
小麦新品种周麦32号的遗传构成解析
2024-01-09 13:54杨瑞晗孔子明高思亮
扬州大学学报(农业与生命科学版) 2023年4期
杨瑞晗, 孔子明, 高思亮
(1. 驻马店市农业科学院, 河南驻马店 463000; 2. 遂平县农业农村局, 河南遂平 463100)
小麦是我国重要的口粮作物,对保障粮食安全具有直接影响。优质是小麦遗传改良的重要目标之一,发展优质小麦亦已成为小麦供给侧改革的重要内容,而培育优质小麦品种则是支撑发展优质小麦产业的关键[1]。近年来,随着一批优质强筋小麦品种(如新麦26、郑麦366、郑麦7698、西农979、冀师02-1、济麦44、周麦32、西农511等)的选育与大面积推广,市场对优质强筋小麦的需求在一定程度上得到了缓解。在小麦生产实践中,优质强筋小麦品种与高产类型的品种在产量、抗病性、抗逆性等方面仍存在一定的差距。因此,从遗传角度上对具有代表性的优质强筋小麦新品种的遗传构成进行分析,探究其内在的遗传关系,可为优质强筋小麦品种的选育和新种质的创制提供指导。
随着测序技术的快速发展,具有高通量、自动化程度高的SNP标记芯片在小麦遗传作图与新基因发掘、群体结构分析、比较基因组学研究等方面得到了极大的应用[2-6]。小麦55K SNP芯片是在Affimetrix660K芯片的基础上精选优化开发出来的SNP标记芯片,目前已广泛应用于小麦遗传研究[7]。马艳明等[8]利用小麦55K SNP芯片对188份新疆冬小麦品种进行了遗传多样性分析,发现育成品种与地方品种的遗传差异较大; 水志杰等[9]利用人工合成小麦RIL群体,通过小麦55K SNP芯片对穗长和穗宽等穗部性状进行了QTL发掘,鉴定出具有一因多效QTL位点; Li等[10]通过384份小麦种质和55K SNP芯片对小麦产量相关性状进行了全基因组关联分析,发掘出一些与产量性状相关的优异等位基因位点。这些研究表明, 55K SNP芯片在小麦主要农艺性状遗传解析方面具有良好的应用前景,但迄今较少有研究关注小麦主栽品种的遗传解析。
周麦32号是周口市农业科学院以黄淮麦区主栽品种矮抗58为母本,以优质强筋、高产稳产、抗病、广适的周麦24号为父本,通过有性杂交选育出的高产优质强筋型小麦新品种。该品种能满足市场对优质强筋小麦的需求,市场推广潜力较大。鉴此,本研究通过小麦55K SNP芯片对周麦32号及其亲本进行全基因组扫描,从分子水平上解析周麦32号的遗传构成及亲本对其遗传贡献率,为优质强筋小麦品种的遗传改良和推广应用提供参考。
