
张家界砂岩峰林顶部植物群落构成及空间分布
2024-01-16 07:15琚玉枫刘鲤邓钰烨田建林
中国野生植物资源 2023年12期
琚玉枫,刘鲤,邓钰烨,田建林
(吉首大学 土木工程与建筑学院,湖南 张家界 427000)
张家界地貌自2010年正式命名以来,一直是地貌学家、地质学家、生物学家们关注的焦点。因其形成年代久远、成土母质独特、小气候特殊等因素造就了独特的植物生境,孕育着丰富的植物资源。人们对张家界地貌区的植物类型、生境状况、生态环境及效应等方面开展了重要的调查研究工作。例如李菁和刘应迪[1]对湘西武陵源地区木本植物区系的组成及其基本特征进行分析与讨论,周大松[2]研究了武陵源世界自然遗产地植物群落特征,周辉[3]研究了武陵源世界自然遗产地物种多样性等,然而相关学者都是从较大尺度对该区域植物进行研究。砂岩峰林是张家界地貌演化过程的产物,由于其顶部人迹罕至、相对封闭独立,目前对峰林顶部植物组成及群落的研究相对较少,又因其特殊生境,开展植物群落的构成及其空间分布调查研究,是对张家界峰林地貌植物资源的保护和持续利用,进而对揭示张家界地貌区植被演替及生态效应等方面具有重要意义。
基于此,本研究以湘西武陵山脉东段的武陵源核心景区为研究区域,依托ArcGIS和无人机(UAV)拍照技术,通过野外调查和相关数理统计软件,拟探讨以下问题:(1)海拔梯度下,张家界砂岩峰林顶部植物组成状况;(2)峰林顶部典型植物群落类型、构成和空间分布。通过解决以上问题,以期查清峰林顶部植物状况,为张家界砂岩峰林植物资源科学保护与利用提供参考。
1 材料与方法
1.1 研究区概况
研究区位于世界自然遗产地武陵源核心景区,地势由西北向东南倾斜,处中亚热带气候带向北亚热带气候带的过渡地区,属亚热带山原型季风湿润气候[4],该区域是峰林地貌典型发育区。
峰林地貌是当地生物多样性的生态关键区[5]。研究区内有石英砂岩峰林3 103 座,峰林独立分布,相对集群且存在相对高度差[6]。峰林是顶部植物的立地条件,为植物生长提供水分、养分、空气和物理支撑。峰林顶部岩石有单一的石英砂岩和石英砂岩夹页岩层两种,由于这两种岩石的岩性不同,在长期流水侵蚀、重力崩塌、风化作用下形态不同,在水平层岩上,多呈柱形或锥形。依据峰林海拔高度和不同海拔的植被类型,可将峰林分为高峰林、中峰林和低峰林。海拔1 000 m以上以针叶林为主的峰林称高峰林;海拔800~1 000 m以针阔混交林为主的峰林称中峰林;海拔800 m以下以常绿阔叶林为主的峰林称低峰林。
1.2 研究方法
1.2.1 野外调查
根据研究区内高、中、低3类峰林的占比情况,兼顾峰林空间分布情况及其顶部可达性,参考研究区地形图、旅游景区分布图和遥感影像数据,把研究区的3 103座峰林作为样本总体,设计概率比例规模抽样方案,辅以UAV、GPS等技术手段,于2021年12月5日至2022年5月15日对研究区内11个区域51个采样地(图1)的153座峰林进行踏查,涉及低峰林63座、中峰林57座、高峰林33座。调查以现场拍照、采样和记录的方式进行,对于无法到达顶部的峰林,则采用UAV、单反相机拍照,望远镜环绕观察记录的方式进行。调查内容包括峰林位置、形状、大小和海拔,峰林顶部植物种类、植物生物学特征、群落结构、环境状况等。
