
甲醛足底致痛大鼠脊髓后角神经元内蛋白激酶C活性改变对一氧化氮合酶的影响
2011-06-08 02:10吕兴业
河北医药 2011年17期
吕兴业
热痛觉过敏的产生与一氧化氮(nitric oxide,NO)生成有关。虽然痛觉过敏机制尚未搞清,但目前认为在伤害性因素刺激下,脊髓伤害性信息传入神经元释放性氨基酸(excitatory amino acids,EAAs),激活 N-甲基-D-天冬氨酸(CN-methyl-D-aspartic acid,NMDA)受体。NMDA受体激活一方面可引起蛋白激酶(protein kinasec,PKC)激活;另一方面,NMDA受体的激活,也可引起一氧化氮合酶(nitric oxide synthase,NOS)的激活并生成NO。但目前尚无直接证据表明NMDA受体激活后,PKC激活是否影响NOS的活性,即NOS除受NMDA受体直接调控外,是否也受到PKC活性的调控。本实验旨在观察PKC兴奋剂与抑制剂对甲醛炎性痛及痛觉过敏中脊髓后角诱生型一氧化氮合酶(inducible nitric oxide synthase,iNOS)活性的影响,以阐明疼痛及痛觉过敏时脊髓后角神经元内PKC活性是否影响NO的生成。
1 材料与方法
1.1 实验动物及分组 健康纯种Sprague-Dawley(SD)大鼠54只,由河北医科大学实验动物中心提供,体重260~280 g,雌雄不限,分9组,每组6只。(1)正常组:不加任何处理,取材固定观察;(2)甲醛12 h组:右后掌注射甲醛后12 h取材观察;(3)甲醛24 h组:右后掌注射甲醛后24 h取材观察;(4)甲醛12 h加0.9%氯化钠溶液组:于右后掌注射甲醛前30 min鞘内注射0.9%氯化钠溶液1次,6 h再重复注射1次,注射甲醛后12 h取材观察;(5)甲醛24 h加0.9%氯化钠溶液组:于右后掌注射甲醛前30 min鞘内注射0.9%氯化钠溶液1次,12 h再重复注射1次,注射甲醛后24 h取材观察;(6)甲醛12 h加PMA组:于右后掌注射甲醛前30 min鞘内注射佛波醇脂(Phobol-12-myristate-13-acetate,PMA)1次,6 h再重复注射1次;注射甲醛后12 h取材观察;(7)甲醛24 h加PMA组于右后掌注射甲醛前30 min鞘内注射PMA 1次,12 h再重复注射1次,注射甲醛后24 h取材观察;(8)甲醛12 h加灯盏花素乙(Chelerythrine chloride,CH)组:于右后掌注射甲醛前30 min鞘内注射CH 1次,6 h再重复注射1次,注射甲醛后12 h取材观察;(9)甲醛24 h加CH组:于右后掌注射甲醛前30 min鞘内注射CH 1次,12 h再重复注射1次,注射甲醛后24 h取材观察。
1.2 观察指标 机械与热痛阈。脊髓后角免疫组织化学NOS阳性细胞数量;脊髓后角免疫组织化学NOS阳性细胞染色深度变化;脊髓后角免疫组织化学NOS阳性神经纤维染色深度变化。
1.3 仪器 微量进样器(上海医用激光仪器厂,规格10 μl);Olympus光学显微镜(日本Olympus光学工业株式会社,型号IX71);XSJ-2实验室显微镜(重庆光学仪器厂);显微摄像装置(s^msung);切片机(上海医疗器械总厂,型号DPQ-9010);电子Von Frey测痛仪(2390系列,美国IITC公司,Woodland Hills);热敏测试仪BME-410C(中国医学科学院生物医学研究所,天津)。
