
枯草芽孢杆菌发酵对豆类粗多糖结构与抗氧化活性的影响
2023-08-25 07:16屈雅宁许梦粤唐双庆陈禅友王亚珍王红波
食品工业科技 2023年17期
屈雅宁,许梦粤,唐双庆,刘 琴,陈禅友,王亚珍,王红波,,,
(1.江汉大学生命科学学院,食品营养与安全研究中心,湖北武汉 430056;2.湖北省豆类(蔬菜)植物工程技术研究中心,湖北武汉 430056;3.湖北省健康代糖产品企校联合创新中心,湖北武汉 430056)
豆类营养价值高,口感良好,是人们喜食的优质食品资源。豆类除含有丰富的蛋白质、脂肪和维生素等营养物质外,还含有多糖、黄酮和皂苷等生物活性成分,使豆类具有抗氧化、抑菌、降血糖和降血压等功能,从而更加受到关注[1-2]。多糖的生物学功能是食品营养领域的研究热点。YANG 等[3]分离纯化了绿豆中的多糖,其表现出良好的抗氧化活性。BAI等[4]研究了6 种豆科植物粗多糖的化学组成,结果表明较高含量的粗多糖和具有特定结构的粗多糖对降血糖具有一定的作用。
发酵是一种传统的食品生物加工技术,通过微生物发酵作用能改善其风味,也能显著提高其营养品质和生物学功能。国内对传统发酵豆制品的研究大多集中于产品营养与风味方面,目前对发酵豆制品发酵过程中产生新物质的种类、结构和功能挖掘不够。JHAN 等[5]研究发现枯草芽孢杆菌发酵能提高红豆食品的抗氧化能力。邰佳等[6]利用枯草芽孢杆菌和纤维素降解菌混合菌种接种玉米须,固态发酵后玉米须中的总黄酮含量提高了38.89%,多糖含量提高了32.69%。目前已有研究报道大豆水溶性多糖具有抗肿瘤、抑菌、抗氧化等活性,在食品工业中应用广泛。黑豆多糖、绿豆多糖和豇豆多糖等杂豆多糖的部分结构信息已有一些研究者解析,而关于其他杂豆类多糖的研究偏少,且有关微生物发酵对豆类多糖结构影响的研究报道较少。
本研究选取了大豆、豇豆、红豆、绿豆、扁豆、豌豆、蚕豆和菜豆8 种常见的食用豆子为研究对象,以枯草芽孢杆菌为发酵菌株,采用固态发酵方式,比较研究了8 种豆子发酵前后粗多糖的化学组成、分子量分布、红外光谱结构以及抗氧化活性能力的变化,为发酵豆类功能食品的开发提供理论基础。
1 材料与方法
1.1 材料与仪器
大豆(MD-065,自选材料)、豇豆(鄂豇豆7 号)、红豆(红珍珠)、绿豆(绿宝石)、扁豆(德扁3 号)、豌豆(汉豌1 号)、蚕豆(CD-035,自选材料)、菜豆(SJ-0052,自选材料) 均由湖北省食用豆类植物自然科技资源中心提供;菌种枯草芽孢杆菌(Bacillus subtilissubsp.Subtilis,ATCC 6051) 保藏于湖北省豆类(蔬菜)植物工程技术研究中心;LB 培养基 实验室自制(蛋白胨1 g,酵母粉0.5 g,NaCl 1 g,蒸馏水100 mL,琼脂2 g);2,4,6-三(2-吡啶基)-1,3,5-三嗪(2,4,6-Tri (2-pyridine)-1,3,5-triazine,TPTZ) 