
施氮和间作对春小麦土壤丛枝菌根真菌群落结构的影响
2023-11-21 13:53王爱萍姜彧宸冯月池田董琦
山西农业大学学报(自然科学版) 2023年5期
王爱萍,姜彧宸,冯月,池田,董琦*
(1.山西农业大学 农学院,山西 晋中 030801;2.黄土高原特色作物优质高效生产省部共建协同创新中心,山西 晋中 030801)
丛枝菌根真菌(Arbuscular mycorrhizal fungi,AMF)是一种在土壤中广泛分布的真菌,属于球囊菌门(Glomeromycota),可以侵染80%的陆生植物根系并与植物形成互利共生关系[1],扩大根系的吸收范围和吸收表面积,给寄主植物提供氮、磷等矿质营养,并提高植物对逆境的抗性,同时植物为AMF 生长提供所需要的碳源[2]。AMF 对维持自然生态系统的功能和稳定具有重要意义。全球AMF物种丰富度范围在341~1600 个OTU,甚至可能多于1600 个OTU[3],Öpik 等[4]报道鉴定形态的AMF约250 种,鉴定分子的为350~1000 种,Tedersoo等[5]报道共发现11 科25 属250 种,Davison 等[6]报道AMF 有348种。
农业施肥对AMF 群落组成有显著影响,施氮影响AMF 群落组成,降低群落多样性[7]。增施氮肥降低AMF 生物量、物种丰富度和多样性[8-10]。还有研究表明,在磷含量有限的土壤中,施氮可增加AMF 物种的丰富度和多样性;富磷土壤中,AMF物种减少[11-12]。也有研究表明,长期施氮对微生物生物量和群落组成没有影响[13],关于氮肥影响植物AMF 群落组成前人有报道,但肥料驱动AMF 群落组成变化的功能也知之甚少[14]。
土地利用方式对AMF 群落组成的影响很大[15],间作与AMF 之间存在互利关系,由于间作间的相互作用,AMF 的多样性和群落组成发生了变化[16]。Zhang 等[17]认为间作增加了大豆根际土壤AMF 多样性,而相邻玉米则没有这种影响。大豆和玉米根际AMF 优势属(Glomus)相对丰度随施氮量的增加呈相反的变化趋势。然而,也有研究表明间作不能改变AMF 的群落组成[18-20]。综上所述,不同的研究者关于间作对AMF 群落组成影响的研究结果不一致。
小麦和豌豆间作是我国传统间作方式之一,本研究通过施用不同的氮肥水平,对小麦根系土壤AMF 群落结构和功能进行分析,探究不同施氮量和间作种植模式对AMF 生物群落结构的影响,旨在为小麦的可持续生产以及农田土壤有益微生物的发掘和利用提供理论依据。
1 材料和方法
1.1 试验区概况和供试品种
