
先天性恐惧防御行为的神经回路研究进展
2023-11-04 08:42赵佳佳吴永叶杨丽萍
浙江大学学报(医学版) 2023年5期
赵佳佳,宋 琦,吴永叶,杨丽萍
河南中医药大学医学院,河南 郑州 450046
恐惧是危险刺激诱发的一种负性情绪,依据是否依赖后天学习将恐惧分为先天性恐惧和习得性恐惧[1-3]。先天性恐惧防御反应是与生俱来的反应,如恐高症和老鼠面对天敌时的恐惧情绪反应。先天性恐惧刺激主要包括天敌、具有攻击性的同类、疼痛或危险的环境特征。在面对威胁生命安全的恐惧刺激时,动物会做出先天性防御反应以维持生存。先天性恐惧防御反应的产生离不开感知系统的参与,高级中枢通过视觉、听觉、触觉以及嗅觉等多感官输入外界信息,用以趋利避害。先天性恐惧防御反应主要包括冻结、逃跑、攻击等[4]。先天性恐惧对于物种生存不可或缺[5],但过度恐惧也会导致精神障碍类疾病,如恐惧症、焦虑症、创伤后应激综合征等[6-8]。因此,对先天性恐惧防御反应的神经环路结构和功能解析不仅可以深入了解先天性防御行为产生的工作机制,而且可以为精神疾病的诊断和治疗提供神经生理学依据。本文从不同感官系统接收的信号诱导防御行为角度综述了动物先天性恐惧防御行为神经环路的研究进展,尤其是嗅觉、视觉、听觉三个系统的投射环路,以期为揭开特定神经环路参与精神障碍类疾病的机制研究提供参考。
1 嗅觉信号诱导的先天性恐惧防御行为神经环路
嗅觉系统是动物感知外部环境的化学成分信息,以定位和评估食物、交配、社会交往以及避免危险的特殊系统。对于啮齿类动物来说,嗅觉是最重要的感觉系统,其嗅觉敏感度是灵长类动物的四倍[4],因此基于嗅觉系统对先天性恐惧环路的研究主要集中于啮齿类动物。
通过嗅觉系统检测的危险信号有捕食者的分泌物(如粪便、尿液)、攻击性或受威胁的同物种释放的警觉信息素等,均会诱发动物产生强烈的恐惧情绪[1,9-10]。目前对嗅觉恐惧神经环路的研究多数使用狐狸尿液、TMT 和2MT[11-13]。TMT 和2MT均能模拟天敌气味,且可以稳定诱导啮齿类动物产生冻结行为,多用于先天性恐惧研究[14-16]。
动物的嗅觉系统包括主嗅系统、副嗅系统、Grueneberg 神经节、马赛拉中膈器官和三叉神经系统,而对先天性恐惧起作用的主要是主嗅系统和副嗅系统。研究发现,主嗅系统是由主嗅球识别到TMT 并进行初步处理,而后将信号传给杏仁核皮质部进而调控冻结反应[17]。而副嗅系统是通过犁鼻器-副嗅球-内侧杏仁核环路来调控TMT诱导的冻结行为[18]。利用光遗传选择性激活外侧缰核向外背侧被盖区投射的Glu能神经元足以产生冻结行为[19]。CART 神经肽是先天性恐惧反应的重要调节因子。Rale等[20]发现CART信号可以通过N-甲基-D-天冬氨酸受体介导的Glu 能输入调节中央杏仁核-腹侧终纹床核环路活性,从而参与TMT 诱导的冻结反应。中央杏仁核中表达5-羟色胺2A 受体的神经元集群优先控制TMT诱导的先天性防御的行为输出[21]。以上研究表明,TMT 暴露能稳定诱发啮齿类动物产生冻结反应。此外,光遗传激活前扣带皮层-基底外侧杏仁核环路可抑制狐狸尿诱导的冻结行为[22-23]。
其他防御反应还包括回避行为和应激激素水平的变化等。缰核-脚间核环路介导了狐狸尿引起的防御反应,抑制这条环路能显著减少回避行为[24]。还有研究发现,嗅皮层亚区和室旁核同时被天敌气味激活,嗅皮层亚区向室旁核的促肾上腺皮质激素释放激素神经元的投射可以调控应激激素水平[25]。