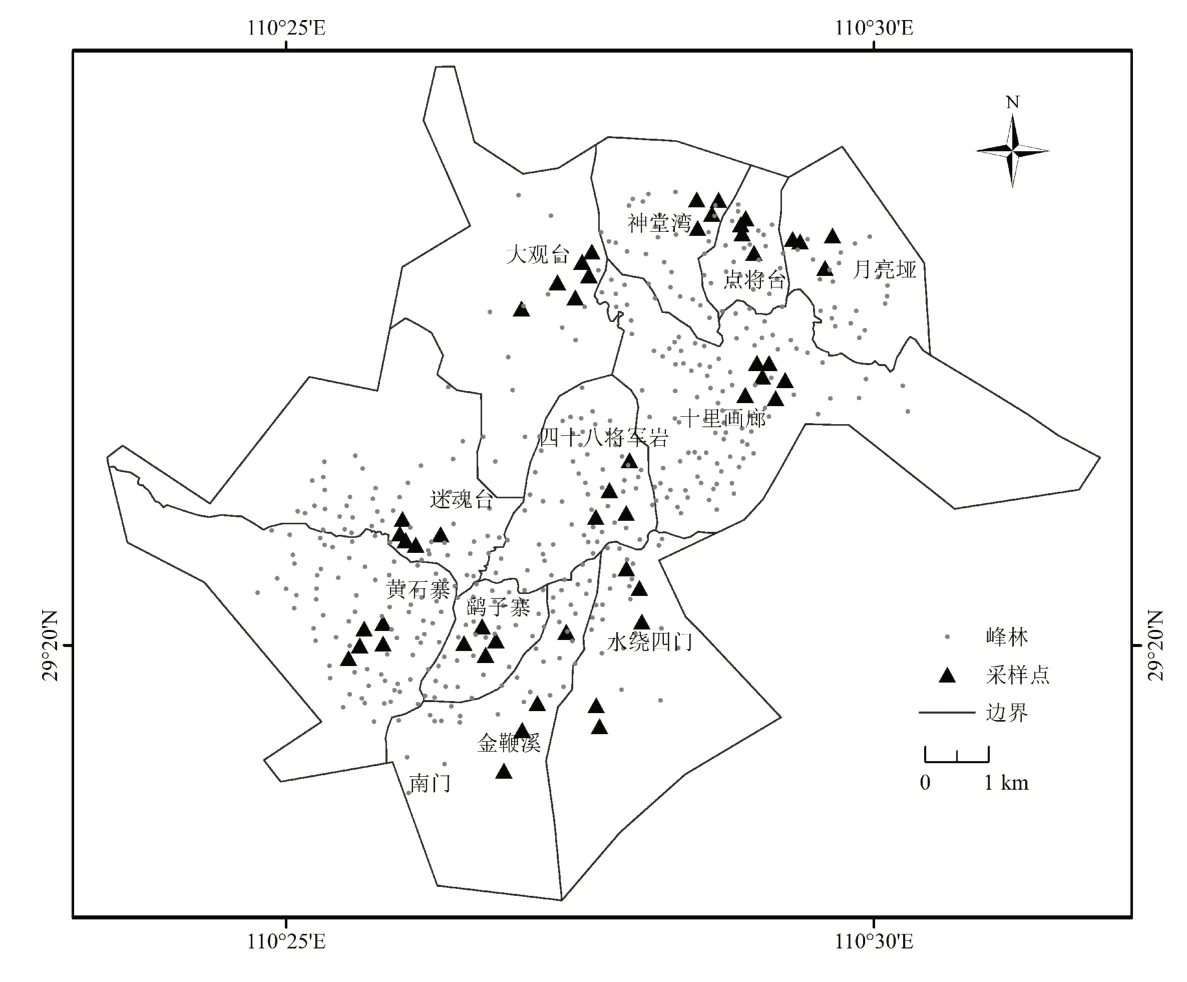
图1 张家界砂岩峰林分布及采样点Fig.1 Distribution and sampling points of the Zhangjiajie sandstone peaks
1.2.2 室内分析
室内分析工作包括植物种类鉴定、群落划分和群落多样性测度[7-9]。
(1)利用峰林顶部植物特写照片和采集到的植物标本,借助计算机画图工具的图片放大功能,参照《湖南植物名录》[10]和中国科学院植物科学数据中心(http://www.plantplus.cn/cn)的植物微细结构描述,结合张家界地区植物种属研究资料[11]进行物种鉴定。
(2)在Excel中将采集的峰柱海拔、坐标和植物种类信息进行标记,进而统计植物科属信息。
(3)通过判读同一植物群落不同角度的图片,确定植物群落的植物构成、结构等,根据植物群落学-生态学原则,以群丛为单位对植物群落进行分类[12],采用“联名法”对群丛进行命名,并在ArcGIS 10.2软件平台绘制峰林顶部典型植物群落空间分布图。
(4)计算β多样性。群落多样性是生物群落的重要特征,反映群落自身特征及其与环境之间的关系。β多样性表示物种沿海拔梯度所发生替代的程度或物种变化的速率,某一环境梯度不同样方之间的共有种越少,β多样性越大,计算公式[13]如下:
其中, Jaccard指数反映群落物种组成的相似性,Cody指数反映物种沿海拔梯度的替代速率,CJ为Jaccard指数,βC为Cody指数,a和b分别为两个海拔梯度的植物种数,c为两个海拔梯度的共有植物种数,g(H)为沿海拔梯度增加的植物种数,l(H)为沿海拔梯度失去的植物种数。
2 结果与分析
2.1 峰林顶部植物状况分析
2.1.1 峰林顶部植物种类与组成
本次调查共鉴定117种峰林顶部植物(表1),分属52科94属,包括蕨类植物6科6属6种,裸子植物2科2属4种,被子植物44科86属107种,单子叶植物3科5属5种,双子叶植物41科81属102种,说明双子叶植物是峰林顶部植物的主要类型。
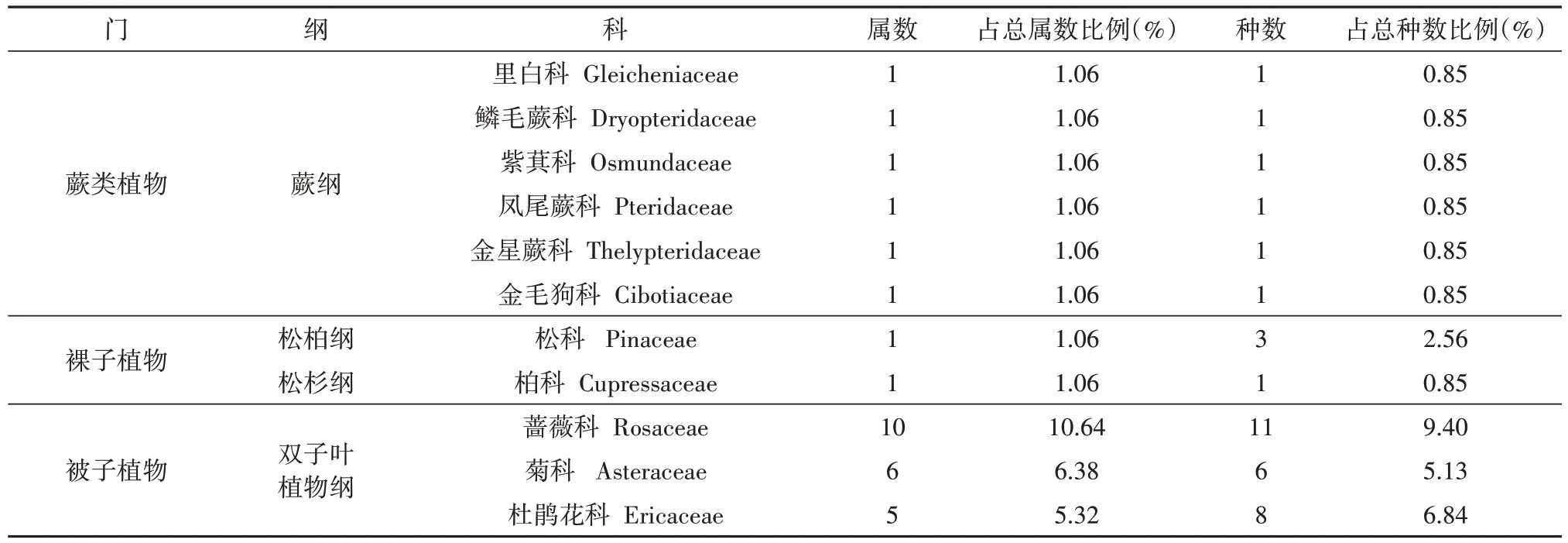
表1 峰林顶部植物状况Tab.1 Statistics of plants at the top of peaks
从科级水平看,种数最多的科为蔷薇科,其次为杜鹃花科和壳斗科,其种数分别为11、8和8种。峰林顶部含1种的单种科有28科,占总科数的54%,占总属数的30%。这反映出峰林顶部部分植物并非以一定亲缘关系群居,而是依据其生物和生态学特性趋同组合[14]。峰林顶部含5种以上的科有5科,包括蔷薇科、杜鹃花科、壳斗科、菊科和樟科,这5科植物占总科数的12%,包含29属39种,占属种总数的31%和33%。