1.4 试剂 PMA;CH;甲醛溶液(formaldehyde solution,石家庄市有机化工厂);标本固定液[磷酸盐缓冲液(phosphate buffer saline,PBS)加10%甲醛,河北医大病理实验室];DAB(diaminobenzidine,二氨基联苯胺)显色系统(北京中山生物技术有限公司);兔抗鼠iNOS多克隆抗体(第一抗体,北京中山生物技术有限公司);通用型工作液(第二抗体,北京中山生物技术有限公司);辣根过氧化物酶标记的链霉素卵白素工作液(北京中山生物技术有限公司)。
1.5 实验程序及方法 将实验动物置于有锯末覆盖盒底的鼠盒内,给予充足的水和饲料,并在实验室内饲养2~3 d,动物适应实验室环境后开始实验,然后在预定的时间内取材,制备组织切片,进行预定指标的观察。
1.5.1 炎性疼痛模型:大鼠右后掌底面皮下注射5%甲醛0.2 ml。
1.5.2 鞘内注射:鉴于传统的枕骨大孔插管留置给药法操作难度大、干扰因素多等,参照Terence[1]所建立的方法,大鼠用乙谜作短暂麻醉后,在L4~5腰椎直接穿刺。鞘内注射采用微量进样器进行。穿刺针用2 ml或5 ml注射器针头烧去底座制成。穿刺针与微量进样器之间用拉制的细塑料管(直径小于0.5 mm)相连。先用已连接好的微量进样器抽取 PMA(0.136 nmol)或 CH(0.324 nmol)至 10 μl刻度,然后从大鼠L4~5穿刺进入椎管,以有轻度脊髓刺激征为标志,固定穿刺针,缓慢推入药液,时程不少于1 min。该操作法为本实验首创。
1.5.3 取材:在1%戊巴比妥钠(pentobarbital sodium)腹腔麻醉(40 mg/kg)下进行。逆行(因椎板呈叠瓦式结构)打开椎板,暴露脊髓,末根肋骨下为第1腰椎,依次向下找到L5,在椎间孔处找到L5神经节,从神经节处沿后根向上找到此后根相连的脊髓节段即为该神经节所对应的腰髓。在L5后根的上下端根丝处切断脊髓,取出L5节段,置于同定液中24 h。
1.5.4 iNOS免疫组织化学法:先将标本梯度乙醇脱水:70%乙醇4 min→80%乙醇5 min→90%乙醇5 min→95%乙醇10 min→100%乙醇(无水乙醇)10 min,其中100%乙醇2次。2次二甲苯(dimethyl benzene)透明,石蜡包埋。取脊髓石蜡切片,厚度5 μm,用兔抗大鼠iNOS多克隆抗体为第一抗体,生物素标记的羊抗兔抗体为第二抗体,辣根标记的链霉素卵白素为第三抗体来检测脊髓组织的iNOS。步骤:石蜡切片用二甲苯及系列乙醇脱蜡至水→PBS冲洗5 min→3%过氧化氢室温孵育10 min,灭活内源性过氧化酶→PBS冲洗5 min→10%正常山羊血清(PBS)稀释封闭,室温孵育10 min→滴加1∶50兔抗大鼠iNOS一抗,4℃冰箱过夜→PBS冲洗,5 min×3次→滴加生物素标记的二抗,37℃孵育30 min→PBS冲洗,5 min×3次→滴加辣根过氧化物酶标记的链霉素卵白素→37℃孵育30 min→PBS冲洗,5 min×3次→DAB显色→自来水冲洗,终止显色→苏术素复染,脱水、封片(用DAB显色,阳性信号呈黄色或棕褐色)。
1.6 观察指标
