
小林窗对人工更新紫椴苗木生长和叶片性状的影响
2023-07-05 09:50付瀚萱许冠洲申方圆杨立学
中南林业科技大学学报 2023年5期
付瀚萱,许冠洲,申方圆,杨立学,2
(1. 东北林业大学 a. 林学院;b. 东北乡土树种工程技术研究中心,黑龙江 哈尔滨 150040;2. 森林生态系统可持续经营教育部重点实验室,黑龙江 哈尔滨 150040)
林窗是由于自然或人为因素的干扰而形成的林中空隙,能够改变林地内光照、气温、相对湿度以及土壤性质等环境因子[1-2]。刘海英等[3]研究了受冰雪灾害干扰后形成的不同大小林窗发现,林内的光照强度随林窗面积的增大而增大,且非林窗下土壤(0 ~5 cm)的有机质、全氮、水解氮和有效磷含量均显著高于林窗下土壤。廖睿燕等[4]研究常绿阔叶林中不同林窗和林下光照条件的差异发现,林窗下的光通量是林下的10.43 倍。因此,林窗的形成会对其内部及周围小气候环境产生影响。
林窗不仅改变林地内的环境因子,也会对林窗内的苗木更新产生影响[5]。李佳明等[6]以1年生紫椴Tilia amurensis实生苗为对象,在不同林隙中开展造林试验发现,紫椴苗木在中光区(2 280 ~3 700 lx)和弱光区(400 ~2 250 lx)的成活率相对较高,地径年生长量在中光区显著高于强光区(3 700 ~5 300 lx)和弱光区。朱凯月等[7]比较不同大小林隙下蒙古栎Quercus mongolica和水曲柳Fraxinus mandshurica人工更新幼树生长和形态特征的差异发现,蒙古栎和水曲柳幼树的树高和地径随林隙减小而显著减小,且较大的林隙能促进其生长。光合指标能最直接反映植物对光环境的适应性[8],此外植物叶片的形态和元素含量也会产生可塑性变化以提高对光能的吸收和利用[9]。刘辉等[10]研究马尾松Pinus massoniana人工林窗中油樟Cinnamomum longepaniculatum幼苗叶片形态和光合生理特征的差异发现,在一定范围内,油樟幼苗的最大净光合速率和叶片氮含量随林窗面积增大而显著增加。张培等[11]对不同林窗下格木Erythrophleum fordii幼树叶片形态特征的研究发现,其平均单叶面积随林窗面积增大而显著减小,比叶面积则显著增加。可见,林窗改变了林分内的光、热、水和肥等条件,对林下苗木更新和生长具有重要的驱动力。以往的研究大多采用林窗长轴与边缘木平均高的比值(比值在0.25 ~0.5范围为小林窗、0.5 ~1.0 为中林窗、大于1.0 为大林窗)来衡量林窗的大小[12]。苗木年龄越小对光环境的变化越敏感[3-4],而以往研究涉及到的林窗面积较大[2,10],关于小林窗对苗木生长、成活的影响以及作用机理尚不明确。
紫椴是东北地区珍贵的阔叶树种,同时也是重要的用材、蜜源、药用植物以及城市的绿化树种。由于过度采伐,天然林中紫椴的数量和质量急剧下降[13]。紫椴苗木虽然对光照环境适应性较广,但研究显示其在全光环境下人工更新效果并不好[14],鉴于此,在林窗下开展人工更新紫椴造林试验具有重要意义。目前,关于紫椴苗木的研究主要集中在人工遮阴对其形态和生理等方面的影响[15]以及不同立地条件对苗木的生长及成活的影响[6],而针对不同小林窗下紫椴更新苗生长和成活的作用机理尚不明确。本研究以黑龙江省尚志市小九林场天然次生林内人工更新的紫椴(2年生)苗木为对象,研究小林窗(面积:6 ~45 m2)对苗木的生长(树高、地径)、存活(存活率)、光合能力(Pn、Tr、Ci、Gs、Chlt)以及叶片形态特征(叶面积、比叶面积)、碳氮含量的影响,旨在阐明紫椴苗木对小林窗的适应特点,为紫椴在小林窗及林冠下人工更新提供理论依据。