从属级水平看,含1种的属有80属,占总属数的85%;含两种的属有10属,占总属数的11%;含3 ~ 5种的属有3属,占总属数的3%;栎属植物所含种数(6种)最多,占总种数的5%。同属的种具有共同起源和相同进化趋势,其生物学、生态学特征较科更为接近,属比科更能反映植物进化、变异情况,峰林顶部植物优势属现象不明显,属种较为单一,表明峰林顶部进化变异较少,植物古老原始。
2.1.2 海拔梯度下峰林顶部植物组成分析
海拔高度小于800 m的低峰林顶部植物有31科52属63种,海拔在800 ~ 1 000 m之间的中峰林顶部植物有46科75属90种,海拔大于1 000 m的高峰林顶部植物有24科37属45种。中峰林顶部物种数量最多,高峰林顶部物种数量最少,峰林顶部植物科、属、种的数量随海拔升高先增加后减少(图2)。
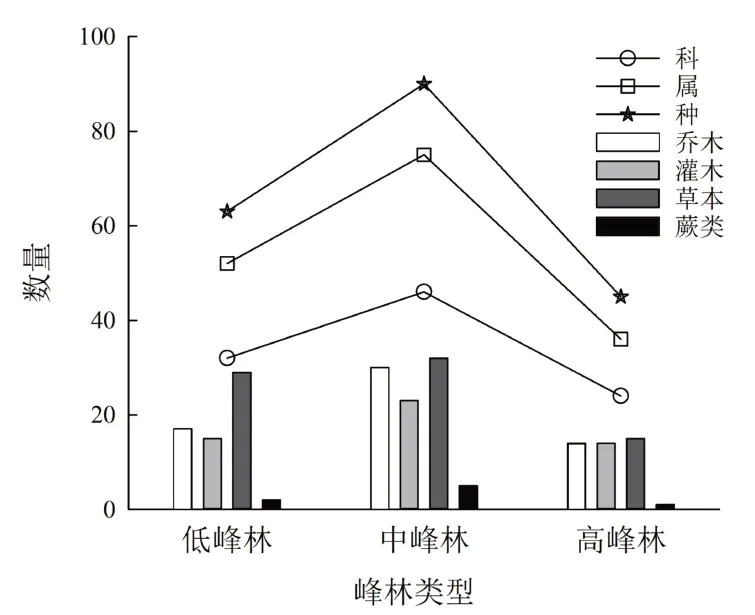
图2 海拔梯度下峰林顶部植物型和植物科、属数量Fig.2 Statistics of plant life form, families and genera on top of peaks with altitude
植物生活型是植物在其系统发育过程中为适应各种环境因素所呈现的综合形态[15]。相同的生活型表明植物对环境具有相同或相似的适应能力[16],海拔对植物生活型具有重要影响。武陵源核心景区峰林顶部植物总体生活型的大小序列为:草本 > 灌木 > 乔木 > 蕨类,各植物型的物种数分别为41、37、34、5,占全部调查植物生活型的比例为35.04%、31.62%、29.06%与4.28%。随着海拔从800 m上升到1 000 m,峰林顶部草本、灌木、乔木、蕨类植物分别从29、15、17、2种先增加到32、30、23、5种后减少到15、14、14、1种(图2)。峰林顶部植物型的数量具有随海拔升高先增加后减少的趋势,草本植物在各海拔梯度中均占有重要地位。
进一步用Jaccard指数和Cody指数比较3个海拔梯度下植物的相似性和植物随海拔梯度替换的速率(表2)发现:各海拔梯度间Jaccard指数在0.35 ~ 0.44时中峰林和高峰林之间的相似性最低,低峰林和中峰林之间的相似性最高。这是因为高海拔峰林顶部气候严酷,太阳辐射强,干旱风大,土壤相对贫瘠,植物生长发育受水分限制;中海拔和低海拔峰林顶部常年云雾缭绕,水汽充沛,植物生长主要受峰顶面积限制;相比之下,中海拔和低海拔峰林顶部更可能为植物生长提供相对优越的水热资源配置[17]。Cody指数数值范围在23.00 ~ 32.50时,中峰林和高峰林间的植物替换率最大,低峰林和高峰林间的植物替换率最小。

表2 海拔梯度下峰林顶部植物相似性和替换率Tab.2 Plant similarity and replacement rate at the top of peaks under altitude gradient
2.2 峰林顶部典型植物群落及分布制图
根据植物群落学 - 生态学原则,采用联名法将峰林顶部植物分为8个典型群落,分析群落组成、结构、分布等(表3)。