索莱宝公司;1,1-二苯基-2-三硝基苯肼(1,1-Diphenyl-2-picrylhydrazyl,DPPH)、2,2′-联氨-双-3-乙基苯并噻唑啉-6-磺酸(2,2'-Azinobis-3-ethylbanzthiazo-line-6-sulfonate,ABTS)、铁离子还原能力工作液(Ferric ion reducing antioxidant power,FRAP) 麦克林公司;三氯乙酸(Trichloroacetic acid,TCA)、牛血清白蛋白(Bovine albumin,BSA)、磷酸缓冲盐溶液(Phosphate buffered saline,PBS)、葡萄糖、蒽酮乙酸乙酯、半乳糖醛酸、间羟基联苯、四硼酸钠/硫酸、右旋糖酐等试剂 国药集团化学试剂有限公司。
1515 高效液相色谱仪 美国Waters;OHpak SB-803 HQ、Ohpak SB-804 HQ、Ohpak SB-805 HQ聚合物基质水溶性SEC(GFC)色谱柱 日本Shodex;FA124C 分析天平 上海力辰科技;TGL-16A 离心机 长沙平凡仪器;NICOLET iS50R 傅里叶变换显微红外光谱仪 美国Thermo Scientific;JP-020 超声波清洗机 深圳市洁盟清洗设备有限公司;SPX-150B-Z 型生化培养箱 上海博讯实业有限公司医疗设备厂;SMF01 磨粉机 苏泊尔电器公司;FreeZone 6 Plus 冷冻干燥机 美国LABCONCO 公司;1510全波长酶标仪 Thermo Fisher 公司。
1.2 实验方法
1.2.1 菌株活化与扩大培养 将枯草芽孢杆菌划线接种至LB 培养基,于37 ℃下培养2 d,挑取枯草芽孢杆菌单菌落于LB 液体培养基中,37 ℃摇床培养24 h,得到种子液。
1.2.2 固态发酵与冷冻干燥 参考LI 等[7]和BJMA等[8]的实验方法。将8 种豆子用磨粉机磨碎(直径小于1 mm),分别称取12 g 置于锥形瓶内,121 ℃灭菌15 min,冷却,以5%接种量将菌株种子液接种于豆粉,置于37 ℃培养箱发酵培养7 d。发酵结束后,将发酵样品置于冷冻干燥机,真空度0.040 MBar,温度-85 ℃,冷冻干燥12 h,将冻干后的发酵样品于-20 ℃冰箱储存备用。
1.2.3 粗多糖的提取
1.2.3.1 样品脱脂 参考李安琪等[9]的方法对样品进行预处理。将未发酵和发酵后豆粉,分别取6 g 于烧杯,按料液比1:10(g/mL)加入95%的乙醇,室温摇床震荡6 h,6000 r/min 离心10 min,收集沉淀,于60 ℃烘箱烘干,密封保存备用。
1.2.3.2 粗多糖提取 取脱脂后豆粉,按料液比1:20(g/mL)添加蒸馏水,95 ℃条件下提取6 h,6000 r/min 离心10 min,收集上清液。