PAG 是恐惧表达网络的下游脑区,是协调先天和后天恐惧防御反应的关键。PAG 在不同的防御行为中激活程度不同,在逃跑时的激活程度更为显著[11]。且PAG 编码不同程度的捕食性威胁,而不是直接协调不同的威胁导向行为[26]。目前已经发现的嗅觉信号诱导的先天性恐惧不同神经环路交互模式见图1。
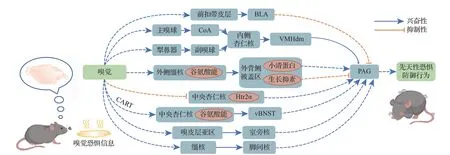
图1 嗅觉信号诱导的先天性恐惧神经环路示意图Figure 1 Neural circuit pattern of olfactory congenital fear
2 视觉信号诱导的先天性恐惧防御行为神经环路
相比其他感知系统,视觉获取的信息更为丰富、快速和高效,尤其人类视觉信息的获取占所有感知觉信息的70%以上[27]。在面临威胁时动物依赖视觉系统能快速分辨天敌的物种和距离,并结合周围环境信息做出最适宜的防御行为。
能激发先天性恐惧的视觉刺激有looming 刺激、天敌暴露和高度刺激等。Looming刺激是一种模拟天敌从高空逐渐逼近的视觉刺激(上视野不断变大的阴影)[28],能稳定诱发各种动物的先天性防御行为[29-33]。天敌暴露包括猫、大鼠、蛇和蜘蛛等。动物面临威胁时采取何种防御行为依赖周围环境和自身状态。在有遮蔽物存在的情况下,使用looming刺激时,若小鼠对周围环境熟悉会采取逃跑行为,若处在陌生环境中则会产生冻结行为[34]。由此可知,对先天性恐惧的防御反应并不是简单的刺激反应,而是结合先前经验和自身环境的灵活反应从而选择最恰当的防御行为[5,35-36]。
相比嗅觉系统,视觉系统诱发的先天性防御反应更多是逃跑。作为视网膜接收器的上丘与视觉诱导的先天性恐惧关系最为密切[37-38]。目前已发现多条上丘相关神经环路参与looming 刺激诱发的先天性防御反应。例如,Lin 等[39]发现上丘-ZIr-丘脑连接核的三突触环路介导looming 刺激诱导的逃跑反应。上丘内的小清蛋白阳性神经元向二叠体旁核投射的环路可以调节逃跑行为[40]。但又一项研究表明,上丘-二叠体旁核环路并不能介导逃跑行为,而是由二叠体旁核向上丘的反向投射发挥作用。有学者还发现危险信号在内侧上丘进行整合处理,传递到dPAG通过突触阈值启动逃跑行为[41]。上丘的Glu能神经元向腹侧被盖区的GABA 能神经元投射,腹侧被盖区再将信息传递给中央杏仁核的三级投射环路介导了looming刺激诱导的逃跑行为[42]。vLGN向内侧上丘的GABA 能投射环路同样可以调节逃跑反应[43-44]。此外,表达CCK 的下丘脑PMd 神经元对于逃跑是必要的,PMdCCK-dlPAG 环路的激活足以诱导先天性防御反应[45-46]。下丘脑VMHdm/c的类固醇生成因子神经元的不同投射分别调节不同的防御行为,如激活VMHdm/c-下丘脑前核环路促进逃避防御行为,而激活VMHdm/c-dPAG 环路则促进冻结行为[5,47-48]。一项研究发现背侧海马-背外侧隔膜-下丘脑神经环路的活化减少了逃跑和抵抗等防御行为[49]。最近一项研究表明,下丘脑的外侧臂旁核中表达CCK 的阳性神经元向室旁核的投射环路参与逃跑行为[50]。
研究发现,内侧中层上丘-丘脑外侧后核-外侧杏仁核环路介导了looming 刺激诱发的先天性恐惧防御行为,其可能是通过中央杏仁核-PAG输出环路发挥作用[51]。上丘内的PV 阳性神经元向丘脑外侧后核的投射环路同样介导looming 刺激引起的冻结行为[40]。Salay 等[34]通过化学遗传发现vLGN 向丘脑连接核和上丘的投射对防御反应有相反的影响,激活vLGNGABA-丘脑连接核环路活性显著延长looming 刺激诱导的冻结时间;而激活vLGN-上丘环路活性则缩短冻结时间[52]。