表3 峰林顶部典型植物群落组成及分布Tab.3 Composition and distribution of typical plant communities at the top of peaks
(1)武陵松群丛。在武陵源风景区的各区域均有分布。群落植物以武陵松(Pinus massonianavar.wulingensisC. J. Qi et Q. Z. Lin)为主,组成群落的植物种类少,群落结构简单。群落外貌主要由武陵松的生长状况决定,当水热充足时,藓类先锋植物会附着在峰林顶部和岩壁,与武陵松共同组成植物群落。
(2)武陵松 - 鹿角杜鹃 - 芒群丛。主要分布在黄石寨、鹞子寨、金鞭溪、月亮垭、点将台、神堂湾和大观台7个区域。主要乔木有武陵松,伴生灌木鹿角杜鹃(Rhododendron latoucheaeFranch.)和草本植物芒(Miscanthus sinensisAnderss.)、艾(Artemisia argyiLévl. et Van.)、野茼蒿[Crassocephalum crepidioides(Benth.) S. Moore]及苔藓等。
(3)乌冈栎 - 银木荷群丛。主要分布在黄石寨、金鞭溪、水绕四门、十里画廊和迷魂台5个区域。乔木有乌冈栎(Quercus phillyreoidesA. Gray)、银木荷(Schima argenteaPritz. ex Diels);伴生灌木有山茶(Camellia japonicaL.)、鹿角杜鹃、穗序鹅掌柴(Heptapleurum delavayiFranch.)、青榨槭(Acer davidiiFranch.)等;草本植物有紫金牛[Ardisia japonica(Thunberg) Blume]、拉拉藤(Galium spuriumL.)、直刺变豆菜(Sanicula orthacanthaS. Moore)、竹叶草[Oplismenus compositus(L.) Beauv.]、大叶苎麻[Boehmeria japonica(Linnaeus f.) Miquel]等。
(4)武陵松 - 乌冈栎 - 鹿角杜鹃群丛。主要分布在四十八将军岩、鹞子寨、水绕四门、十里画廊、点将台和神堂湾6个区域。乔木层有武陵松、乌冈栎;伴生灌木有鹿角杜鹃、西南红山茶(Camellia pitardiiCoh. St.)、青灰叶下珠(Phyllanthus glaucusWall. ex Muell. Arg)、矩叶鼠刺(Itea omeiensisC. K.Schneider)、小果珍珠花[Lyonia ovalifoliavar.elliptica(Sieb. et Zucc.) Hand. -Mazz.]、广东紫珠(Callicarpa kwangtungensisChun)等;草本植物相对丰富,有多花黄精(Polygonatum cyrtonemaHua)、白头婆(Eupatorium japonicumThunb.)、鸭跖草(Commelina communisL.)、龙牙草(Agrimonia pilosaLdb.)、南方露珠草(Circaea mollisSieb. et Zucc.)、紫萁(Osmunda japonicaThunb.)、柄花天胡荽(Hydrocotyle himalaicaP. K. Mukherjee)等。
