根长的发育存在差异,其中编辑植株的根最短,过表达植株的根相比对照组较长。由图2-B可知,苗期进行自然干旱处理后,3 种植株叶片都表现出不同程度萎蔫,基因敲除植株叶片萎蔫程度最大,过表达植株萎蔫程度最小,由此推测,NtabNAC087基因可以提高烟草的抗旱性。同时,对3 种烟草植株叶片进行扫描电镜观察,在同一面积内对气孔密度进行统计,结果如图2-C 所示。对照K326植株的平均气孔密度为12 个,过表达植株为18 个,编辑植株为12 个,过表达植株的气孔密度高于其他两组。通过对保卫细胞进行观察,发现对照K326 植株细胞较为饱满,细胞外露,而过表达植株细胞较小且在气孔内嵌,编辑材料的气孔形态介于两者之间(图2-D)。
由图3 可知,过表达植株叶片气孔长度相比于对照组及编辑植株气孔长度显著较短,对照K326与编辑材料无显著差异;过表达植株叶片宽度最短,对照组其次,编辑植株叶片气孔宽度最长,三者之间均存在显著差异。气孔长度与气孔宽度相乘表示气孔的大小。因此,过表达植株的气孔小于对照K326,而编辑植株的气孔大于对照K326。
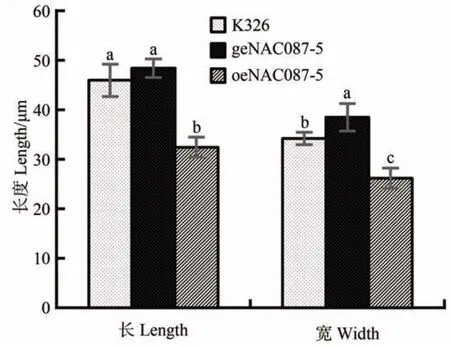
图3 气孔长宽对比图Fig.3 Comparison of pore length and width
2.3 干旱胁迫下酶促反应的变化
由图4-A 可知,在干旱胁迫处理下,3 种材料的SOD 活性都表现出随处理时间延长而升高的趋势,且过表达植株在所有时间段内SOD 活性均显著高于其他植株,在12 h时达到最高,为1 553.96 U·g-1;编辑植株在0、1、3 和6 h 时活性显著高于对照K326,在12 h时与对照K326无显著差异。
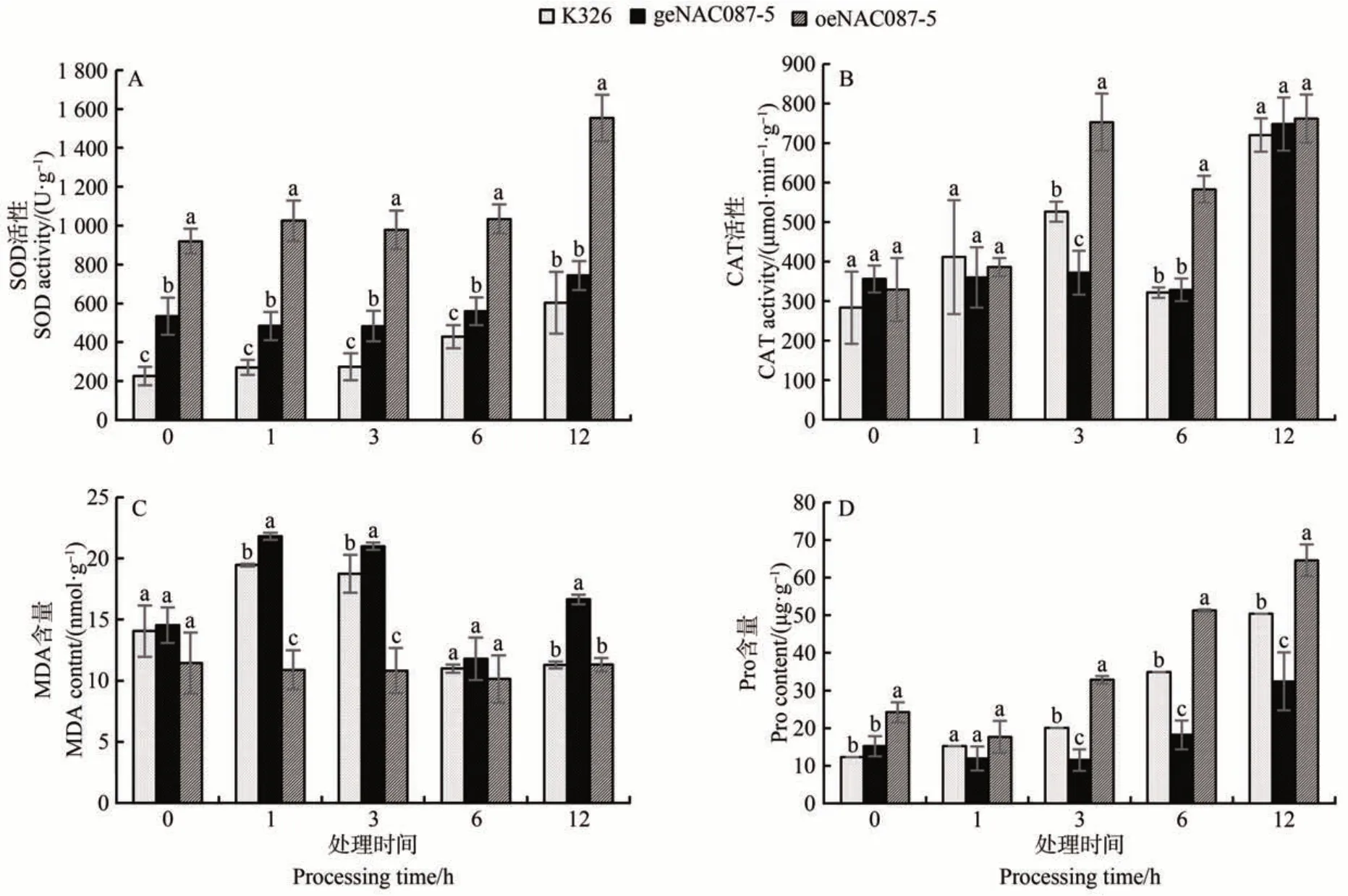
图4 抗氧化酶活性Fig.4 The activities of the antioxidant enzymes
由图4-B可知,在干旱胁迫处理下,3种材料中CAT活性均表现为先增加后减少再增加的趋势。其中,在0和1 h时3种植株中CAT活性无显著差异;在3 h时过表达植株显著高于其他材料,为753.36 μmol·min-1·g-1,编辑植株显著低于其他材料;在6 h时过表达植株活性显著高于其他材料,编辑植株与K326 之间无显著差异;在12 h 时3 种材料无显著差异且均达到最高值。结果表明,过表达植株在短期胁迫下,CAT 活性在3 h 前无显著变化,3~6 h 较对照组显著升高,12 h 时无明显变化。
由图4-C 可知,在干旱胁迫处理下,MDA 含量整体为先上升后下降趋势,在0 h 时3 种材料之间无显著差异;1 和3 h 时过表达材料显著低于其他材料,编辑植株显著高于其他材料;6 h 时3 种材料无显著差异;12 h 时编辑材料与过表达植株显著低于K326。过表达植株在整个处理过程中MDA 含量较为稳定,K326和编辑植株均在1 h时达到最高值。