1 材料与方法
1.1 研究地概况
研究地点位于黑龙江省尚志市国有林场管理局小九林场(127°38′ ~127°51′E,45°11′ ~45°25′N),本研究样地设置为海拔、坡度、土壤类型相同或相近的天然次生林内。该地区属大陆中温带季风气候,平均海拔420 m,平均坡度为26°,年降水量550 ~740 mm,年平均气温2.3 ℃,结冰期长达5 个月,无霜期120 d,日照时数2 561 h;土壤以暗棕壤为主。该区顶级植被原为红松阔叶混交林,后经历史性采伐,现逐渐形成阔叶混交林。主要乔木树种有紫椴、五角枫Acer mono、青楷槭Acer tegmentosum、蒙古栎、白桦Betula platyphylla、黑桦Betula dahurica、胡桃楸Juglans mandshurica、 黄檗Phellodendron amurense、 水曲柳等,主要灌木有平榛Corylus heterophylla、暴马丁香Syringa reticulatasubsp.amurensis、忍冬Lonicera japonica、绣线菊Spiraea salicifolia等,草本植物有苔草Carex rigescens、蚊子草Filipendula palmata、 白花碎米荠Cardamine leucantha、旱芹Apium graveolens等。
1.2 研究方法
1.2.1 样地设置
2020年4月,调查了研究地点中的林窗大小,发现样地内天然形成的林窗面积较小,在9 个林窗和3 个林冠下,以1 m×1 m 的株行距栽植2年生紫椴苗木,其生长状况相对一致,没有明显病虫害,共栽植486 株。2020年9月,林窗面积均按照椭圆形面积公式S=πLW/4 计算(π 为圆周率,L为林窗的长轴长度,W为林窗的短轴长度)[1],得到林窗面积范围为6 ~45 m2。其中,面积大于30 m2的林窗为G1(31、36、45 m2),介于20 ~30 m2之间的为G2(21、24、28 m2),小于20 m2的为G3(6、12、15 m2),林冠下为对照样地,共计12 块样地。按林窗长轴与边缘木平均高的比值来衡量林窗的大小[12],确定本研究所调查到的林窗均属于小林窗。
1.2.2 样品采集与测定
2020年9月,在各样地分别设置3 个小样方,取0 ~20 cm 的土壤,充分混匀后做好标记带回实验室并测定其土壤理化性质(表1)。2021年6—7月,测定各样地所有存活苗木的树高和地径,统计紫椴苗木的存活株数并计算存活率。使用照度计测量各样地的光照强度,在各林窗中心的东、南、西、北4 个方向分别读取数值,取平均值作为该样地的光照强度(表1)。每个样地根据树高和地径分别确定3 株平均木作为标准木,用Li-6400 便携式光合仪于晴朗天气的9:00—11:00 测定每株标准木叶片的气体交换参数(Pn、Ci、Tr、Gs),对其同一部位的成熟叶片进行测定,重复3 次。每株标准木取20 片健康叶片,将叶片做好标记带回实验室并测定其形态、叶绿素和碳氮含量。
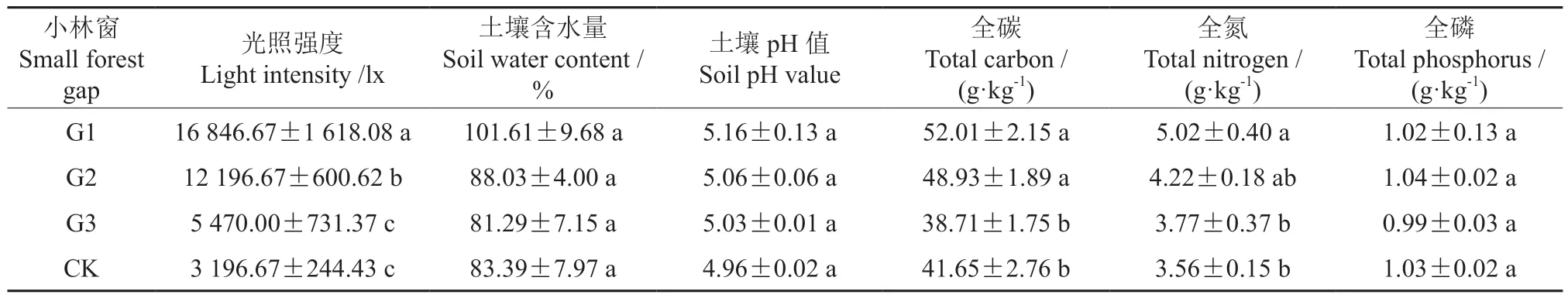
表1 小林窗和林冠下林地的光照强度和土壤养分†Table 1 Differences in the light intensity and soil nutrients in forests with small forest gaps and closed canopy