(5)武陵松 - 马尾松 - 交让木群丛。主要分布在黄石寨、迷魂台、鹞子寨、神堂湾和点将台5个区域。乔木层有武陵松、马尾松(Pinus massonianaLamb.)、交让木(Daphniphyllum macropodumMiq.)、樟(Camphora officinarumNees ex Wall.)等,灌木有鹿角杜鹃、马银花[Rhododendron ovatum(Lindl.)Planch.],伴有三脉紫菀(Aster ageratoides Turcz.)、一年蓬[Erigeron annuus (L.) Pers.]、蛇含委陵菜(Potentilla kleinianaWight et Arn.)、悬铃叶苎麻(Boehmeria platanifoliaFranchet & Savatier)等草本植物。
(6)曼青冈 - 山茶群丛。主要分布在神堂湾、点将台、大观台和月亮垭4个区域。乔木层有曼青冈(Quercus oxyodonMiq.)、樟等,伴生灌木山茶、鹿角杜鹃、猫儿刺(Ilex pernyiFranch.)、马银花等;林下草本植物有芒、杏叶沙参[Adenophora petiolatasubsp.hunanensis(Nannfeldt) D. Y. Hong & S. Ge]、三脉紫菀等。
(7)柳杉 - 鹿角杜鹃群丛。主要分布在金鞭溪、十里画廊和四十八将军岩3个区域。群落乔木层有檫木[Sassafras tzumu(Hemsl.) Hemsl.]、樟、虎皮楠[Daphniphyllum oldhamii(Hemsley) K. Rosenthal]等;灌木层有鹿角杜鹃、绣线梅(Neillia thyrsifloraD. Don)、香粉叶(Lindera pulcherrimavar.attenuataAllen)等;草本植物有尖裂假还阳参[Crepidiastrum sonchifolium(Maxim.) Pak & Kawano]、艾、野茼蒿、三脉紫菀等。
(8)枹栎 - 樟 - 柯群丛。主要分布在黄石寨、鹞子寨、金鞭溪、水绕四门和迷魂台5个区域。乔木层有枹栎(Quercus serrataMurray)、樟、柯[Lithocarpus glaber(Thunb.) Nakai]、交让木、肉桂(Cinnamomum cassiaPresl)、构树[Broussonetia papyrifera(L.)L′Hér. ex Vent.]、刺槐(Robinia pseudoacaciaL.)等;灌木层有马银花、锦绣杜鹃(Rhododendron × pulchrumSweet)、鹿角杜鹃等;林下草本植物有野茼蒿、一年蓬、苎麻[Boehmeria nivea(L.) Gaudich.]、艾、狗肝菜[Dicliptera chinensis(L.) Juss]等。