1 材料与方法
1.1 试验材料
供试材料为周麦32号及其亲本矮抗58、周麦24号,由西北农林科技大学农学院提供。
1.2 小麦55K SNP芯片检测
每个供试品种于3叶期选取长相一致的幼嫩叶片,采用改良的CTAB法[11]提取基因组DNA, 通过10 g·L-1琼脂糖凝胶电泳对样品DNA质量进行检测,利用分光光度计对DNA浓度进行测定。供试材料小麦55K SNP芯片检测由北京博奥晶典生物技术公司完成。利用Excel对周麦32号及其亲本基因型数据进行分析,剔除杂合位点,缺失位点及染色体未知的位点后,最终有48 613个SNP位点用于后续的遗传分析。
1.3 周麦32号亲本遗传贡献率分析及遗传型图谱构建
根据周麦32号亲本间SNP位点的多态性进行遗传贡献率的计算,统计出亲本间多态性的SNP位点,在此基础上若周麦32号的SNP位点与某个亲本一致则表明在该SNP位点上周麦32号继承了该亲本的基因座。亲本的遗传贡献率为某一亲本遗传给周麦32号的SNP位点数与亲本间差异SNP位点总数的百分比。根据所有SNP位点在不同染色体上的位置[12], 利用Excel对每条染色体上SNP位点的高低进行排序,通过GGT 2.0软件[13]绘制周麦32号的基因型图谱。此外,根据SNP标记的参考图谱位置信息[12], 将染色体上连续相邻3个及其以上SNP标记且位置长度大于1 Mb的区段视为一个染色体区段,通过Excel统计双亲在不同染色体上染色体区段的数量。亲本特异性SNP标记基因注释信息参考Sun等[12]对小麦55K SNP标记的基因注释,并通过Excel完成亲本基因注释信息的数量统计,其他相关的图形均通过Origin 2018进行绘制。
2 结果与分析
2.1 矮抗58和周麦24号对周麦32号的遗传贡献率
利用小麦55K SNP芯片对供试材料在全基因组上扫描发现,周麦32号的亲本矮抗58和周麦24号在全基因组范围内有20 652个多态性SNP位点,通过分析亲本的遗传贡献率,结果发现矮抗58和周麦24号特异SNP位点传递给周麦32号的比例分别为73.75%和26.35%, 矮抗58对周麦32号的遗传贡献率远高于周麦24号(图1)。就基因组水平而言,矮抗58在A基因组上对周麦32号的遗传贡献率最高(79.88%), 其次是D基因组(77.29%), 再次是B基因组(64.58%)。在不同染色体上,双亲对周麦32号的遗传贡献差异较为明显,矮抗58在大多数染色体上的遗传贡献率高于周麦24号,尤其是在1D、2A、4D、6B、7A等染色体上的遗传贡献率超过90%, 而周麦24号仅在1B、4B和6A等染色体上的遗传贡献率略高于矮抗58。