1.2.3 数据处理
采用Excel(Microsoft, 2010)和SPSS(SPSS公司,19.0)软件对数据进行整理和分析;采用单因素方差分析(One-way ANOVA)对紫椴苗木和土壤各指标进行分析;采用LSD 检验不同处理之间的差异(α=0.05),所有数据为平均值±标准误;相关分析采用Pearson 相关法;使用SigmaPlot(SYSTAT 公司,12.0)软件作图。
2 结果与分析
2.1 不同小林窗和林冠对紫椴苗木生长和存活率的影响
不同林窗对紫椴苗木的树高和存活率影响显著,对地径无显著影响。随着林窗面积的减小,紫椴苗木的树高呈现出下降的趋势,其中,G1(63.16±0.90)cm 和G2(62.20±1.34)cm 显著高于G3(51.27±0.89)cm 和CK(50.52±1.21)cm(P<0.05),G1 与G2 以及G3 与CK 间均无显著差异(P>0.05,图1);紫椴苗木的存活率在各林窗下均显著高于CK(P<0.05),但在各林窗间无显著差异(P>0.05,图1)。

图1 不同小林窗和林冠下紫椴苗木生长和存活率Fig. 1 Growth and survival rate of Tilia amurensis seedlings in different small forest gaps and closed canopy
2.2 不同小林窗和林冠对紫椴叶片形态特征和碳氮含量的影响
不同林窗对紫椴叶片面积、比叶面积和叶片氮含量影响显著,对叶片碳含量无显著影响。紫椴叶片的叶面积、比叶面积和氮含量随林窗面积的减小呈现明显的上升趋势。紫椴的叶片面积在G1(25.94±0.79)cm2和G2(26.44±0.64)cm2下显著低于CK(31.08±1.88)cm2(P<0.05),G3(28.10±1.41)cm2与其他3 个林窗环境以及G1 与G2 之间均无显著差异(P>0.05,图2);紫椴的比叶面积在G1(167.15±8.80)cm2·g-1下显著低于CK(191.23±7.51)cm2·g-1(P<0.05),G2(172.51±6.16)cm2·g-1及G3(184.82±5.59)cm2·g-1与其他3 个林窗环境之间均无显著差异(P>0.05,图2);紫椴叶片的氮含量在G1(3.17±0.19)g·kg-1下显著低于G3(3.73±0.11)g·kg-1和CK(3.91±0.11)g·kg-1(P<0.05),G2(3.53±0.14)g·kg-1与其他3 个林窗环境以及G3 与CK之间均无显著差异(P>0.05,图2)。

图2 不同小林窗和林冠下紫椴叶片形态特征和碳氮含量Fig. 2 Morphological characteristics and carbon and nitrogen contents of Tilia amurensis leaves in different small forest gaps and closed canopy
2.3 不同小林窗和林冠对紫椴叶片气体交换参数和叶绿素含量的影响