1.6.1 疼痛行为学检测:①机械痛觉过敏测量:实验动物测试在一组22 cm×22 cm×23 cm的笼中进行,动物适应1 h处于安静状态后,用测痛仪刺大鼠足底柔软的部分,2~3 s,出现缩足反应时的读数为机械痛痛阈。每次测试至少间隔5 min,重复3次取平均值。左右足分别测定作为自身对照。②热痛觉过敏试验:在所有机械痛觉过敏试验结束后,大鼠放入测试笼中,底部换成3 mm透明玻璃,适应30 min,待大鼠安静不再走动时开始测试。用热敏测试仪照射大鼠足底,记录开始照射至出现缩足逃避反应的潜伏期时间。光辐照强度先用8只未经任何处理的空白对照大鼠调定至潜伏期(15±3)s,cut-off时间设定为30 s以免造成大鼠足底部损伤,每只大鼠测试间隔10 min以上,测试3次取平均值。
1.6.2 免疫组织化学结果观测:于光学显微镜下进行定性观察。采用ZL-2000型细胞分析系统进行定量分析,包括iNOS阳性细胞数量、阳性细胞及背景的染色深度。后者用吸光度来表示。吸光度(A值)=1 g空白灰度值/标本灰度值,吸光度越大,染色越深。
1.7 统计学分析应用SPSS 13.0统计软件,计量资料以表示,组间比较、行样本均数采用非配对t检验,P<0.05为差异有统计学意义。
2 结果
2.1 痛觉比较 足底注射甲醛后1~2 min,动物即出现疼痛相关反应如舔、摇动后掌、后掌抬离盒面等。30 min后注射部位出现炎性反应,如红、肿等。炎性反应于注射甲醛后24 h左右达高峰。各试验组均出现相应的行为学变化。见表1。
鞘内注射PMA后3~5 min后,动物出现烦躁不安,搔抓盒底,舔咬食物及其它物品等,个别大鼠发出尖叫,互相撕咬。鞘内注射CH 30 min后,动物运动减少,安静卧于盒底。各试验组均出现相应的行为学变化。见表1。
2.2 免疫组织化学比较 L5脊髓后角NOS阳性细胞多呈菱形,有些为圆形或椭圆形,部分细胞可见明显突起,提示为神经细胞。正常组中iNOS阳性细胞数目较少,染色较浅。此外,还可见交错的阳性-染色神经纤维(图1)。甲醛加0.9%氯化钠溶液组与甲醛组差异无统计学意义(P>0.05)。见表2,图2~5。与正常组比较,甲醛12 h组及24 h组L5脊髓后角iNOS阳性细胞数均显著增加(P<0.01),以24 h增加更明显,且细胞及神经纤维染色深度也明显增加(P<0.01)。见表2,图1~3。与甲醛12 h组比较,甲醛12 h加PMA组iNOS阳性细胞数明显增加(P<0.01),细胞及神经纤维染色也明显加深(P<0.01)。见表2,图4、5,接近于甲醛24 h水平,差异无统计学意义(P >0.05)。见表2,图6;甲醛12 h加CH组iNOS阳性细胞数明显减少(P<0.01),细胞及神经纤维染色明显变浅(P <0.01)。见表2,图4、7。与甲醛24 h比较,甲醛24 h加PMA组iNOS阳性细胞数增加不明显(P>0.05),细胞及神经纤维染色也无明显加深(P >0.05)。见表2,图5、8甲醛24 h加CH组NOS阳性细胞明显减少(P<0.01)。细胞及神经纤维染色也明显变浅(P <0.01)。见表2,图6、9。此外,各试验组变化结果注射甲醛侧均高于对侧。
表1 大鼠右前爪注射甲醛再分别鞘内注射PMA或CH后机械痛阈和热痛阈比较n=6,

表1 大鼠右前爪注射甲醛再分别鞘内注射PMA或CH后机械痛阈和热痛阈比较n=6,
注:与正常组比较,*P <0.01;与甲醛12 h组比较,#P <0.01;与甲醛24 h组比较,△P <0.01