试验于2022 年3 月至7 月在山西农业大学太谷校区农作站进行,地理坐标东经112°57′,北纬37°42′。太谷区2021 全年降雨量611.4 mm,2022年1-7 月降雨量199.7 mm。播前土壤含水量13.43%,试验田土壤为壤土,全氮含量为0.945 g·kg-1,碱 解 氮33.88 mg·kg-1,有 效 磷10.45 mg·kg-1,速 效 钾157.60 mg·kg-1,有 机 质21.96 g·kg-1。供试春小麦品种为宁春5 号,豌豆品种为中豌8号。
1.2 试验设计
采用随机区组设计,设置4 个施氮量:0 kg·hm-²(N0)、90(N1)、180(N2)和270 kg·hm-²(N3),2 种种植模式:春小麦单作(SS)、春小麦和豌豆间作(SI)。施氮量以纯氮计算,分别作为基肥在播种前一次性全部施入。春小麦和豌豆均于2022 年3 月1 日播种,且播量相同,均为225 kg·hm-²。间作小区为2行小麦间隔2 行豌豆,所有处理均为等行距种植,行距20 cm。小区面积5 m×5.2 m。每个处理设3 次重复。全生育期无追肥,只灌1 次水(3 月30 日),其余同当地管理。
1.3 样品的采集
春小麦成熟期,每个小区随机选取5 株小麦植株,采集0~10 cm 深度的小麦根系,去掉大块土壤后,轻轻抖落小麦根系附着的土壤,将混合后的土壤样品标记后立即放入液氮中保存备用。
1.4 AMF 群落结构的测定
采用SDS 法提取土壤样品的基因组DNA,检测质量后以稀释的基因组DNA 为模板,使用特异引 物(F:5′-AAGCTCGTAGTTGAATTTCG-3′和 R:5′-CCCAACTATCCCTATTAATCAT-3′)进行PCR 扩增。琼脂糖凝胶电泳检测PCR 产物,对检测合格的PCR 产物回收后构建文库。通过Illumina HiSeq 高通量测序平台进行测序。对得到的原始序列根据QIIME 过滤后获得高质量的序列,并与Unite 数据库进行比较,使用UCHIME 算法检测嵌合体序列并去除,最终得到有效序列。基于有效序列进行OTUs(Operational Taxonomic Units)聚类和物种分类。根据OTUs 聚类结果对每个OTU 的代表序列做物种注释以及进行丰度和Venn 图分析。选用FunGuild 进行功能注释,通过PCA 降维分析不同样本间的差异。
1.5 数据统计与分析
应用Excel 2016 记录数据,用T-test 进行差异显著性检验,使用Origin 2020绘图。
2 结果与分析
2.1 不同施氮量和间作处理春小麦根系土壤AMF 群 落OTU 组 成
物种积累箱形图反映了连续采样下新物种的出现率。在一定范围内,随着样本量的增加,如果箱形图的位置出现急剧上升,说明群落中发现了大量的物种;当箱形图的位置趋于平坦时,说明该环境下的物种不会随着样本量的增加而显著增加。从图1 可以看出,小麦根系土壤AMF 群落OTU 组成的所有样品中新种的出现趋于平缓,说明抽样充分,数据可以用于分析。