腹侧中线丘脑的两个亚区剑突核和丘脑连接核介导两种性质相反的防御行为,丘脑连接核-内侧前额叶环路促进攻击性反应如颤尾,而亚区剑突核-基底外侧杏仁核环路介导逃跑、冻结等防御反应[53]。通过功能性磁共振成像、电生理及光遗传等技术手段对灵长类动物进行研究发现,当暴露于蛇时,猴子和人类的VMH 均被显著激活,而暴露于恐怖面容时杏仁核的激活明显高于中性和快乐面容[54-55]。视觉信号诱导的先天性恐惧神经环路交互模式见图2。
3 听觉信号诱导的先天性恐惧防御行为神经环路
听觉系统在检测危险信号中同样发挥重要作用,尤其是对于夜间活动的动物。听觉系统接受危险的声音信息并将其传递到中枢听觉系统,中枢听觉系统对声音进行加工、分析,感知声音时程、音色、音强和空间信息,再整合其他感官系统信息判断危险的方位、来源等后做出相应的防御反应,这对维系个体与种群的生存具有极其重要的意义。
诱发动物产生先天性恐惧情绪的听觉刺激主要有噪音、同类发出的警报声、逐渐逼近的声音刺激等[56]。大鼠在遇到威胁生命的恐惧刺激时会发出22 kHz的超声,这个声音会引起同类产生先天性防御行为,而小鼠在1~64 kHz的宽频信号下会产生恐惧情绪[57]。
ACx在听觉系统中发挥重要作用。听觉刺激信息由ACx 传递给下丘皮质,进而驱动dPAG 产生逃跑行为,且该环路可以放大先天性防御反应[58]。ACx 向lPAG 的Glu 能投射神经元被噪音显著激活,光遗传兴奋ACxGlu-lPAG 神经环路引起逃跑行为[59]。此外,Li等[60]发现兴奋ACx-上丘环路活性同样可以产生逃跑行为。未定带在调节听觉诱导的先天性防御行为方面发挥了重要作用。ZIr向dPAG的GABA能投射调控高分贝噪声诱导的逃跑行为[61]。此外,Wang 等[62]发现初级体感皮层-腹侧未定带-丘脑后内侧核环路同样参与噪声引起的逃跑反应。
ACx 向纹状体尾部表达D2 受体的椎体神经元的投射介导了渐强声音诱导的冻结行为[60]。ZIrGABA-vPAG 的抑制性投射可以调控高分贝噪声诱导的冻结行为[61]。最近研究发现,RtTg 的Glu能神经元被白噪音特异性激活,且耳蜗核-RtTgGlu-脊髓运动神经元环路介导听觉惊吓反射[63]。低强度聚焦超声刺激通过激活PAG 介导多种防御行为,如位置特异性被动回避、逃跑和冻结等行为[64]。听觉信号诱导的先天性恐惧不同神经环路交互模式见图3。

图3 听觉信号诱导的先天性恐惧神经环路示意图Figure 3 Neural circuit pattern of auditory congenital fear
4 其他感知觉信号诱导的先天恐惧防御行为神经环路
除了嗅觉、视觉和听觉外,其他感知觉信号也可触发先天性恐惧防御反应,如触觉、前庭觉和窒息感等。不同感官系统信号诱导的先天性恐惧神经环路研究见表1。Choi等[65]在前外侧通路中发现两个对皮肤机械刺激(热刺激、有毒刺激)高度敏感的脊髓投射神经元集群,分别表达速激肽1 型受体和G 蛋白偶联受体83,这两个集群投射向外侧臂旁核的不同亚核分别调控逃跑行为和自主反应。广场恐惧症是一种空间感知改变综合征,诊断标准是前庭症状和对视觉空间运动刺激的敏感性,采用多种影像学方式对亚临床广场恐惧症患者的大脑结构和功能进行研究后发现,患者的前庭视觉-空间-额叶网络的功能结合性降低[66]。临床研究表明,相较于健康者,杏仁核受损患者暴露于危险刺激时恐惧显著减少[67-68]。但另一项研究发现,三位杏仁核损伤患者吸入35%二氧化碳后会发生恐惧和惊恐发作,其恐惧程度远高于健康者[69]。说明了杏仁核在恐惧中发挥了重要作用,但不是恐惧产生所必需的,如二氧化碳诱发的恐惧不会被杏仁核损伤所影响。