1.2.3.3 粗多糖除蛋白 多糖提取上清液中加入等体积3% TCA 溶液于4 ℃冰箱静置过夜[10-11],8000 r/min 离心10 min 除去蛋白质,取上清液。在上清液中加入等体积无水乙醇,4 ℃静置过夜,8000 r/min 离心10 min,收集沉淀,冷冻干燥得粗多糖。
1.2.3.4 粗多糖得率的计算 称量冷冻干燥后的粗多糖,按以下公式计算发酵前和发酵后豆粉粗多糖的得率。
1.2.4 粗多糖组成测定
1.2.4.1 总糖含量测定 参考刘艺珠等[12]的方法,量取标准(0.1 mg/mL)葡萄糖溶液0、0.2、0.4、0.6、0.8、1.0 mL,用蒸馏水补齐至2 mL,加入蒽酮乙酸乙酯0.5 mL,再加入5 mL 浓硫酸,沸水浴5 min 后冷却,在620 nm 波长下测定吸光度,以葡萄糖浓度为横坐标(mg/mL),吸光值为纵坐标,得到回归方程y=3.8829x+0.0494,R2=0.9992。取样品20 mg 溶于40 mL 水中配成0.5 mg/mL 的样品溶液,记录吸光度,计算总糖的含量(%)。
式中,C 为样品质量浓度,mg/mL;N 为稀释倍数;V 为样品定容体积,mL;m 为样品质量,mg。
1.2.4.2 糖醛酸含量测定 以半乳糖醛酸为标准品,采用间羟基联苯比色法[13]测定各粗多糖样品中糖醛酸含量。取半乳糖醛酸标准溶液0.5 mL 定容至10 mL,取1 mL 该定容溶液,冰水浴,缓慢加入四硼酸钠/硫酸溶液6 mL,涡旋混匀,沸水浴5 min 后冷却,加0.1 mL 间羟基联苯溶液,充分混匀振荡5 min,超声处理除气泡,在200~800 nm 波长内扫描,确定最大吸收波长,以糖醛酸浓度(mg/mL)为横坐标,吸光值为纵坐标,得回归方程y=2.6536x+0.072,R2=0.9918。分别取样品20 mg,用蒸馏水配制成浓度为0.5 mg/mL的样品溶液。取样品溶液1 mL,后续同上述操作,计算糖醛酸含量(%)。
式中,C 为样品质量浓度,mg/mL;V 为样品定容体积,mL;m 为样品质量,mg。
1.2.4.3 蛋白质含量测定 以牛血清白蛋白(BSA)作为标准品[14],采用考马斯亮蓝法测定各豆粉粗多糖样品中的蛋白质含量,记录各反应液在595 nm 波长下的吸光值,以蛋白质浓度(mg/mL)为横坐标,吸光值为纵坐标,得回归方程y=2.4714x+0.2221,R2=0.994。取样品20 mg 溶于40 mL 水中配成0.5 mg/mL的样品溶液,后续操作步骤同标准溶液,根据标准曲线计算样品中蛋白质含量(%)。
式中,C 为样品质量浓度,mg/mL;N 为稀释倍数;V 为样品定容体积,mL;m 为样品质量,mg。
1.2.5 分子量分布测定 称量不同分子量的右旋糖酐标准品,使用高效凝胶渗透色谱(HPGPC)串联柱来进行检测,得校正曲线。称取样品5 mg,向样品中加入0.05 mol/L NaCl 溶液1 mL,配制成5 mg/mL供试样品溶液,8000 r/min 离心10 min,取上清液用0.22 μm 的微孔滤膜过滤,然后将样品转置于2 mL进样瓶中,进行多糖分子量分布检测[15]。色谱条件:色谱柱:聚合物基质水溶性SEC(GFC)色谱柱(8×300 mm)3 根串联;流动相:0.05 mol/L NaCl 溶液;流速:0.65 mL/min;柱温:40 °C;进样量:30 μL。