韦恩图可以直观看出不同处理春小麦根系土壤AMF 群落OTU 组成之间的差异及共有物种的情况。图2 显示,单作春小麦根系土壤AMF 群落OTU 个数为13,而间作为10 个。单作模式中,N0特 异 性AMF 群 落OTU 为2 个,占 总OTU 数 的15.38%,N1 和N3 中特异性OTU 均为3 个,占比均为23.08%,N2 中特异性OTU 最少,为1 个,占比7.69%。N0、N1、N2 和N3 共 有 的OTU 数 量 为2个,占比15.38%。间作模式中,N0 特异性AMF 群落OTU 为1 个,占总OTU 序列数的10.00%,N2和N3 处理特异性OTU 均为2 个,占比均为20.00%,N1 没有特异性OTU。N0、N1、N2 和N3共有的OTU 数量为3,占比30.00%。
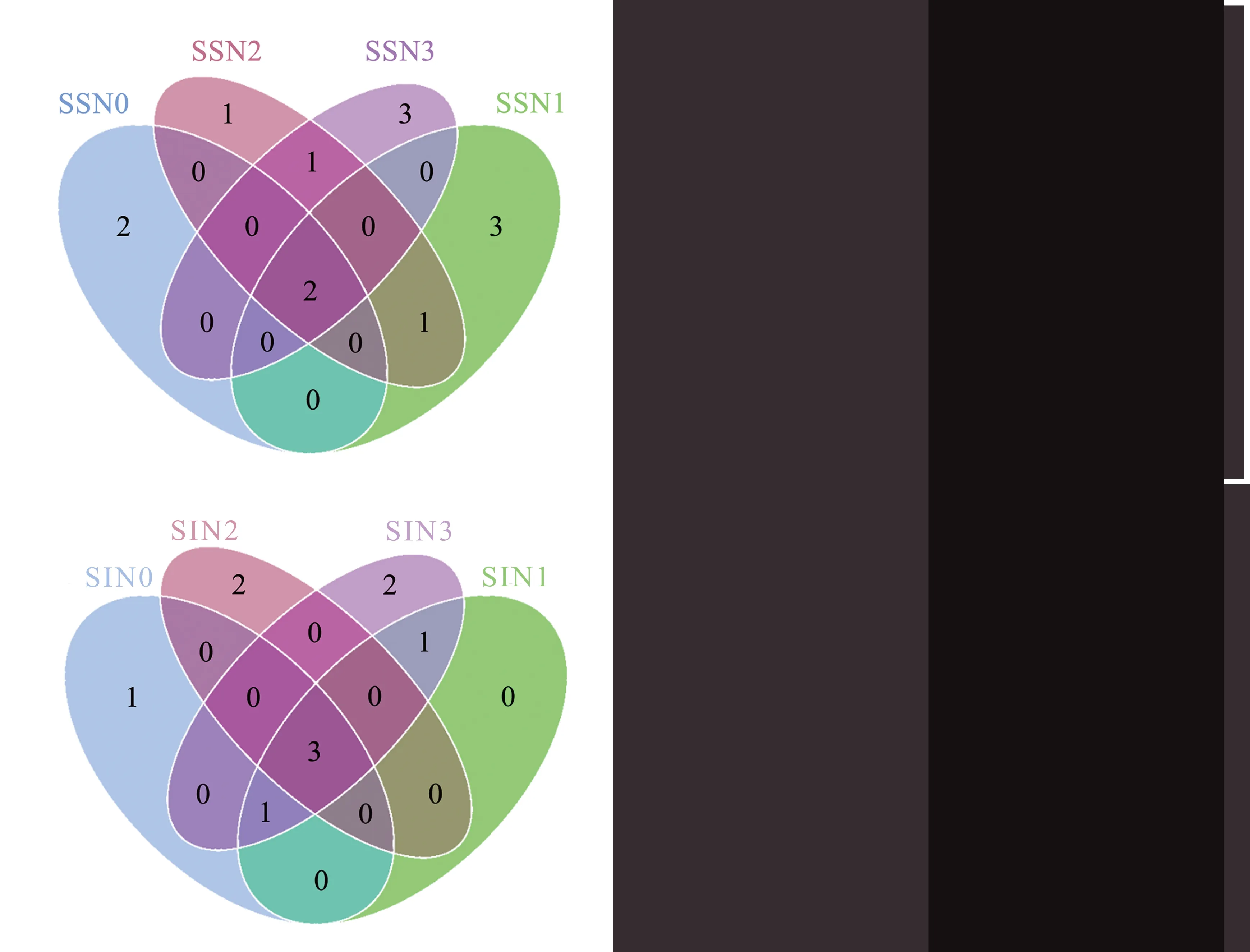
图2 不同施氮量和间作处理春小麦根系土壤AMF 韦恩图Fig.2 Venn graph of AMF community of spring wheat root soil under different nitrogen application rates and intercropping treatments
2.2 不同施氮量和间作处理春小麦根系土壤AMF 群落结构
2.2.1 科水平相对丰度的变化
图3 为AMF 群落科水平的相对丰度,可以看出,AMF 科水平主要有3 种,相对丰度排名从高到低依次为球囊霉科(Glomeromyceae,95.14%)、原囊霉目未知科(unidentified Archaeosporales sp.,3.47%)和类球囊霉科(Paraglomeraceae,1.39%),分别属于球囊菌纲(Glomeromycetes)球囊霉目(Glomerales)、原囊菌纲(Archaeosporomycetes)原囊 霉 目(Archaeosporales)和 类 球 囊 菌 纲(Paraglomeromycetes)类球囊霉目(Paraglomerales)。单作中,随着施氮量增加,球囊霉科的相对丰度增加,至N3 最高,占据100%丰度。施氮量为N0 时,与其他施氮水平相比,原囊霉目未知科的相对丰度最高;施氮量为N1、N2 时,类球囊霉科的相对丰度最高;施氮量为N3 时,球囊霉科的相对丰度最高。在间作处理中,施氮量增加至N1 处理后,只有球囊霉科1 种。施氮量为N0 时,与其他施氮水平相比,原囊霉目未知科的相对丰度最高;施氮量为N1、N2、N3 时,球囊霉科的相对丰度最高。同样的施氮水平,N0 处理下,间作明显提高了原囊霉目未知科的相对丰度,降低了球囊霉科的相对丰度;当施氮量为N1、N2 时,间作处理降低了类球囊霉科的相对丰度,提高了球囊霉科的相对丰度。说明种植方式和氮肥对于AMF 科水平的群落结构均有影响,施氮和间作二者的相互作用明显降低了科类水平的AMF 群落结构的丰富度,施氮导致AMF 群落丰度低的科水平群落有更替,高氮(N3)处理明显降低了科类水平AMF 的群落多样性。