表1 不同感官系统信号诱导的先天性恐惧神经环路研究一览Table 1 Summary of innate fear neural circuits of different sensory systems
5 结 语
近些年来随着遗传学的发展出现了各种记录和操纵神经元群体的工具,如光遗传、在体钙成像和神经环路示踪技术等,先天性恐惧防御反应的神经机制研究因此取得更多进展。相比对脑功能探索的传统技术,这些工具证实了对细胞类型特异性的大脑区域研究的重要性,同时提示了调控不同先天性防御行为神经环路机制的多样性和复杂性。相同威胁刺激诱发的同一恐惧防御行为常有多条神经环路参与,但这些神经环路之间是如何相互作用尚不清楚。而对于相同威胁刺激诱发不同恐惧防御反应,除了由不同神经环路介导之外,脑内活性物质如神经递质是否参与的研究尚少。目前对先天性恐惧神经机制的研究集中在啮齿类动物上,对灵长类动物的研究较少,因此研究结果难以实现临床转化。
未来对先天性恐惧神经环路的研究还应注意以下方面:①参与同一威胁刺激诱发相同防御行为的神经环路如何相互作用;②结合神经递质荧光分子探针等技术实现特异性动态检测先天性恐惧相关脑区活性物质的含量变化;③随着影像学的不断发展,可以利用功能性磁共振成像、脑电图和脑磁图等先进技术对灵长类动物的先天性恐惧防御行为的神经环路进行更详细的研究;④通过跨物种比较,揭示不同物种之间先天性恐惧防御行为神经环路的共同特点和差异,探究这些神经环路的保守性和多样性。
志谢 研究得到国家自然科学基金(81973596)、河南省科技攻关项目(212102310341)支持
Acknowledgements This study was supported by the National Natural Science Foundation of China (81973596),Key Science and Technology Project of Henan Province (2121 02310341)
利益冲突 所有作者均声明不存在利益冲突
Conflict of Interests The authors declare that there is no conflict of interests
©The author(s) 2023.This is an open access article under the CC BY-NC-ND 4.0 License (https://creativecommons.org/licenses/by-nc-nd/4.0/)
猜你喜欢
浙江大学学报(医学版)(2023年2期)2023-08-08
磁共振成像(2022年6期)2022-07-30
天津医科大学学报(2021年4期)2021-08-21
中国医学影像技术(2021年2期)2021-03-30
心肺血管病杂志(2020年5期)2021-01-14
音乐生活(2020年12期)2020-01-05
上海公路(2018年4期)2018-03-21
中国当代医药(2015年30期)2015-03-01
电测与仪表(2014年8期)2014-04-04
现代防御技术(2014年6期)2014-02-28