组别 机械痛阈(g)热痛阈(s)左右左右64.8 ±3.7 63.9 ±3.4 15.8 ±1.4 15.6 ±1.8甲醛12 h 组 39.7 ±2.4* 37.6 ±2.2* 10.7 ±1.9* 10.2 ±1.5*甲醛24 h 组 17.6 ±1.8* 16.9 ±1.6* 6.3 ±2.1* 5.9 ±1.7*甲醛12 h+0.9 氯化钠溶液组 28.9 ±2.8 27.4 ±2.5 11.4 ±2.2 10.9 ±1.9甲醛24 h+0.9 氯化钠溶液组 17.9 ±1.4 16.7 ±1.6 6.4 ±2.7 6.8 ±2.0甲醛12 h+PMA 组 18.1 ±1.7# 17.6 ±2.1# 5.7 ±1.4# 4.9 ±1.8#甲醛24 h+PMA 组 14.4 ±1.6 13.2 ±2.2 5.2 ±1.5 5.0 ±1.4甲醛12 h+CH 组 57.3 ±4.8# 55.2 ±5.2# 14.8 ±2.3# 14.5 ±1.8#甲醛24 h+CH 组 49.6 ±3.4△ 47.5 ±2.9△ 10.8 ±2.3△ 10.6 ±2.1正常组△
表2 大鼠右前爪注射甲醛再分别鞘内注射PMA或CH后脊髓L5水平NOS阳性细胞数、阳性细胞及背景染色深度(吸光度)值比较n=6,

表2 大鼠右前爪注射甲醛再分别鞘内注射PMA或CH后脊髓L5水平NOS阳性细胞数、阳性细胞及背景染色深度(吸光度)值比较n=6,
注:与正常组比较,*P <0.01;与甲醛12 h 组比较,#P <0.01;与甲醛24 h 组比较,△P <0.01;与左侧比较,☆P <0.01
组别 阳性细胞数 阳性细胞吸光度(×10-3)背景吸光度(×10-3)左右左右左右正常组 8.2 ±1.3 11.2 ±1.7 208.3 ±2.4 214.4 ±2.6 131.3 ±2.2 129.6 ±1.9甲醛12 h组 15.2 ±1.6* 18.6 ±1.1*☆ 226.1 ±2.9* 247.4 ±2.7*☆ 158.8 ±2.1* 183.8 ±2.8*☆甲醛24 h组 21.7 ±2.3* 32.4 ±1.4*☆ 255.8 ±1.9* 284.7 ±2.7*☆ 177.4 ±2.4* 198.4 ±2.2☆甲醛12 h+0.9氯化钠溶液组 14.3±1.6 19.4±1.9☆ 247.2±2.8 254.2±2.3☆ 168.4±2.1 171.7±1.5☆甲醛24 h+0.9氯化钠溶液组 22.1±1.7 23.4±1.6☆ 247.2±2.9 259.2±2.4☆ 156.4±2.5 172.8±1.8☆甲醛12 h+PMA 组 21.0 ±1.8# 28.5 ±2.3#☆ 268.2 ±1.7# 295.1 ±2.1#☆ 173.7 ±2.3# 182.3 ±2.9#☆甲醛24 h+PMA 组 23.6 ±1.7 35.2 ±2.1☆ 268.5 ±1.9 289.4 ±2.7☆ 179.3 ±2.4 199.3 ±1.6☆甲醛12 h+CH 组 11.5 ±1.7# 13.5 ±1.8#☆ 212.3 ±1.3# 230.6 ±2.8#☆ 139.8 ±2.2# 141.8 ±1.9#☆甲醛24 h+CH 组 13.2±1.4△ 19.5±1.5△☆ 219.7±1.7△ 238.6±2.0△☆ 137.2±1.7△ 141.8±2.2△☆
3 讨论