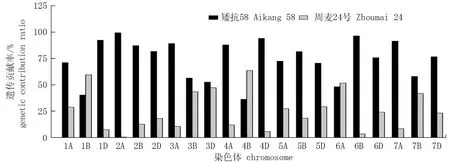
图1 矮抗58和周麦24号在不同染色体上的遗传贡献Fig.1 The genetic contribution of Aikang 58 and Zhoumai 24 on different chromosomes
2.2 周麦32号来源于亲本的染色体区段
根据周麦32号及其亲本SNP分型结果,通过GGT2.0绘制周麦32号的基因型图谱(图2)。在周麦32号的不同染色上均检测到来自其亲本的SNP标记,每条染色体上周麦32号继承双亲的染色体标记的比例均表现不同。周麦32号在1A、1B、1D、2A、3A、4A、5A、6B、7A、7B、7D等染色体上存在来源于矮抗58的较大染色体区段,在1B、3B、4B、5A等染色体上也存在来源于周麦24号的较大染色体区段。通过进一步分析周麦32号获得亲本染色体区段的数量(图3), 发现周麦32号在1A、1D、2A、2B、2D、3A、4A、4D、5B、6B、7A、7D等染色体上存在较多的遗传片段来源于矮抗58, 而1B、4B、6A等染色体上存在较多的染色体区段来源于周麦24号。在3B、3D和7B等染色体上周麦32号来源于双亲的染色体区段较为接近。
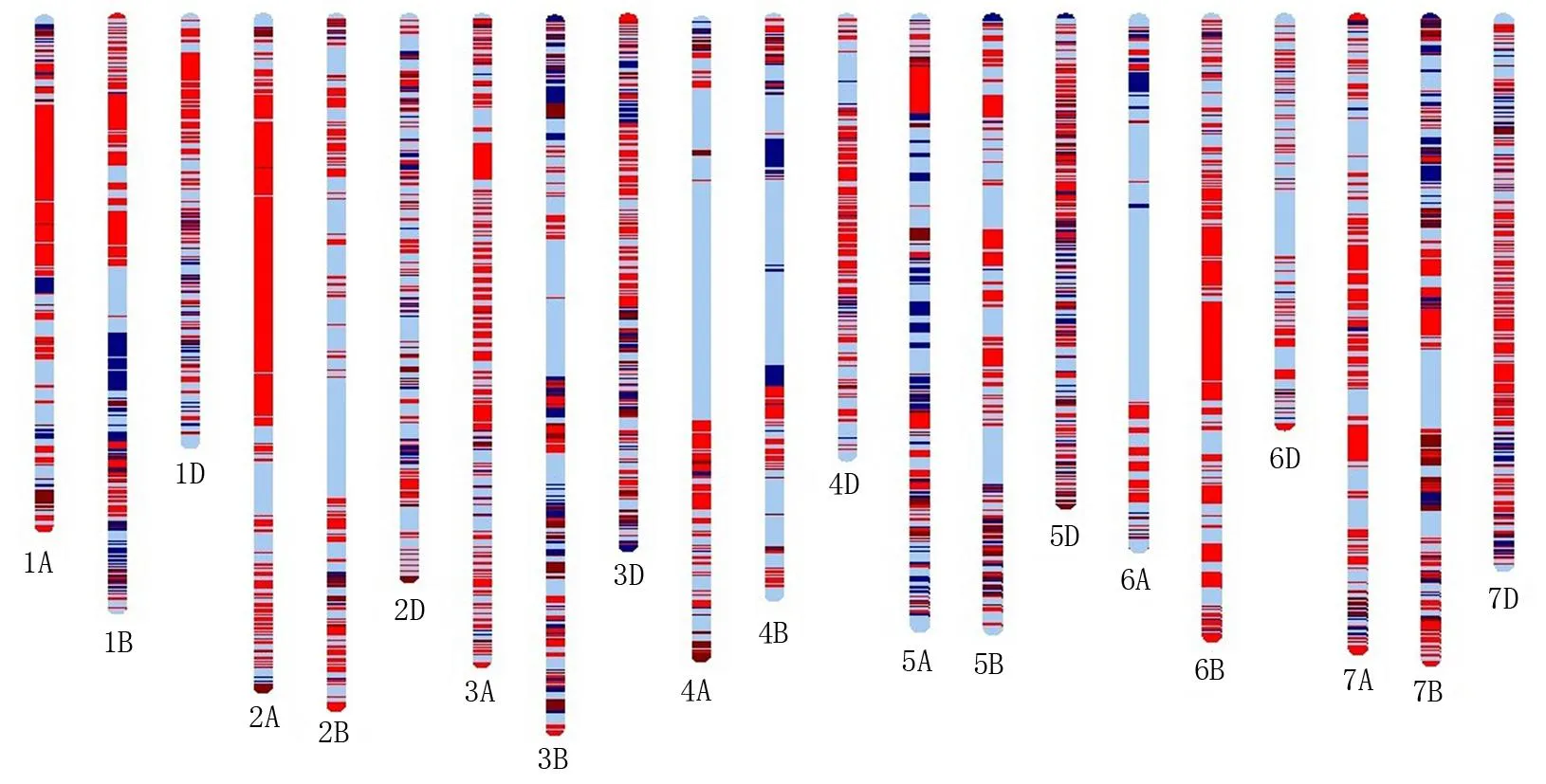
图2 周麦32号与双亲的基因型图谱*Fig.2 SNP genotypic maps of Zhoumai 32 and both parents on different chromosomes* 红色: 矮抗58区段; 深蓝色: 周麦24号区段; 浅蓝色: 矮抗58、周麦24号和周麦32号共有的区段; 褐色: 周麦32号特有区段。* Red: Aikang 58 fragment; Dark blue: Zhoumai 24 fragment; Wathet: identical fragment of Aikang 58, Zhoumai 24 and Zhoumai 32; Brownness: Zhoumai 32 specific fragment.