由图4-D 可知,在干旱胁迫处理下,Pro 含量随时间变化一直升高,在1 h 时过表达植株Pro 含量较对照组不存在显著差异;在0、3、6 和12 h 时过表达植株显著高于K326,编辑植株显著低于K326,说明在胁迫后期NtabNAC087基因对Pro 含量的影响较为明显,提高了植株的抗旱性。
2.4 基因表达量分析
通过qRT-PCR 对NtabNAC087基因表达量进行测定,得到结果如图5 所示。随着处理时间延长,植株中基因的表达量均呈现为先上升后下降的趋势,并且过表达植株在0、1、6 和12 h 时NtabNAC087基因表达量显著高于K326,编辑植株在0、1、3 和6 h 时基因表达量显著低于K326,过表达和对照K326 植株在6 h 时基因表达量达到最高,而编辑植株在3 h时达到最高。
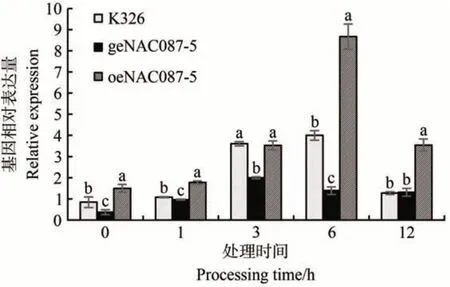
图5 NtabNAC087基因表达量Fig.5 The expression of NtabNAC087 gene
3 讨论
气孔作为控制水分和CO2进出植物的重要途径,气孔的大小、密度和数量等指标可以用于评估植株的抗旱能力[18]。气孔的打开和关闭是由保卫细胞膨胀的变化引起的,保卫细胞膨胀时气孔打开,保卫细胞松弛时气孔关闭,干旱胁迫下会诱导气孔关闭[19]。干旱和低温都会导致植物水分状况失衡,并诱导促进气孔关闭的脱落酸(abscisic acid,ABA)合成。因此,气孔关闭是对干旱和寒冷的适应性反应[20]。在本研究中通过对过表达和基因编辑NtabNAC087基因株系与K326野生型植株的气孔表型进行观察,发现过表达植株相较于编辑和K326 植株中气孔数量较多,气孔较小。贾东峰[21]研究发现苹果中过表达MdNAC1基因会降低气孔大小,提高气孔密度,与本研究结果相符。气孔密度的增加及气孔大小的减小可以保持或改善总气孔的面积,同时也提供了更短的扩散途径,可能会改善气体的交换[22]。水稻中NAC家族OsNAC1基因可以促进叶片气孔关闭,减少水分损失,从而增强过表达植株的耐旱性和耐盐性[23],同样证实了NAC 基因家族可以参与气孔的调节活动。
活性氧(reactive oxygen species,ROS)的产生是植物在正常和胁迫条件下的常见现象,并且在非生物胁迫逆境中,ROS 的产生将可能超出抗氧化防御系统的能力[24]。若植物体内ROS 的动态平衡被打破,植物会通过酶抗氧化系统来维持体内ROS平衡[25]。在干旱胁迫下,SOD和CAT活性上升,从而降低膜脂过氧化损伤程度,提高腺嘌呤核苷三磷酸(adenosine triphosphate,ATP)活性,缓解干旱光合生理的伤害[26]。本研究中SOD活性随处理时间的延长逐渐升高,且过表达植株的SOD 活性与K326存在显著差异。说明干旱胁迫下,过表达植株表现出更好的耐旱性。