不同林窗对紫椴Pn、Tr、Ci、Gs、Chlt 和Chla/b 影响显著。紫椴的Tr、Gs和Chla/b 随林窗面积的减小呈明显下降的趋势,Ci和Chlt 随林窗面积的减小呈明显的上升趋势。紫椴的Pn和Tr在G1 和G2 下显著高于G3 和CK(P<0.05),G1与G2 以及G3 与CK 间均无显著差异(P>0.05);紫椴的Ci在G1 和G2 下显著低于CK(P<0.05),G3 与其他3 个林窗环境以及G1 和G2 间均无显著差异(P>0.05);紫椴的Gs在G1 下显著高于G3 和CK(P<0.05),G2 与其他3 个林窗环境以及G3 和CK 间均无显著差异(P>0.05);紫椴的Chlt 在G1 下显著低于G2、G3 和CK(P<0.05),G2、G3 和CK 间无显著差异(P>0.05);紫椴的Chla/b 在G1 下显著高于CK(P<0.05),G2 及G3 与其他3 个林窗环境之间均无显著差异(P>0.05,图3)。

图3 不同小林窗和林冠下紫椴叶片气体交换参数和叶绿素含量Fig. 3 Gas exchange parameters and chlorophyll content of Tilia amurensis leaves in different small forest gaps and closed canopy
2.4 人工更新紫椴苗木叶片指标的相关性分析
Pearson 相关性分析表明,光照强度与紫椴苗木的树高、存活率以及树高与存活率呈极显著正相关关系(表2)。光照强度与紫椴叶片的比叶面积和氮含量呈极显著负相关关系;紫椴叶片的比叶面积与氮含量呈极显著正相关关系(表3)。光照强度与Pn、Tr和Gs呈极显著正相关关系,与Ci和Chlt 呈显著负相关关系;Pn与Tr和Gs呈显著正相关关系,与Chlt 呈显著负相关关系;Tr与Gs呈极显著正相关关系,与Ci和Chlt 呈显著负相关关系;Gs与Chlt 呈极显著负相关关系(表4)。

表2 光强、紫椴生长和存活率的相关性分析†Table 2 Correlation analysis of light intensity and the growth and survival rate of Tilia amurensis

表3 光强、紫椴叶片形态和碳氮含量的相关性分析Table 3 Correlation analysis of light intensity, leaf morphology and carbon and nitrogen content of Tilia amurensis
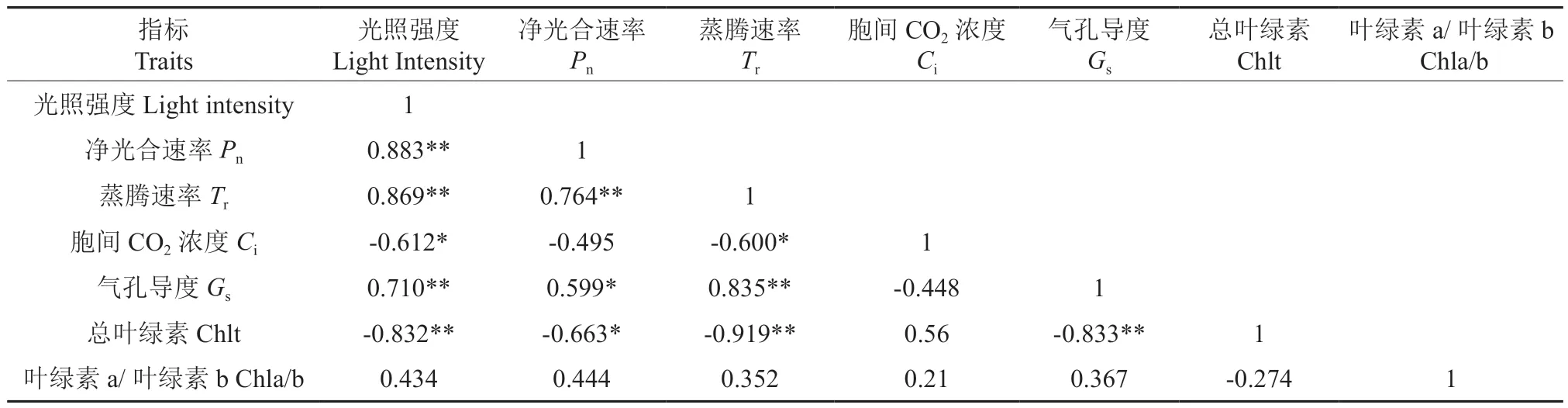
表4 光强、紫椴叶片气体交换参数和叶绿素含量的相关性分析Table 4 Correlation analysis of light intensity, gas exchange parameters and chlorophyll content of Tilia amurensis leaves
3 结论与讨论
3.1 结 论
本研究通过对天然次生林小林窗下人工更新紫椴(2年生)苗木生长、光合能力以及叶片形态和碳氮含量的测定,确定适合紫椴苗木生长和存活的林窗面积,并分析紫椴苗木在不同林窗和林冠下的适应特点,得出主要结论如下:
1)林窗面积的变化导致紫椴苗木的树高和存活率产生差异,其树高和存活率在林冠下(CK)显著低于面积大于20 m2的林窗(G1 和G2)。且在林冠(CK)和面积小于20 m2的林窗(G3)下,紫椴苗木的净光合速率显著低于面积大于20 m2的林窗(G1 和G2)。这表明在面积大于20 m2的林窗下,紫椴苗木光合作用更强,光合产物较多,具有较强的生存竞争力,而当林窗面积小于20 m2或林冠下,紫椴更新苗无法获取足够的阳光,生长受到抑制。因此,紫椴苗木更适合在面积大于20 m2的林窗(G1 和G2)下人工更新。