1.2.6 红外光谱分析 称取各干燥的粗多糖样品1~2 mg 于在研钵中,分别加入KBr 粉末200 mg 研磨均匀,压片,使用傅里叶变换显微红外光谱仪扫描样品,波数范围4000~400 cm-1,绘制红外光谱图[16]。
1.2.7 粗多糖抗氧化活性测定
1.2.7.1 DPPH 自由基清除活性测定 参考辛玥等[17]的DPPH 测定方法。制备0.1 mmol/L DPPH 溶液,分别取0.2 mL 粗多糖样品溶液和0.2 mL 0.1 mmol/L DPPH 溶液混合均匀,避光反应30 min 后,在517 nm波长处测量吸光值,结果按公式(5)计算DPPH 自由基清除率:
式中:A0为0.2 mL H2O 加0.2 mL DPPH 的吸光度;A1为0.2 mL 样品溶液加0.2 mL 50%乙醇的吸光度;A2为0.2 mL 样品溶液加0.2 mL DPPH 的吸光度。
1.2.7.2 ABTS+自由基清除活性测定 将7.4 mol/L ABTS 溶液和3.8 mmol/L 硫酸钾溶液等体积混合,避光反应12 h 得到母液。在734 nm 下将母液用PBS(pH7.4)缓冲溶液调整吸光度到0.70±0.02,得到反应溶液[18]。将25 μL 粗多糖样品溶液和250 μL的反应溶液混匀,静置15 min,用酶标仪在734 nm下测量吸光值。结果按公式(6)计算ABTS+自由基清除率:
式中:A0为25 μL H2O 加250 μL 反应液的吸光度;A1为25 μL 样品溶液加250 μL PBS 缓冲液的吸光度;A2为25 μL 样品溶液加250 μL 反应液的吸光度。
1.2.7.3 铁离子还原能力测定(FRAP) 将300 mmol/L醋酸盐缓冲液(pH3.5)、10 mmol/L TPTZ 溶液和20 mmol/L 氯化铁溶液以10:1:1 体积混合,制成FRAP 工作液。取900 μL FRAP 工作液与100 μL 粗多糖样品溶液混合,于37 ℃下反应5 min,用酶标仪在595 nm 波长下测定吸光度值[19]。相同条件下以FeSO4为参照标准,作浓度变化曲线。计算粗多糖样品的铁离子还原能力(μmol/L)。
1.3 数据处理
本实验数据均重复测定三次。实验数据以平均值±标准偏差表示。运用Microsoft Excel 2010 整理数据,采用SPSS (version 27.0)进行单因素方差分析,P<0.05 表明具有显著性差异。GrapPad 软件计算IC50值,Origin Pro 2021 绘制相关数据图。
2 结果与分析
2.1 发酵前后豆类中粗多糖得率与组成比较分析