外周伤害信息传入可导致疼痛及痛觉过敏。研究表明,痛及痛觉过敏的产生,与脊髓后角神经元内NMDA受体的激活及由此引起的细胞内PKC激活及NO生成增多有关。结扎大鼠坐骨神经引起的热痛觉过敏,其脊髓内PKC活性明显升高[2]。大鼠后掌皮下注射蜂毒引起的痛觉过敏可被PKC抑制剂CH所减弱[3]。PKC参与了前列腺素2(prostaglandin E-2,PGE2)诱导的机械性痛觉过敏[4]。Ca2+依赖性第二信使PKC激活可加剧炎性大鼠的伤害性反应,可能与脊髓sigma-1受体激活有关[5]。事先抑制大鼠大脑内PKC和PKA可阻止吗啡耐受作用(morphine tolerance,是指长时间使用吗啡后,其镇痛作用逐渐减弱以至消失,但其具体机制仍不清楚)[6]。以上说明了PKC参与了热痛觉过敏的形成。鞘内注射亚硝基化合物-18[2,2-(hydroxynitrosohydrazino)bis-ethanamine,NOC-18,一种 NO 供体],可使结扎坐骨神经所造成的热痛觉过敏的开始时间提前,并加速痛觉过敏的发展[7]。经腹腔、静脉或口服给小鼠一氧化氮合酶(nitric oxide synthase,NOS)抑制剂硝基左旋精氨酸甲酯(NG-nitro-L-arginine methylester,L-NAME),在 甲 醛 足 底 注射[8]、坐骨神经结扎[9]所致的疼痛模型上,均表现出明显而持久的抗伤害性作用。免疫组织化学证实NO供体硝普钠可使大鼠三叉神经脊束核(spinal trigeminal nucleus,STN)NOS阳性细胞增多,染色加深[10]。三种 NOS:诱生型(induciable NOS,iNOS)、神经型(neuronal NOS,nNOS)、内皮型(endothelial NOS,eNOS)在辣椒素诱发咬肌痛觉过敏(capsaicin-induced masseter hypersensitivity)中表达上调[11]。
皮下注射甲醛为常用的化学性致痛和痛觉过敏的方法。

图1 iNOS阳性细胞数目较少,染色较浅,可见交错的阳性染色神经纤维,左右无差别(免疫组化×200)

图2 甲醛12 h组,iNOS阳性细胞数目较正常组明显增多,染色加深,神经纤维染色加深,右侧略强于左侧(免疫组化×250)

图3 甲醛24 h组,iNOS阳性细胞数目较甲醛12 h明显较多,染色加深,神经纤维染色加深,右侧强大左侧(免疫组化×400)

图4 甲醛12 h+0.9氯化钠溶液组与甲醛12 h组接近,无明显变化(免疫组化×400)

图5 甲醛12 h+PMA组iNOS阳性细胞数目较甲醛12 h组明显增多,染色加深,神经纤维染色加深,右侧略强于左侧(免疫组化×400)

图6 甲醛24 h+0.9氯化钠溶液组与甲醛24 h组接近,无明显变化(免疫组化×400)
鞘内注射PMA后3~5 min后,动物出现烦躁不安,搔抓盒底,舔咬食物及其他物品等,个别大鼠发出尖叫,互相撕咬,这说明PMA对大鼠有强烈的致痛和致痛觉过敏作用。鞘内注射CH 30 min后,动物运动减少,安静卧于盒底。这说明CH对痛及痛觉过敏有明显的抑制作用。

图7 甲醛12 h+CH组iNOS阳性细胞数目较甲醛12 h组明显减少,染色变浅,神经纤维色变浅,左侧弱于右侧(免疫组化×400)
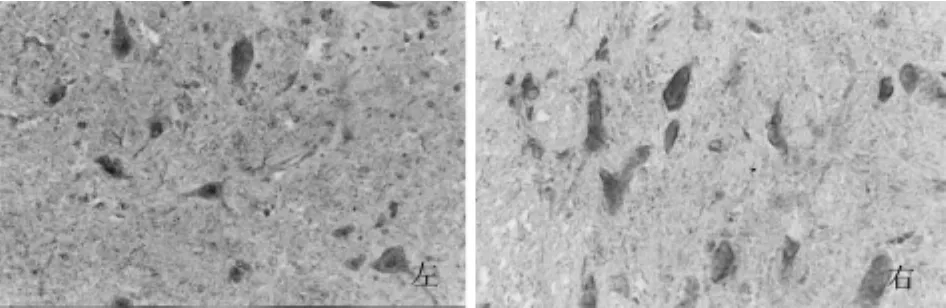
图8 甲醛24 h+PMA组iNOS阳性细胞数较甲醛24 h明显增多,染色加深,神经纤维染色加深,右侧强于左侧(免疫组化×400)

图9 甲醛24 h+CH组iNOS阳性细胞数目较甲醛24 h组明显减少,染色变浅,神经纤维色变浅,左侧略弱于右侧(免疫组化×400)
本研究观察PKC激动剂及抑制剂对甲醛炎性大鼠脊髓后角神经元iNOS活性的影响。发现L5脊髓后角iNOS阳性细胞多呈菱形,有些为圆形或椭圆形,部分细胞可见明显突起,提示为神经细胞。正常组中NOS阳性细胞数目较少,染色较浅。此外,还可见交错的阳性染色神经纤维,这说明正常神经组织中在生理状态下即有一定的NOS活性。