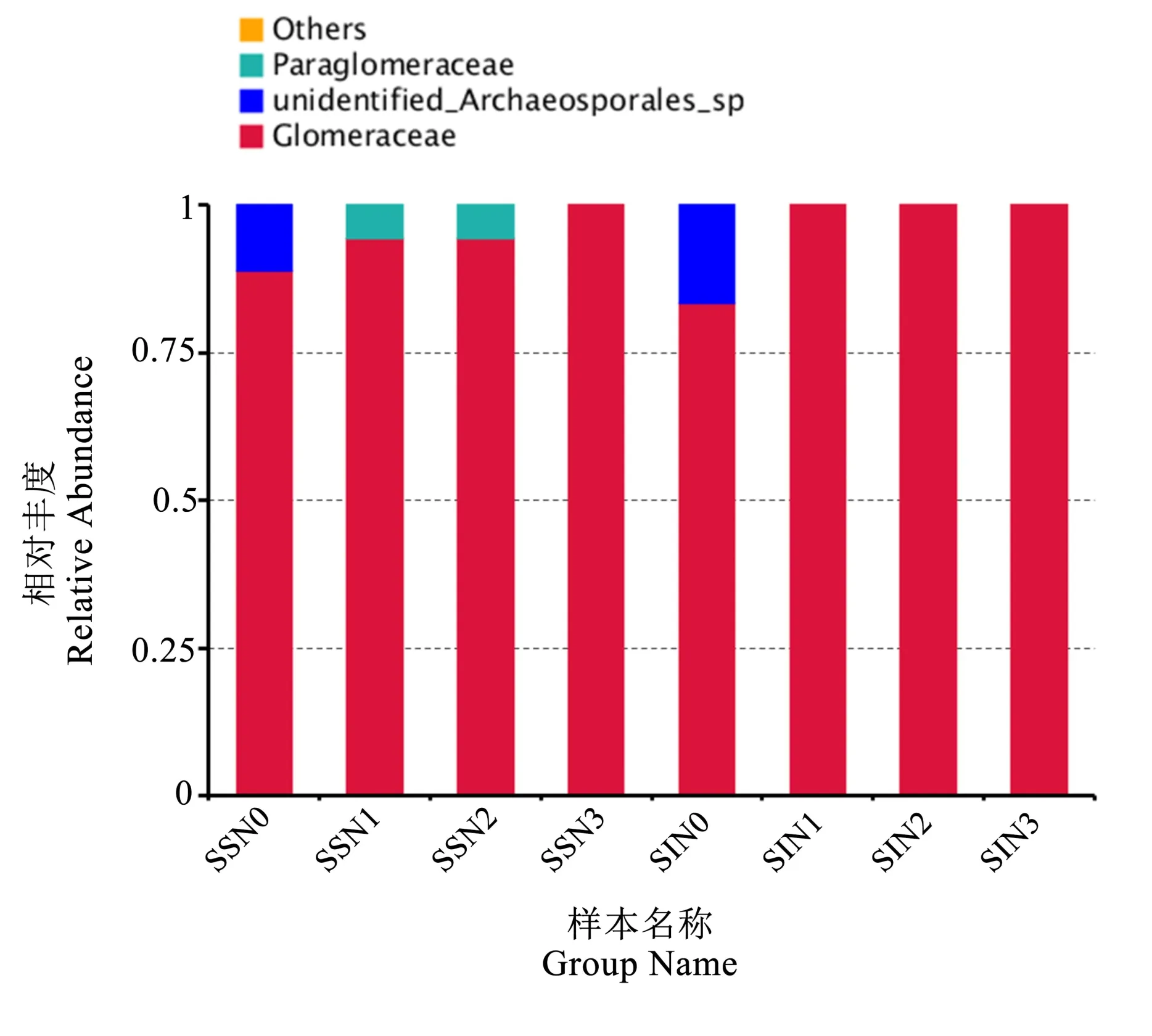
图3 春小麦根系土壤AMF 群落科水平的相对丰度Fig.3 Relative abundance of AMF community at family level in spring wheat root soils
2.2.2 属水平的相对丰度变化
图4 显示AMF 群落属水平主要有5 种不同的属,相对丰度排名由高到低依次为管柄囊霉属(Funneliformis,51.39%)、未 知 属(undefined,38.89%)、根孢囊霉属(Rhizophagus,4.86%)、其他未知原囊霉目类真菌(undefinedArchaeospora-les,3.47%)和类球囊霉属(Paraglomus,1.38%)。管柄囊霉属(Funneliformis)和根孢囊霉属(Rhizophagus)均属于球囊霉科(Glomeromyceae)。单作模式中,随着施氮量的增加,未知球囊霉属的相对丰度增加,在N3 达到43.2%;管柄囊霉属相对丰度随着施氮量增加先增加后减少,在N1 时达到最高;根孢囊霉属相对丰度随着施氮量的增加呈现先降低再增加趋势,在N1 时相对丰度为0;其他未知原囊霉目类真菌仅在N0 时出现;类球囊霉属在单作N1、N2 时存在。间作所有施肥处理中,未知球囊霉属相对丰度随着施氮量的增加表现为先增加后降低再增加趋势,在N1 处最高;管柄囊霉属在N2时相对丰度最大;根孢囊霉属仅存在于N3 处理下;其他未知原囊霉目类真菌同单作一样,仅在N0 出现。相同的施氮水平下,N0、N1、N2 水平单作的属丰度多样性明显高于间作,N3 水平二者数目持平。说明施氮量和种植模式影响植物根系土壤AMF 群落属水平的多样性,低氮(N1)与中氮(N2)水平下,间作明显降低了AMF 群落属水平的群落多样性。
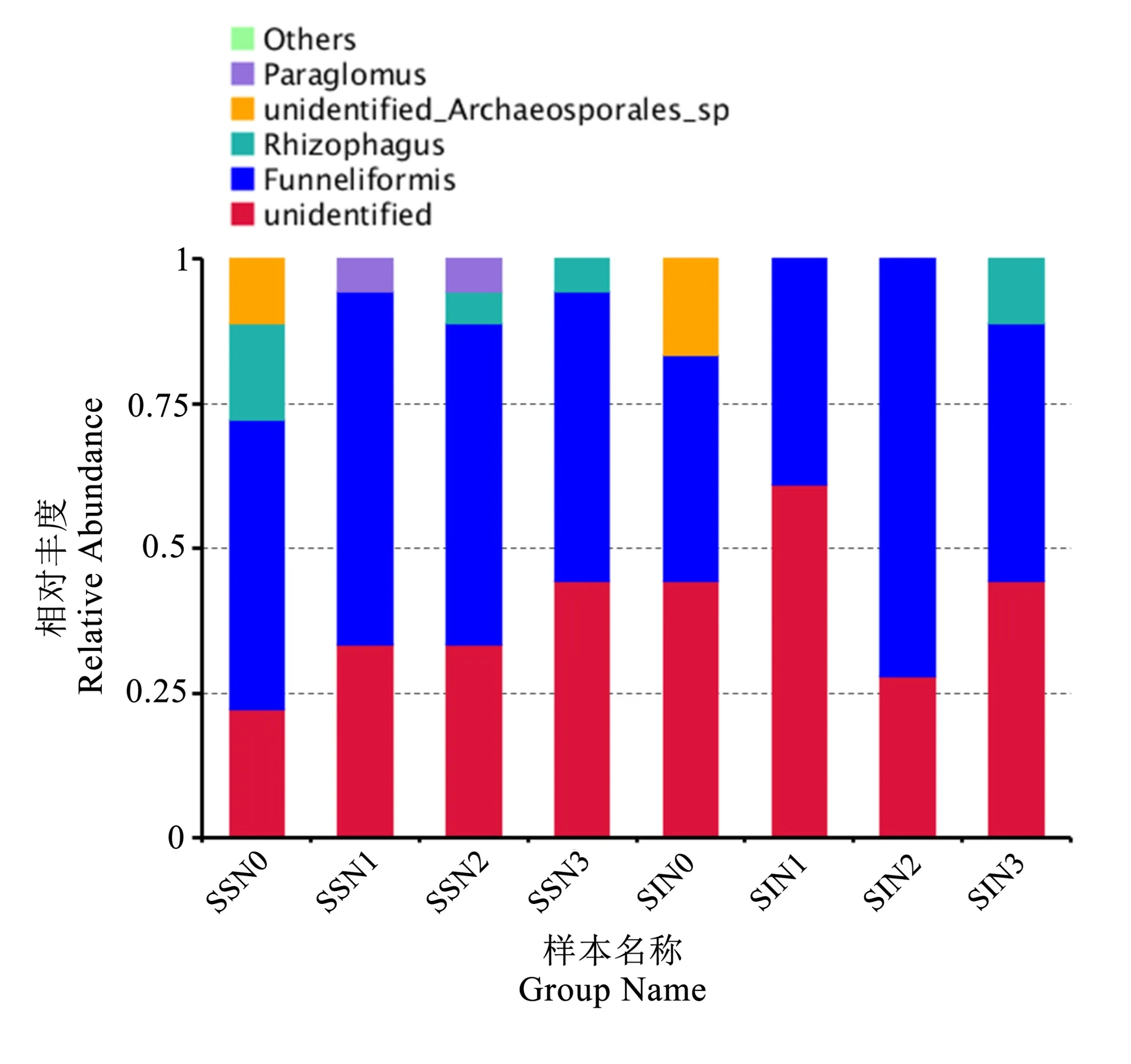
图4 春小麦根系土壤AMF 群落属水平的相对丰度Fig.4 Relative abundance of AMF community at genus level in spring wheat root soils
2.2.3 种水平的相对丰度及种类变化
种水平AMF 共鉴定出5种,相对丰度由高到低依次为地斗管囊霉(Funneliformis geosporum,51.39%)、球 囊 霉 科 未 知 种(Glomeraceaesp.,38.89%)、根 孢 囊 属 未 知 种(Rhizophagussp.,4.86%)、原囊霉目未知种(Archaeosporalessp.,3.47%)和隐类球囊霉(Paraglomus laccatum,1.39%)。