发酵前后8 种豆粉粗多糖组分变化见表1。未发酵豆粉粗多糖得率显著高于发酵后豆粉(P<0.05),由于豆子中蛋白质含量较高(蛋白质含量40%左右)[20],三氯乙酸(TCA)未将蛋白质除尽,未发酵豆粉粗多糖中含有少量蛋白质。未发酵豆粉粗多糖总糖含量在3.35%~35.63%之间,发酵后豆粉粗多糖中总糖含量在16.61%~77.11%之间。发酵后豆粉粗多糖的总糖含量显著增加(P<0.05),且蚕豆和豇豆增加最显著(P<0.05),分别提高至58.84%和77.11%。发酵后豆粉粗多糖中蛋白质被除尽,提高了多糖的纯度。未发酵的蚕豆、绿豆、红豆和豇豆粗多糖中含有少量的糖醛酸,发酵后8 种豆粉粗多糖中的糖醛酸含量均增加,且豌豆增加最显著(P<0.05),糖醛酸含量达16.63%。TCA 方法脱蛋白处理对提高粗多糖中多糖含量具有显著的效果[21],豆粉发酵过程中,微生物消耗了豆粉中的部分蛋白质,使得发酵后的豆粉粗多糖样品均未发现蛋白质,微生物发酵联合TCA 处理是去除豆粉粗多糖中蛋白质的理想方法。胡彦波等[22]研究结果表明样品中少量蛋白质对后续粗多糖抗氧化实验结果无明显影响,因此,粗多糖样品无需进一步的脱蛋白处理。本研究结果表明枯草芽孢杆菌发酵能显著提高豆粉粗多糖的纯度、增加粗多糖中糖醛酸的含量。豆粉中含有纤维素,纤维素的细胞壁被枯草芽孢杆菌的酶系代谢,促进了多糖的释放[6]。因此,经微生物发酵后豆粉粗多糖的含量均有提高。
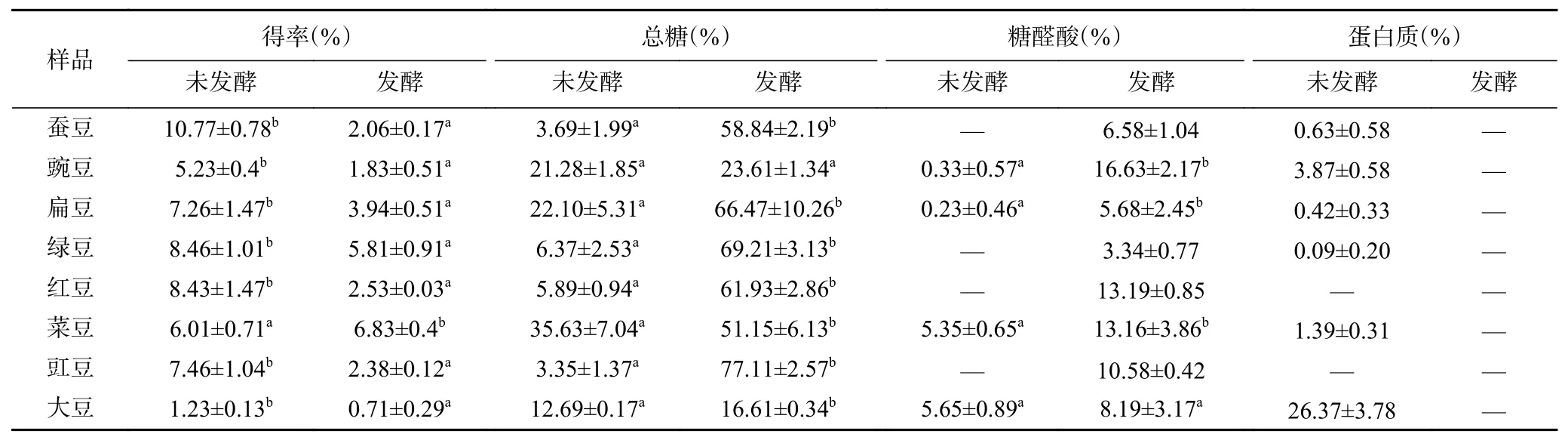
表1 8 种豆粉发酵前后粗多糖得率与组成比较分析Table 1 Comparison and analysis of the yield and composition of eight kinds of legume crude polysaccharides before and after fermentation
2.2 发酵前后豆类中粗多糖分子量比较分析
采用HPGPC 法比较分析8 种豆粉发酵前后粗多糖的分子量,研究结果如图1 所示。发酵前大豆、豇豆、红豆、绿豆、扁豆、豌豆、蚕豆和菜豆均出现1 个明显的特征吸收峰,其分子量分别为0.45、6.10、4.80、14.80、4.17、1.80、4.14 和2.25 kDa;而发酵后豆粉粗多糖特征峰分子量分别为45.39、43.60、30.90、28.10、18.00、56.56、40.60 和36.85 kDa。结果表明豆粉粗多糖分子量分布不均一,且发酵后粗多糖分子量均增加。粗多糖分子量增加可能是由于多糖分子中含有大量的羟基,羟基伸缩振动,分子间作用力相互聚集,尤其是酸性多糖容易形成不同程度的聚集体,导致多糖分子量增加[23];也可能是由于枯草芽孢杆菌分泌的多糖聚合酶将小分子多糖酶促聚合生成大分子的多糖化合物[24]。通过微生物发酵后大豆粗多糖特征分子量明显,从0.45 kDa 增加到45.39 kDa。由于粗多糖分子量的差异,会使其具有不同的生物活性[11]。因此,枯草芽孢杆菌发酵增加了豆粉粗多糖的分子量,枯草芽孢杆菌发酵或许是制备新的豆类多糖的理想方法。