根据已有资料,脊髓后角的iNOS表达在甲醛注射12 h后明显增加,24 h达到高峰,72 h恢复正常[12]。本实验选择注射甲醛后12 h及24 h 2个时间点为鞘内注射PMA和CH的对照组,来观察激动与抑制PKC对脊髓NOS的影响。
与正常组比较,甲醛12 h组及24 h组机械痛阈与热痛阈明显降低,以24 h降低更明显。与甲醛组比较,鞘内注射PMA组机械痛阈与热痛阈降低,以12 h更明显。鞘内注射CH组机械痛阈与热痛阈明显升高。
与正常组比较,甲醛12 h组及24 h组L5脊髓后角iNOS阳性细胞数均显著增多,以24 h增加更明显,且阳性细胞及神经纤维染色深度也明显增加。与甲醛组比较,鞘内注射PMA组iNOS阳性细胞数明显增多,细胞及神经纤维染色也明显加深,以12 h更明显。这表明激动PKC能明显增加iNOS的活性。鞘内注射CH组iNOS阳性细胞数明显减少,细胞及神经纤维染色明显变浅,这表明抑制PKC能明显降低iNOS的活性。本实验结果表明,在甲醛所致的炎性痛及痛觉过敏中,iN-OS活性与PKC活性有关,PKC激活可促进iNOS进一步激活。
已有实验表明,在甲醛炎性疼痛及痛觉过敏时,脊髓后角神经元内有PKC激活,并有NOS的活性增强,二者都与痛及痛觉过敏的产生有关[13]。一般认为,iNOS激活及NO生成是由于NMDA受体激活及随后钙离子内流所致。本研究中,在甲醛注射前后鞘内注射CH(一种PKC抑制剂),可抑制PKC激活,这一药物并未改变NMDA受体的功能,但却发现iNOS活性受到抑制。这表明在甲醛炎性痛时,脊髓后角PKC的激活是iNOS激活的另一原因,即NOS活性不仅受到NMDA受体的影响,也受到其它因素的影响。如本实验通过观察PKC的活性,从而进一步证实了PKC在痛及痛觉过敏产生中的具体作用。其可通过影响NO生成而改变神经元的兴奋性。
为进一步确定PKC对iNOS的影响,本研究观察PKC兴奋剂对iNOS的影响,结果进一步支持上述观察。PMA,一种PKC兴奋剂,有类似二脂酰甘油(diacylglycerol,DAG)样作用,可持续激活PKC。在甲醛12 h组,此时单纯炎症并未使PKC充分激活,而甲醛注射前后鞘内注射PKC兴奋剂PMA可进一步增加PKC的活性,使相同时间点的iNOS活性增强,也进一步证实iNOS激活受PKC活性的调控。
众所周知,PKC和NO为细胞内两个不同信号转导系统的信号分子,以往观点认为,它们为两个不同信号转导系统。但两个系统之间是否存在着相互影响尚缺乏充足的证据。本研究证明甲醛引起的炎性痛及痛觉过敏时,不仅NMDA受体的激活可以使PKC激活及NO生成增多[14],同时PKC的活化也对神经元内iNOS活性及NO生成起促进作用,即NO生成在受NMDA受体激活影响的同时也受PKC调控。
甲醛炎性大鼠鞘内注射PKC兴奋剂PMA与抑制剂CH可分别明显地促进脊髓后角NOS或明显抑制NOS的生成,大鼠表现出相应的行为学变化。这表明大鼠脊髓后角中PKC是NOS生成的一条重要调节途径。
1 Terence J,Coderre M.Contribution of protein kinase C to central sensitization and persistent pain following tissue injury.Neuroscience Letters,1992,140:181-184.