AMF 种在各处理中出现的频度如表1 所示,地斗管囊霉和球囊科未知种出现的频度最高,且所有处理均有出现。根孢囊属未知种、原囊霉目未知种和隐类球囊霉在各处理的出现频度不同,单作4 个施肥处理根孢囊属未知种的出现频率高于间作;单作与间作N0 处理均有原囊霉目未知种出现,但添加氮肥处理后均消失;单作N1 和N2 处理均有隐类球囊霉出现,间作4 个施肥处理则未出现。说明施氮处理降低了原囊霉目未知种的相对丰度,间作降低了隐类球囊霉的相对丰度。表明,施氮处理和间作均会影响AMF 的种群分布,从而改变AMF的群落结构和特征,但对主要优势种地斗管囊霉和球囊霉科未知种的相对丰度影响不大。

表1 不同施氮量和间作下春小麦根系土壤AMF 种类分布Table 1 Distribution of arbuscular mycorrhizal fungi in root soil of spring wheat under different nitrogen application rates and intercropping
2.3 不同施氮量和间作处理春小麦根系AMF 群落功能的变化
根据样品在数据库中的功能注释及丰度信息绘制热图,并从功能差异层面进行聚类,不同的颜色梯度反映不同处理中物种功能预测的含量,越接近红色,含量越高。由图5 可以看出,单作中,N0 在动物病原体-内生菌-附生植物-植物病原-未定义的腐生物(Animal_Pathogen-Endophyte-Epiphyte - Plant_Pathogen - undefined_Saprotroph)菌群上存在优势,N1 在动物病原体-植物病原-土壤腐生菌-未定义的腐生物(Animal_Pathogen - Plant_Pathogen - Soil_Saprotroph - undefined_Saprotroph)菌群上存在优势,N2 在未定义的腐生物(undefined_Saprotroph)、真菌寄生虫-未定义的腐生物(Fungal_Parasite-undefined_Saprotroph)、动物内共生体-未定义的腐生物(Animal_Endosymbiont-undefined_Saprotroph)、植物病原(Plant_Pathogenh)菌群上存在优势,N3 在动物病原体-锁骨内生菌-真菌寄生虫(Animal_Pathogen-Clavicipitaceous_Endophyte-Fungal_Parasite)、腐生木(Wood_Saprotroph)、动物病原体(Animal_Pathogen)、植物病原-土壤腐生菌- 腐 生 木(Plant_Pathogen - Soil_Saprotroph -Wood_Saprotroph)、腐生类(Saprotrophic)等多种功能菌群上均存在优势。