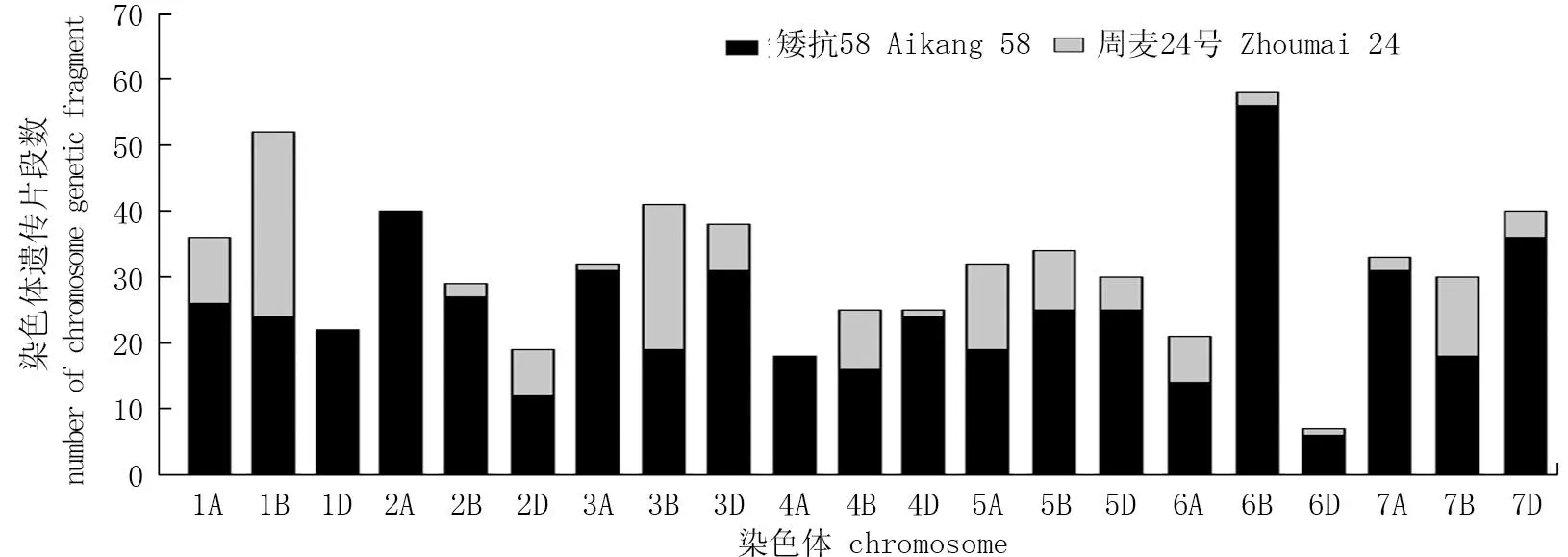
图3 周麦32号来源于双亲染色体区段的数量分布Fig.3 The distribution of genetic fragments from both parents of Zhoumai 32
2.3 周麦32号来源于亲本特异性SNP注释基因
通过对周麦32号及其亲本特异性SNP标记进行基因注释,统计亲本特异性基因注释数量,共提取到4 360个注释基因,矮抗58遗传给周麦32号的特异性注释基因数量为3 188个,占73.12%, 远高于周麦24号注释基因的数量(1 172个)。在不同染色体上,周麦32号获得亲本特异性注释基因的数量差异较大(图4), 但周麦32号在大多数染色体上继承了矮抗58的特异性注释基因,而周麦24号仅在1B和3B染色体上特异性注释基因的数量稍高于矮抗58, 且在2A和4A染色体上检测到的注释基因数量较少,不足10个。
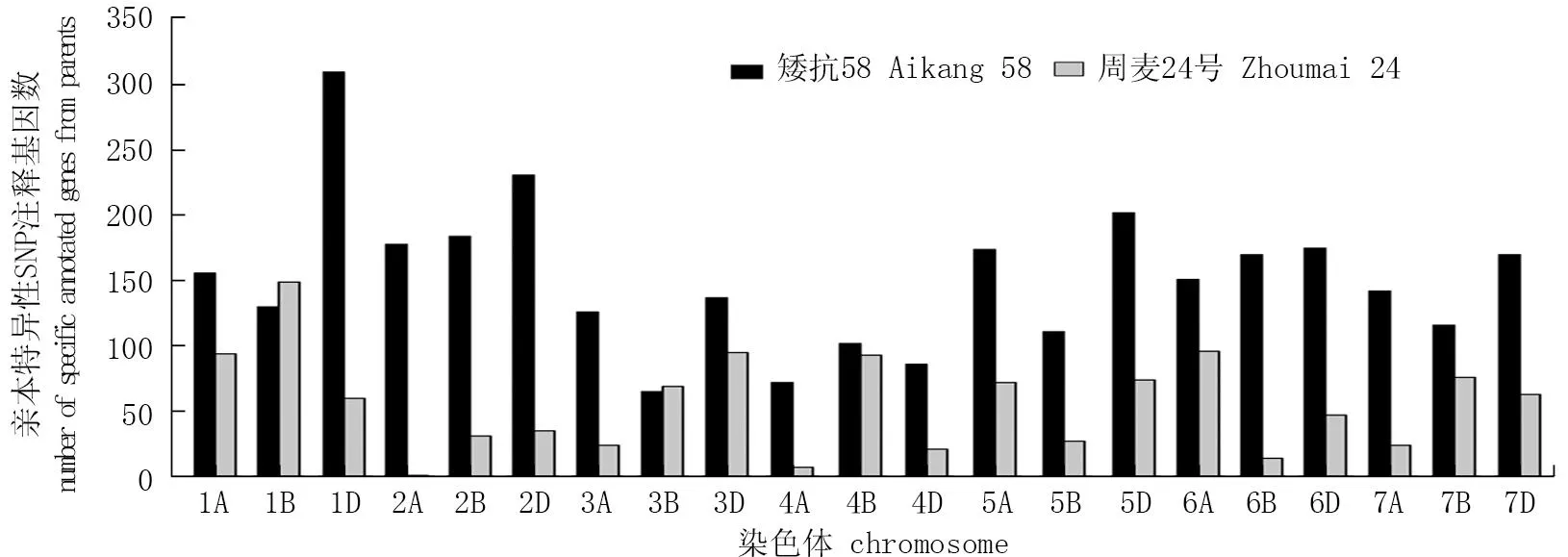
图4 周麦32号双亲特异性SNP注释基因的数量分布Fig.4 The distribution of specific annotated genes originated from both parents of Zhoumai 32
3 讨论
高产、优质、高效是小麦品种遗传改良的主要目标[14], 本地区的骨干亲本或主栽品种是育种家在品种选育过程中倾向选择的亲本资源[15], 选育出的后代品系在主要农艺性状上与这些材料有较大的相似性,多数是在骨干亲本或主栽品种上对目标改良性状的修饰改良,因此小麦品种在选育过程中受到这种定向的人工选择往往会出现不同类型的偏亲现象[16-18]。本研究通过小麦55K SNP标记对周麦32号及其双亲进行标记检测,在全基因组上对周麦32号的遗传构成进行分析,发现矮抗58对周麦32号的遗传贡献率(73.75%)远高于周麦24号(26.25%), 在周麦32号的大多数染色体上矮抗58的遗传贡献率均超过周麦24号,这与杨子博等[19]利用SSR标记对淮麦33的遗传构成分析、孔子明等[20]通过小麦90K SNP标记对周麦16的遗传贡献进行解析以及陈晓杰等[21]利用小麦50K SNP育种芯片对郑品优9号的分子遗传基础进行解析等研究结果较为相似,这表明周麦32号在遗传物质继承上获得了更多矮抗58的遗传物质,表现出明显的偏亲遗传现象。