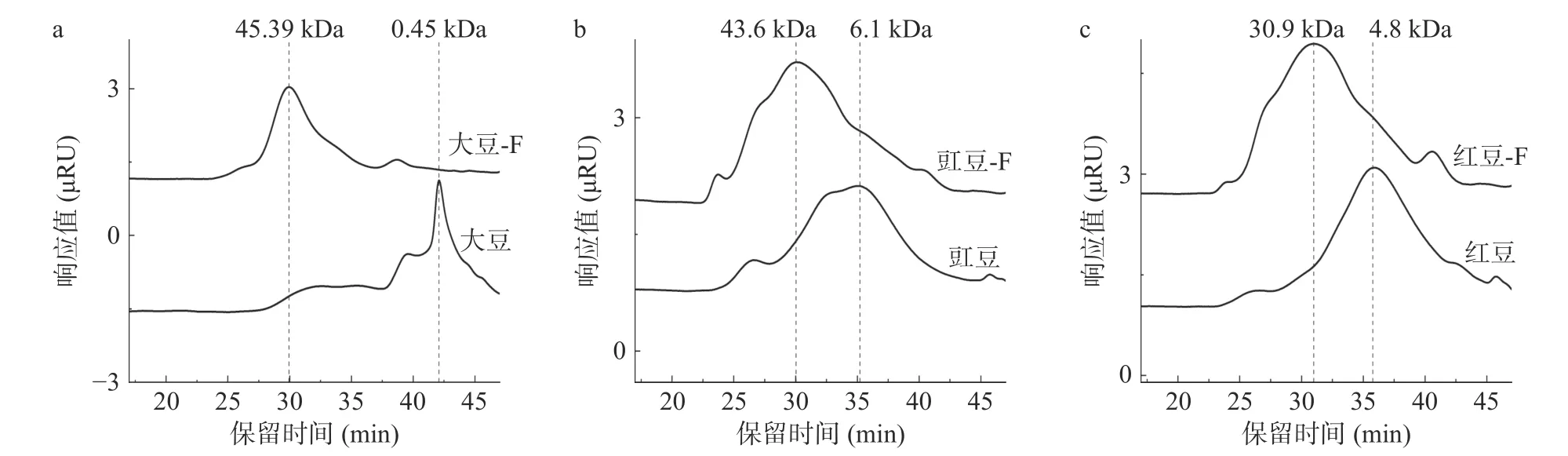
图1 8 种豆粉发酵前后粗多糖的分子量分布Fig.1 Molecular weight distribution of eight kinds of legume crude polysaccharides before and after fermentation
2.3 发酵前后豆类中粗多糖红外光谱比较分析
8 种豆粉发酵前后粗多糖的红外光谱分布如图2 所示。发酵前后豆粉粗多糖在3426 cm-1附近均具有一个强且宽的特征吸收峰,这是由于-OH 官能团的伸缩振动引起的,表明糖类分子形成分子内氢键[25]。发酵前和发酵后8 种豆粉粗多糖在2925 和2932 cm-1之间都有特征峰,并以小肩峰的形式存在,这是糖类C-H 伸缩振动的特征峰[1]。大部分羰基吸收集中1900~1650 cm-1,通常为谱图的最强峰或次强峰,为C=O 官能团的伸缩振动,未发酵豆和发酵豆粗多糖都含有此特征峰,发酵前大豆和豌豆只含有一个C=O 伸缩振动吸收峰,其余豆粉粗多糖含有两个C=O 伸缩振动吸收峰。表明粗多糖样品中可能存在不同种类的糖醛酸[26]。1640 cm-1是蛋白质的特征红外吸收峰[27],所有豆粉粗多糖样品在1640 cm-1附近都未出现强烈的吸收峰,表明豆粉样品用TCA 去除蛋白效果明显。在1500~600 cm-1区域主要提供C-H 弯曲振动的信息[28]。扁豆、扁豆-F、蚕豆、蚕豆-F、豇豆、豇豆-F、红豆、红豆-F、菜豆、绿豆、绿豆-F 和豌豆粗多糖在 763 和 931 cm-1附近出现吸收峰,表明粗多糖中可能存在吡喃糖环结构[29]。此外,840 cm-1附近的峰值表明粗多糖样品中包含α-构型[30],即具有α-糖苷键,这种豆子粗多糖可能是α-吡喃葡萄糖[31]。发酵前后豆子粗多糖中均含有α-糖苷键。红外光谱图结果分析表明,8 种豆类发酵前后提取物粗多糖均符合多糖分子的结构特征。
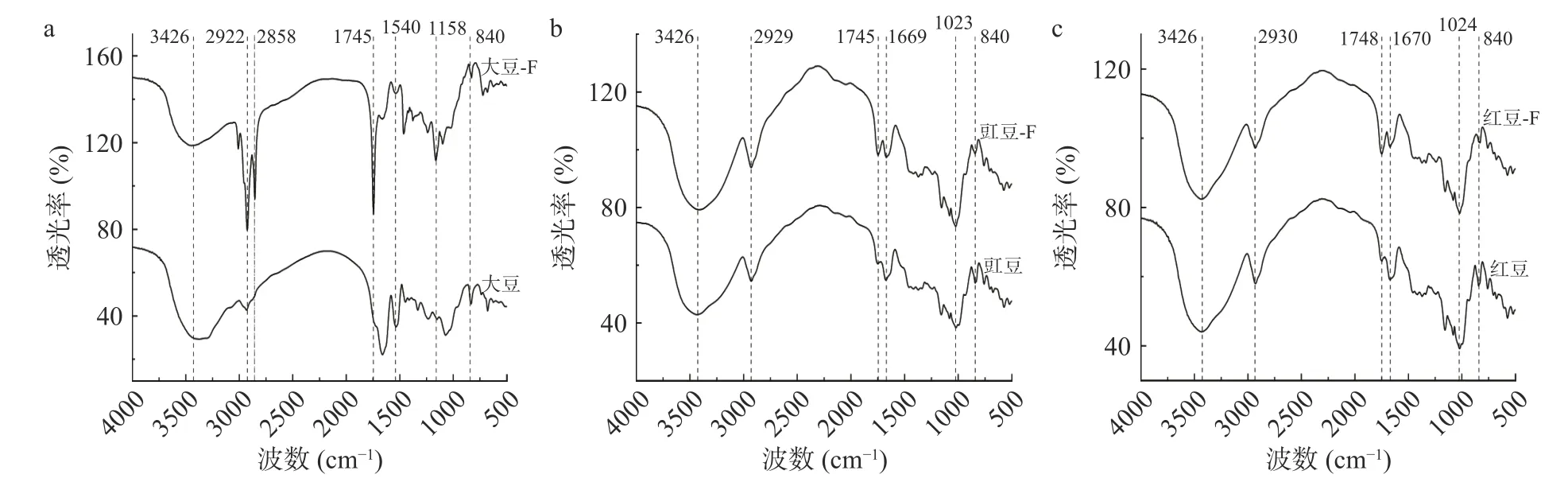
图2 8 种豆粉发酵前后粗多糖红外光谱分布Fig.2 IR spectral distribution of eight kinds of legume crude polysaccharides before and after fermentation
2.4 发酵前后豆类中粗多糖抗氧化活性比较分析