间作模式中,N0 在苔藓化(Lichenized)、粪腐生物-土壤腐生菌(Dung_Saprotroph-Soil_Saprotroph)菌群上均存在优势,N1在内生菌-植物病原(Endophyte-Plant_Pathogen)、粪腐生物-未定义的腐生物-腐生木及其他类型(Dung_Saprotroph-undefined_Saprotroph-Wood_Saprotroph)、其它(Others)、动物病原体-定义的腐生物(Animal_Pathogen-undefined_Saprotroph)功能菌群上均存在优势,N2 在植物病原-腐生木(Plant_Pathogenh-Wood_Saprotroph)菌群上存在优势,N3在地衣寄生虫-苔藓化(Lichen_Parasite- Lichenized)菌 群 上 存 在 优 势。SSN1 和SSN0、SIN0 和SIN2、SIN3 和SSN2 分别聚为一类。说明不同施氮量和种植方式对AMF 功能菌群丰度有明显的影响。
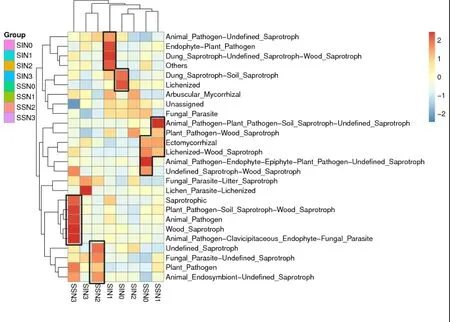
图5 FunGuild 功能注释聚类热图Fig.5 FunGuild annotation clustering heat map
基于功能注释的丰度统计结果进行PCA 降维分析,距离越接近,表示功能组成越相似。图6 中每个彩色点表示1 个样本,同一个处理的3 个重复使用同一种颜色和形状表示。将同一处理3 个重复的点连接起来形成1 个三角形,三角形的面积越小,组内重复性越好。红色的点表示每个处理对应的三角形的重心,点与点之间的距离越大,2 成分的组间的差异越大。由图6 可以看出,主成分1(PC1)与主成分2(PC2)解释方差分别为18.1%和15.9%,2个主成分累积贡献率达到34.0%。除SSN3 和SIN1 外,其余处理的距离较聚集,功能组成相似。在单作中,除N3 外,其余施氮量组内重复性强。间作中,随着施氮量的增加,重复性先变弱再变强,除N1 外,其余处理的功能性成分组内均较为聚集。同样的施氮水平下,N1 处理单作的重复性强于间作,N3 则间作强于单作。说明施氮量和间作对AMF功能性方面存在一定程度上的影响,间作与低氮处理二者的相互作用、单作与高氮处理二者的相互作用均会一定程度改变群落功能组成。
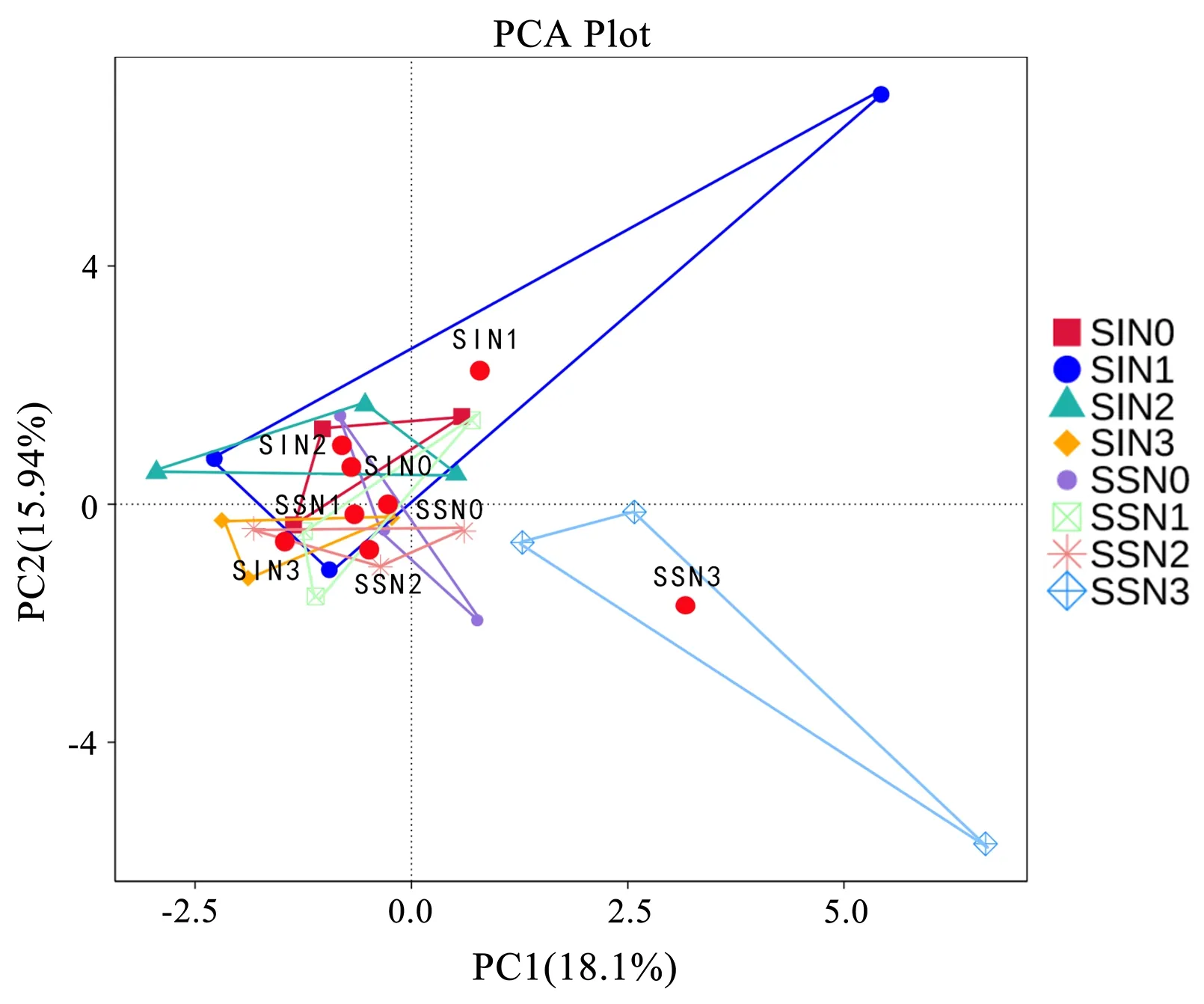
图6 FunGuild 功能注释PCA 结果展示Fig.6 PCA result display of FunGuild function annotation
3 讨论