矮抗58是黄淮麦区优异的骨干亲本,该品种高产稳产、矮秆抗倒、适宜种植地区广,也是黄淮麦区的主栽品种[22]; 周麦32号在品种选育过程中,更倾向于对矮抗58的优异性状的选择,遗传了矮抗58更多高产、广适的遗传区段。同时父本周麦24号属于优质强筋、高产稳产、抗病、广适小麦新品种,其抗条锈病和品质性状可能来源于周麦24号。矮抗58和周麦24号均是国内骨干种质周8425B的衍生品种,周麦32号集聚双亲的高产、多抗、优质的优良基因,遗传基础较好。本研究中小麦55K SNP标记在周麦32号及其亲本各染色体上分布较为均匀,能有效地对周麦32号的遗传构成进行分析,标记解析效力较高,而孔子明等[20]在解析周麦16号的遗传构成时因D组染色体标记较少且分布不均匀,标记的解析效力有限。
染色体区段的传递与继承是品种获得主要性状的重要途径,本研究通过对周麦32号亲本特异性染色体区段数量的分析发现,周麦32号获得了更多矮抗58的染色体区段。同样,对周麦32号亲本特异性注释基因分析发现,周麦32号继承了更多矮抗58的特异性注释基因。染色体区段和注释基因分析均表明周麦32号遗传更多矮抗58的基因资源。此外,本研究还检测出较多的周麦32号特异性SNP位点,这些特异的SNP位点可能是品种繁殖过程中异花授粉(小概率发生)、自然突变、亲本间基因重组所带来的。
猜你喜欢
云南画报(2021年11期)2022-01-18
长江蔬菜(2021年12期)2021-04-04
种子(2020年9期)2020-10-22
中国种业(2019年6期)2019-06-25
中国果业信息(2019年11期)2019-01-05
铁道通信信号(2018年8期)2018-11-10
铁道通信信号(2018年8期)2018-11-10
铁道通信信号(2016年4期)2016-06-01
西南农业学报(2016年5期)2016-05-17