2.4.1 DPPH 自由基清除能力 8 种豆粉发酵前后粗多糖对DPPH 自由基清除能力比较分析结果如图3 所示。研究结果表明,增加8 种豆粉粗多糖的浓度,DPPH 自由基清除能力均增加,但清除能力均小于Vc。由粗多糖溶液对DPPH 自由基的清除曲线经回归处理可求出DPPH 自由基的半清除率(Half aximal inhibitory concentration,IC50),IC50值见表2。除蚕豆外,未发酵豆粉粗多糖对DPPH 自由基清除能力均显著优于发酵后豆粉粗多糖(P<0.05),这可能与未发酵豆粉中含蛋白质有关。SIU 等[32]从三种可食用蘑菇中提取了不同蛋白含量的多糖,结果表明多糖抗氧化活性和其蛋白质含量具有显著相关性。发酵后豆子粗多糖对DPPH 自由基清除能力降低可能是由于发酵使多糖-蛋白质复合物空间结构发生改变,失去了粗多糖结合的蛋白质,从而降低了抗氧化活性。IC50值显著性分析见表2,发酵前和发酵后的豆粉粗多糖清除DPPH 自由基能力差异显著(P<0.05)。发酵后的大豆、红豆、蚕豆、豌豆和豇豆对DPPH 清除率效果较好,当质量浓度为0.5 mg/mL时,清除率均超过50%。发酵后绿豆粗多糖的清除效果较低,当质量浓度为1.5 mg/mL 时,清除率未达到50%。
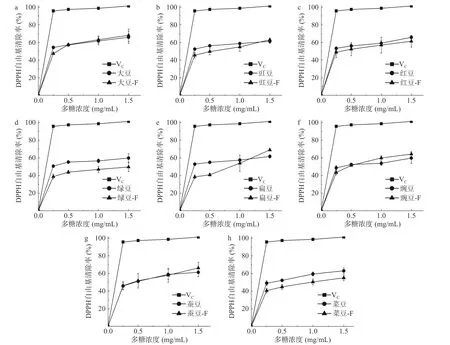
图3 发酵前后8 种豆粉粗多糖DPPH 自由基清除能力Fig.3 DPPH free radical scavenging capacity of eight kinds of legume crude polysaccharides before and after fermentation
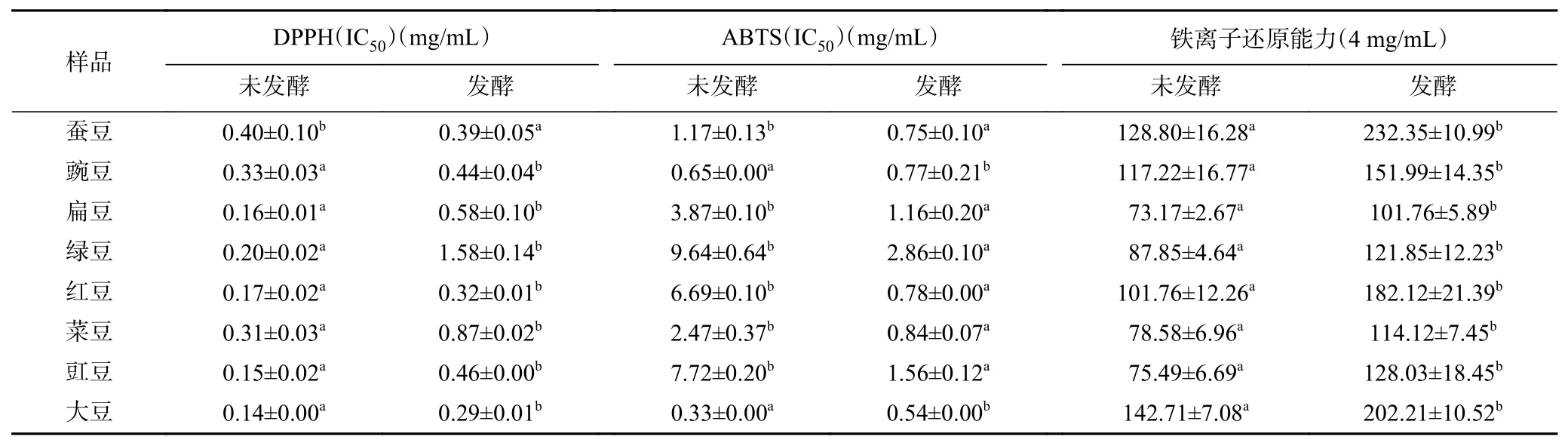
表2 8 种豆粉发酵前后粗多糖抗氧化活性显著性分析Table 2 Significance analysis of antioxidant activity of eight kinds of legume crude polysaccharides before and after fermentation
2.4.2 ABTS+自由基清除能力 8 种豆粉发酵前后粗多糖对ABTS+自由基清除能力的结果如图4 所示。研究结果表明,增加8 种豆粉粗多糖浓度,ABTS+自由基清除率能力均增加。发酵前后豆粉粗多糖对ABTS+自由基的半清除率的IC50值见表2。发酵前后豆粉粗多糖对ABTS+自由基清除率差异显著(P<0.05)。其中豇豆、红豆、绿豆、扁豆、蚕豆和菜豆发酵后豆粉粗多糖对ABTS+自由基清除率均比发酵前显著增加。文愉熙等研究报道,多糖的活性还与糖苷键、空间构象等有密切关系[33],在豆粉粗多糖红外检测中,发酵前大豆和豌豆在C=O 官能团伸缩振动上与其它豆粉粗多糖有差异,可能对ABTS+自由基清除能力有影响。多糖分子结构的作用机制也会影响到多糖的抗氧化活性[34]。当粗多糖质量浓度为1 mg/mL 时,发酵后大豆、蚕豆、红豆、豌豆和菜豆对ABTS+自由基清除率超过50%,清除效果较好。发酵后绿豆粗多糖清除效果不佳,当质量浓度为4 mg/mL 时,清除率达55.70%。