由于许多AMF 在培养基中不可培养,因此鉴定环境中的AMF 分类群主要依赖于基于DNA 的方法[6]。本研究采用Illumina HiSeq 测序平台对麦田土壤AMF 测序分析,结果表明属于球囊菌门的AMF 科分类水平包括3 种,分别为球囊霉科(Glomeromyceae)、原囊霉目未知科(unidentified Archaeosporales sp.)和类球囊霉科(Paraglomeraceae),分别属于球囊菌纲(Glomeromycetes)、原囊菌纲(Archaeosporomycetes)和类球囊霉纲(Paraglomeromycetes),这与储薇等报道AMF 分为3 纲的报道一致[21]。杨文莹等报道,我国陆地农业生态系统土壤中AMF 种类共有20 属、125 种,其中以球囊霉属(Glomus)和无梗囊霉属(Acaulospora)最多[22]。而本研究中,相对丰度在1%以上的AMF 优势属为管柄囊霉属(Funneliformis)、undefined、根孢囊霉属(Rhizophagus)、其他未知原囊霉目类真菌(undefined-Archaeosporales)和类球囊霉属(Paraglomus)。虽 然AMF 的 宿 主 特 异 性 较 低[23],但AMF 对宿主植物具有选择性[24],不同地域与不同植物的AMF 种群有差异,宿主植物的种类影响AMF分布[25],这与宿主植物与AMF 间的亲和力有关,二者的相互作用决定亲和力和共生体的建立[26],小麦根系通过自身活动改变土壤环境,进而选择相应的AMF 种群共生。
间作对AMF 有增殖作用,但宿主和邻近植物对AMF 产生的影响不同[16],AMF 对间作物种也可能表现出不同的反应,这取决于邻近效应的强弱和土壤环境的相对变化[27-28]。本研究结果表明,间作影响小麦根系土壤AMF 群落属水平的多样性和种群分布,这可能与间作豌豆和麦田土壤特性有关,豌豆通过增加土壤有机质和氮的有效性增加AMF多样性[29],此外,麦田土壤pH、矿质营养等对AMF的生长、发育、侵染和繁殖均具有重要影响[30]。施氮影响AMF 群落组成,降低AMF 物种群落多样性[7-10],本研究结果也表明施氮明显促进球囊霉科(Glomeromyceae)相对丰度增加。N0 处理下,间作明显提高了未确定的原囊霉目未知科(unidentified Archaeosporales sp.)的相对丰度,降低了球囊霉科的相对丰度;当施氮量为N1、N2 时,间作处理降低了类球囊霉科的相对丰度,提高了球囊霉科的相对丰度。施氮导致丰度低的AMF 群落更替,高氮(N3)处理明显降低了科类水平AMF 的群落多样性,与前人研究结果一致。说明种植方式和氮肥以及二者的互作对于AMF 科分类水平的群落结构均有影响,施氮和间作二者的相互作用明显降低了科类水平的AMF 群落结构的丰富度。氮肥对AMF群落的影响可能与土壤磷素有效性有关,在磷含量有限的土壤中,施氮可增加AMF 物种的丰富度和多样性,但会导致富磷土壤中的AMF 物种减少[7],当土壤有效氮较高时,AMF 利用土壤氮作为自身的营养,对碳需求增加,AMF 朝着减少互利共生的方向发展[7]。