2)紫椴苗木叶片的形态、化学和生理指标也会随林窗面积的变化而变化。在形态上,紫椴苗木的叶片面积、比叶面积均随着林窗面积的减小而显著增加,这表明苗木通过增大叶面积和比叶面积来增加光能的捕获。在元素含量上,紫椴苗木叶片氮含量在林冠下最高,且氮含量与光照强度呈极显著负相关关系,表明苗木叶片的氮含量与植物的光合作用以及生长状况紧密联系。紫椴苗木的蒸腾速率、气孔导度和Chla/b 随林窗面积的减小而显著降低,而胞间CO2浓度和叶绿素含量显著增加。因此,紫椴苗木通过改变其形态、化学及生理特征来适应弱光环境,表现出一定的耐阴性。
3)植物的年龄越小对光环境的变化越敏感,本研究结果表明紫椴苗木在面积大于20 m2的林窗(G1 和G2)下,生长状况(树高、存活率及光合能力)要显著优于面积小于20 m2的林窗(G3)和林冠下(CK)。即使在小林窗下,林窗面积依然会对紫椴苗木的生长产生影响。因此,在林木培育管理的过程中,要重视小林窗对苗木生长的影响。
3.2 讨 论
3.2.1 不同林窗对紫椴生长和存活率的影响
环境因子中,光照条件对苗木生长和存活的影响较大[8]。树高、地径和存活率可以最直观地反映苗木生长状况,本研究中,随着林窗面积(大于30 m2、20 ~30 m2、小于20 m2及林冠)的变化,紫椴苗木的树高和存活率呈现出一定的差异,这与以往的研究结果类似。例如,张宇阳等[16]比较梓叶槭Acer catalpifolium幼树在无遮阴、林缘、林隙以及林下生长的差异发现,其株高在无遮阴和林缘下显著高于林下和林隙。叶飞英等[17]研究不同光强下伯乐树Bretschneidera sinensis幼苗的生长状况发现,随着光照强度的减弱,苗木的苗高逐渐降低。这可能是因为不同林窗下,苗木所处的生态条件不同,导致其光合产物的合成和累积有差异[18],从而影响了紫椴苗木的生长和存活。
3.2.2 不同林窗对人工更新紫椴苗木叶片形态和碳氮含量的影响
林窗可以改变林地内的光照条件,苗木叶片的形态特征在不同的光照环境下表现出不同的适应性变化[19]。本研究结果表明,紫椴叶片面积和比叶面积在林冠下最大,这与以往的研究结果一致。例如,张兰等[20]研究盆栽辽东栎Quercus wutaishanica幼苗在不同光照环境下叶片的形态特征发现,其苗木比叶面积随光照强度的减弱而显著增加。刘青青等[21]对比不同光照强度下杉木Cunninghamia lanceolata幼苗叶片的形态特征发现,随着光照强度的减弱,杉木幼苗叶片的比叶面积显著增加。有研究表明,苗木叶片的叶面积、比叶面积增加,有利于其在弱光环境下更有效地利用光照资源[19]。在弱光环境下,紫椴叶片可能通过变大变薄的方式来增加光能的捕获,从而减少因光照强度过低对光合速率的限制。紫椴叶片面积以及比叶面积在G1、G2 以及G3 下差异均不显著(P>0.05),这可能是3 种林窗面积的差异较小,使得紫椴苗木在叶片形态上的变化并不明显。
叶片中氮含量对植物的生长以及光合作用具有重要意义[22]。本研究中,叶片氮含量随林窗面积的减小而显著增加,叶片氮含量与光照强度呈显著负相关。这可能是叶片中的氮元素是叶绿素和光合作用中各种酶的组成元素,从而使叶片氮和叶绿素含量与光照强度的关系紧密相关。
3.2.3 不同林窗对人工更新紫椴苗木光合作用的影响