2 Miletic V,Bowen K,Miletic G.Loosligation of the rat scitatic nerve is a accompanied by changes in the subcellular content o f protein kinase C beta П and gamma in the spinal dorsal horn.Neurosi Lett,2000,288:199-202.
3 Li KC,Zheng JH,Chen J.Invoivement of spinal protein Kinase C in induction and maintenance of both persistent Spontaneous flinching reflex and contralateral heat hyperalgesia induced by subcutaneous bee venom in the Concious rat.Neurosci Lett,2000,285:103-106.
4 Sachs D,Villarreal C,Cunha F,et al.The role of PKA and PK Cepsilon pathways in prostaglandin E2-mediated hypernociception.Br J Pharmacol,2009,156:826-834.
5 Roh DH,Kim HW,Yoon SY,et al.Intrathecal administration of sigma-1 receptor agonists facilitates nociception:involvement of a protein kinase C-dependent pathway.Neurosci Res,2008,86:3644-3654.
6 Gabra BH,Bailey CP,Kelly E,et al.Pre-treatment with a PKC or PKA inhibitor prevents the development of morphine tolerance but not physical dependence in mice.Brain Res,2008,27:70-77.
7 Inoue T,Mashimo T,Shibata M,et al.Rapid developent of nitric oxide-induced hyperalgesia depends on an alternate to the cGMP-mediated pathway in the rat neuropathic pain model.Brain Res,1998,792:263-270.
8 Moure PK,Oluyomi AO,Babbedge KC.L-NG-Nitro arginine methyl ester exhibits antinociceptive activity in the mouse.Br J Pharmacol,1991,102:198-202.
9 Yamamuto T,Shimoyana N.Role of nitric oxide in the development of thermal hyperagesia induced by sciatic nerve constriction injury in the rat.Anesthesiology,1995,82:1266-1273.
10 Schlechtweg PM,Roder J,Fischer MJ,et al.Increase in NADPH-diaphorase-positive and neuronal NO synthase immunoreactive neurons in the rat spinal trigeminal nucleus following infusion of a NO donor-evidence for a feed-forward process in NO production involved in trigeminal nociception.Cephalalgia,2009,29:566-579.
11 Lee JS,Zhang Y,Ro JY.Involvement of neuronal,inducible and endothelial nitric oxide synthases in capsaicin-induced muscle hypersensitivity.Eur J Pain,2009,13:924-928.
12 曾静波,李文斌,陈晓玲,等.大鼠甲醛炎性痛及痛觉过敏时脊髓后角NOS的变化及实践过程.中国神经科学杂志,2000,16:65-68.
13 Jung HS,Jeon HB,Jeon IS,at al.Preventing extracellular diffusion of trigeminal nitric oxide enhances formalin-induced oro facial pain.Korean J Physiol Pharmacol,2009,13:379-383.
14 曾静波,李文斌,李清君,等.MK-801降低炎性痛大鼠脊髓NOS的表达.生理学报,2000,84:55-59.
猜你喜欢
骨科(2021年3期)2021-06-05
昆明医科大学学报(2020年12期)2021-01-26
野生动物学报(2020年1期)2020-02-21
川北医学院学报(2018年5期)2018-11-16
电脑知识与技术(2016年34期)2017-04-15
中华骨与关节外科杂志(2016年3期)2016-05-17
癌症进展(2016年11期)2016-03-20
中国组织化学与细胞化学杂志(2016年3期)2016-02-27
中外医疗(2015年18期)2016-01-04
中国当代医药(2015年31期)2015-03-01