种群的多样性代表功能的多样性,AMF 种群高度多样化导致AMF 群落组成和多样性对施加氮的敏感性有差异[31],尤其对低水平氮肥的响应[7]。施用氮肥直接影响AMF 群落内的功能变化,可能减弱植物与AMF 的共生,AMF 功能的变化可归因于氮肥带来的AMF 群落组成的变化[7]。FunGuild功能注释聚类热图结果显示,SSN1 和SSN0、SIN0和SIN2、SIN3 和SSN2 分别聚为一类,表明AMF功能对土壤氮化学计量和间作的响应不同。因此,需要进一步研究氮和间作在介导植物-AMF 动力学中的作用,对于全面了解AMF 在农业中的作用具有重要意义。
4 结论
大 田 春 小 麦 根 系 土 壤AMF 分 为3 纲、3 目、3科、5 属、5 种。其中科分类水平包括3 种,分别为球囊霉科(Glomeromyceae)、原囊霉目未知科(unidentified Archaeosporales sp.)和 类 球 囊 霉 科(Paraglomeraceae);属水平为5 种,分别为管柄囊霉属(Funneliformis)、undefined、根孢囊霉属(Rhizophagus)、其他未知原囊霉目类真菌(undefined Ar-chaeosporales)和类球囊霉属(Paraglomus);种类水平分为5 种,分别为地斗管囊霉(Funneliformis geosporum)、球囊霉科未知种(Glomeraceaesp.)、根孢囊属未知种(Rhizophagussp.)、原囊霉目未知种(Archaeosporalessp.)和隐类球囊霉(Paraglomus laccatum)。施氮有利于球囊霉科(Glomeromyceae)相对丰度的增加,而低氮和中氮条件有利于单作模式类球囊霉科(Paraglomeraceae)相对丰度的增加。未施氮肥条件下,间作有利于原囊霉目未知科(unidentified Archaeosporales sp.)相对丰度的增加。施氮与间作二者的互作有利于球囊霉科(Glomeromyceae)相对丰度的增加。施氮量和间作以及二者的相互作用对AMF 科属种分类水平的菌群结构以及功能性方面均存在明显的影响。
猜你喜欢
农业科技通讯(2023年1期)2023-02-12
经济林研究(2022年4期)2023-01-12
广西林业科学(2022年2期)2022-05-09
河北果树(2020年4期)2020-11-26
山东农业科学(2019年11期)2019-12-24
江苏农业科学(2019年11期)2019-07-22
干旱地区农业研究(2017年5期)2017-12-18
农业环境科学学报(2017年2期)2017-03-20
新疆农垦科技(2016年2期)2016-08-21
农家顾问(2014年9期)2014-10-20