净光合速率是最直接体现植物光合效率的指标,不仅与叶绿素含量、叶面积大小等自身因素密切相关,还受到光照强度等外界环境因子的影响[8,23]。相关研究表明,植物对光环境的适应是沿着有利于其光合作用的方向发展,适当地增加光照会提高植物的净光合速率[24]。本研究表明,随着林窗面积的减小,紫椴的净光合速率总体呈下降的趋势,这与以往的研究结果类似。例如,罗光宇等[25]研究濒危植物长序榆Ulmus elongata在不同光照强度下的光合特性发现,植株的净光合速率日均值随光照强度的减弱而显著降低。这表明林冠下可能发生了光亏缺,紫椴苗木无法获得正常生长发育所必需的光资源,而林窗内光照条件有利于其光合产物的积累,为紫椴苗木在林地生存中保持较强竞争力提供了条件。
相关研究表明,植物的净光合速率与气孔导度紧密相关[25]。本研究结果表明,紫椴的气孔导度随林窗面积的减小而减小,且气孔导度与净光合速率呈显著正相关关系,这与以往的研究结果类似。例如,盛海燕等[24]以延胡索Corydalis yanhusuo为研究对象,比较其苗木在不同光环境下光合指标的差异发现,气孔导度和净光合速率均随光照强度的减弱而降低。陈超等[23]研究不同光照强度下濒危植物景宁木兰Magnolia sinostellata光合能力的季节变化发现,在春季和夏季,其幼苗的净光合速率和气孔导度均随光照强度的下降而减小。这表明,植物与外界通过气孔进行H2O和CO2等气体交换,弱光环境下植物的气孔导度较低,气体交换能力较差,从而导致净光合速率降低。
叶绿素是光合作用的物质基础,在光合作用中起吸收、传递、转化光能的作用,其含量与苗木的光合能力密切相关[20]。相关研究表明,弱光环境下的植物可以通过增加叶绿素含量来提高光能的吸收和传递效率。本研究的结果表明,紫椴叶片的叶绿素含量随着林窗面积的减小显著增加,这与以往的研究结果类似。张玲等[26]比较遮阴条件下黄檗幼苗生理指标间的差异发现,其幼苗总叶绿素含量在重度遮阴下显著高于全光、轻度和中度遮阴。陈菊艳等[27]模拟不同光照条件,探究野鸦椿Euscaphis japonica幼苗光合生理特征的差异发现,其幼苗叶绿素总量随着遮光程度的加大总体呈明显上升的趋势。这表明植物为适应弱光环境采取增加叶绿素含量以最大限度提高采光能力的策略。
Chla/b 降低通常被认为是植物为适应弱光环境而采取的一种调控策略[24,27]。本研究结果表明随着林窗面积的减小,紫椴叶片的Chla/b 逐渐减小,这与以往的研究结果类似。例如,黄河腾等[19]研究不同遮阴环境下木奶果Baccaurea ramiflora幼苗生理特征的差异发现,随着遮阴程度的加大,其幼苗的Chla/b 逐渐降低。李雨霏等[28]比较不同光环境下青钱柳Cyclocarya paliurus苗期生理指标的差异发现,其幼苗的Chla/b 呈明显的下降趋势。
受研究地点的限制,本研究并未涉及大于45 m2的小林窗,具有一定的局限性。且紫椴苗木需光性随着紫椴的生长而发生变化,随着紫椴苗木年龄的增加,小林窗对紫椴苗木的生长及叶片性状是否仍然产生影响,需要进一步研究。未来将探讨小林窗对长期作用下的紫椴更新苗的生长状况、干形以及根系性状的影响,为紫椴苗木的更新提供理论依据。
猜你喜欢
意林(2023年4期)2023-04-28
西南林业大学学报(2022年5期)2022-10-17
Chinese Physics B(2021年9期)2021-09-28
意林(2021年11期)2021-09-10
环保科技(2020年2期)2020-05-21
农业机械学报(2020年1期)2020-03-13
浙江农林大学学报(2019年6期)2019-11-13
意林(2019年16期)2019-09-04
意林(2018年7期)2018-05-03
西部林业科学(2018年4期)2018-02-14