3 讨论
物种组成是植物群落基本特征之一。本次调查中,武陵源核心景区峰林顶部植物共有52科94属117种,包括裸子植物2科2属4种,被子植物44科86属107种,蕨类植物6科6属6种,其中以被子植物中的双子叶植物占优。从物种数量来看,与亚热带地区的云南石林核心区栽植植被样地中木本植物50科101属138种[18]、武夷山国家公园调查样地中维管植物89 科 228属450种[19]、广州象头山国家级自然保护区中兰科植物33属48种相比[20],由于峰林顶部面积较小,植物种类总体上相对丰富。
海拔梯度影响温度、光照和水分等多种环境因子变化,从而直接或间接影响植物群落组成、结构。植物种数的海拔梯度格局常常与研究区域的地理位置、环境条件和尺度有关。在本研究中,峰林顶部植物种数随海拔先增加后减少,低海拔区域植物的相似性高而替换率低,高海拔区域植物的相似性低而替换率高。黄炎根等[21]研究湖南张家界砂岩峰林不同海拔峰林林下植物多样性得出,不同海拔灌木层与草本层的丰富度和多样性表现为随着海拔的增加呈现先增大后减小再增大的趋势。朱家兴等[22]在研究武陵源区崖壁植物多样性中也得出,崖壁植物物种数目与海拔高度变化服从正态分布,物种数量随海拔的升高而下降,在700 ~ 800 m达到峰值。陈水飞等[19]也证实海拔与武夷山国家公园植物群落物种多样性之间存在一定规律,不同海拔区间植物群落的物种相似性差异较大。
受限于调查方法的可达性,现有峰林顶部植物调查均限于观景台附近的小规模研究,尚未开展变化海拔的峰顶植物调查,峰顶植物类型随海拔的变化规律也尚无科学定论。目前,国内外专家学者已利用UVA进行过喜马拉雅植被图的绘制[23],也利用无人机进行崖壁植物空间分布特征的研究[24]。峰林顶部植物具有不可及和不易及的特点,调查面临着高风险、高成本的难题,但UVA辅助技术在一定程度上为进行植物调查提供可能。
4 结论
张家界砂岩峰林顶部植物种类丰富;峰林顶部植物型,植物科、属、种的数量随海拔升高先增加后减少;植物群落构成、分布存在空间差异。开展峰林顶部植物种类、群落类型和群落分布状况的调查研究,对砂岩峰林植物资源保护和利用具有一定科学意义。由于采集数据有限,无法对研究区所有植物及群落状况进行分析。对造成峰林顶部植物群落组成、群落结构特征差异的机理尚未深入讨论,今后可在这方面做进一步研究。
猜你喜欢
东北林业大学学报(2023年2期)2023-02-22
水土保持研究(2022年1期)2022-12-27
岷峨诗稿(2019年4期)2019-04-20
乡村地理(2018年1期)2018-07-06
下一代英才(酷炫少年)(2017年12期)2018-01-19
乡村地理(2017年4期)2017-09-18
商业文化(2017年23期)2017-04-23
中国老区建设(2016年1期)2016-02-28
乡村地理(2015年3期)2015-11-10
山西大学学报(自然科学版)(2014年2期)2014-05-10