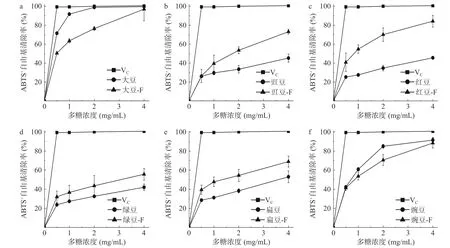
图4 发酵前后8 种豆粉粗多糖ABTS+自由基清除能力Fig.4 ABTS+ radical scavenging ability of eight kinds of legume crude polysaccharides before and after fermentation
2.4.3 铁离子还原能力(FRAP) FRAP 法可作为样品中的总抗氧化能力的指标[35]。8 种豆粉发酵前后粗多糖对铁离子还原能力结果如图5 所示。研究结果表明,随着粗多糖质量浓度的增加,发酵前后豆粉粗多糖对铁离子的还原能力均随浓度增加而增加。发酵过程能提高豆粉粗多糖对铁离子的还原能力。如表2 所示,当粗多糖样品浓度达4 mg/mL 时,发酵后豆粉粗多糖比未发酵豆粉粗多糖对铁离子还原能力均显著增加(P<0.05)。
图5 发酵前后8 种豆粉粗多糖铁离子还原能力Fig.5 Iron ion reducing ability of eight kinds of legume crude polysaccharides before and after fermentation
3 结论
本研究用枯草芽孢杆菌ATCC 6051 固态发酵了大豆、豇豆、红豆、绿豆、扁豆、豌豆、蚕豆和菜豆8 种不同的豆粉,比较分析了发酵前后8 种豆粉粗多糖的分子结构和抗氧化生物活性的变化。通过枯草芽孢杆菌发酵后制备得到的豆粉粗多糖分子量均增加、粗多糖分子中糖醛酸含量也不同程度的提高。发酵豆粉粗多糖与未发酵豆粉粗多糖表现出不同的抗氧化生物活性。除蚕豆外,发酵后的其它豆粉粗多糖对DPPH 自由基清除能力均降低;发酵后的大豆、红豆、蚕豆、豌豆和豇豆对DPPH 自由基清除率较好,当质量浓度为0.5 mg/mL 时,清除率均超过50%。发酵后的豇豆、红豆、绿豆、扁豆、蚕豆和菜豆粗多糖对ABTS+自由基清除率均增加。发酵后的豆粉粗多糖对铁离子还原能力均优于未发酵的豆粉粗多糖。综上,枯草芽孢杆菌发酵后可获得具有较强抗氧化活性的生物活性多糖,通过发酵前后豆类多糖的对比研究,为枯草芽孢杆菌发酵后豆类多糖的实际应用研究提供了科学理论基础,对进一步的应用研究及今后开发枯草芽孢杆菌发酵制备具有新型生物学功能的豆类多糖分子提供了新思路,后续还需对发酵前后豆类粗多糖的具体功能展开进一步研究。
猜你喜欢
食品工业(2021年12期)2021-12-31
湖南饲料(2021年4期)2021-10-13
中国食品(2021年5期)2021-03-24
食品研究与开发(2020年1期)2020-03-09
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年8期)2019-11-23
中国油脂(2019年6期)2019-08-22
扬子江(2019年3期)2019-05-24
当代化工研究(2016年7期)2016-03-20