ROS的积累,包括作为信号转导分子的H2O2,也可能导致广泛的细胞损伤和光合作用抑制[27]。在干旱胁迫下,植物会发生原生质脱水,引起细胞膜结构的变化及膜上细胞的失活[28]。MDA 是膜脂过氧化的最终产物,可作为氧化脂质损伤标志物[29],其含量可以用来评估植株受到的氧化损伤程度。本研究通过对转基因纯合材料的验证,NtabNAC087基因表达量的检测及干旱胁迫处理条件下烟草植株各种生理指标的测定,验证NtabNAC087基因是否参与干旱胁迫的响应功能。结果表明,CAT活性在3~6 h内过表达植株较其余材料存在显著差异,MDA 含量在1~3 h内差异较为明显,推测NtabNAC087基因可能在干旱胁迫前期对CAT 活性和MDA 含量存在影响,其中3 种材料的Pro 含量、SOD活性存在差异,都为上升趋势且在12 h时差异显著,可能对干旱胁迫后期产生影响。Pro 作为渗透调节物质发挥作用[30],当植株受到的干旱胁迫越为严重,其后期含量就越高。通过3种材料的横向比较,发现面对相同程度的干旱胁迫,过表达植株中Pro 含量明显升高,Pro含量越高则代表其抗旱能力越强。同作为茄科植物的番茄,过表达SlNAC6基因可显示出对干旱胁迫的耐受性大大增强,同时表现出较高的Pro含量和抗氧化酶活性[10],这与本研究结果相吻合,证明了NtabNAC087基因与烟草抗旱性相关,过表达该基因后可提高烟草的抗旱性。
在基因表达量分析结果中,CRISPR/Cas9 技术使NtabNAC087基因表达量下降,与李兆伟等[31]的研究结果相似。本研究还发现NtabNAC087基因表达量的变化较为明显,整体表现为先上升后下降,且在6 h时达到最高,与白戈等[32]研究转录因子在干旱胁迫下的表达量呈先增后减趋势的结果相似。由此推测NtabNAC087基因的抗旱能力可能存在阈值,过表达植株的阈值提高,从而增强了植株的抗旱性。
4 结论
通过克隆烟草中与拟南芥RD26基因同源的NtabNAC087基因,进行过表达及编辑植株的构建,在PEG6000模拟干旱胁迫下研究NtabNAC087基因功能,发现烟草NtabNAC087基因过表达植株通过改变植株叶片气孔、生理指标及相关基因的表达,增强了植株的抗旱性。证明NtabNAC087基因作为转录因子参与干旱胁迫的应答,且过表达NtabNAC087基因可以使植株获得更强的耐旱性。
猜你喜欢
亚热带植物科学(2022年1期)2022-05-17
干旱地区农业研究(2022年1期)2022-01-28
装备制造技术(2019年12期)2019-12-25
四川农业科技(2019年5期)2019-07-01
科技创新与品牌(2019年12期)2019-02-06
浙江中西医结合杂志(2017年2期)2017-01-12
当代化工研究(2016年9期)2016-03-20
浙江柑橘(2016年1期)2016-03-11
中国铸造装备与技术(2015年5期)2015-12-10
声屏世界(2014年6期)2014-02-28
