
三七中高产人参皂苷Rd真菌的鉴定*
2012-11-21 02:40王艳代永东葛锋虞泓赵方允
食品与发酵工业 2012年9期
王艳,代永东,葛锋,虞泓,赵方允
1(昆明理工大学生命科学与技术学院,云南昆明,650500)2(云南大学中草药生物资源研究所云百草实验室,云南昆明,650091)3(昆明市延安医院,云南昆明,650224)
三七Panax notoginseng(Burk.)F.H.Chen为五加科人参属植物,主产于云南,主要活性成分为三七总皂苷(Panax notoginseng saponins,PNS)[1]。目前为止,已从三七中分离出70余种人参皂苷,均属达玛烷型四环三萜皂苷[2]。其中人参皂苷Rd的含量较少,西洋参含 0.1%~0.2%,人参含 0.2%~0.3%,三七含0.5%~0.7%[3]。Rd对心脑血管、神经系统、免疫系统等作用独特。目前,人参皂苷Rd已完成Ⅱ期临床试验,正在开展Ⅲ期临床研究,有可能成为治疗脑中风的新型临床治疗药物,同时,在镇痛、神经保护作用方面,人参皂苷Rd有比其他单体皂苷更强的活性[4]。人参皂苷Rd在植物中含量少,化学结构复杂,化学合成至今尚未成功,目前仅能从原植物的根、茎、叶中提取,效率低,成本高,严重制约了人参皂苷Rd的开发利用[5]。本研究组从云南文山三七根茎上分离得到能够高产人参皂苷Rd的真菌,经过形态鉴定及ITS分析,鉴定为长枝木霉Trichoderma longibrachiatum Rifai,关于长枝木霉转化人参皂苷为本文首次报道。
1 材料与方法
1.1 材料
1.1.1 实验材料
菌株WYA2012和WYAC2012均从云南文山的三七根茎分离纯化获得,保存于云南大学生命科学学院云百草实验室。
1.1.2 试剂
三七皂苷R1、人参皂苷Re和人参皂苷Rb1,购于中国药品生物制品检定所;人参皂苷Rg1、人参皂苷Rg3、人参皂苷Rh1、人参皂苷Rc、人参皂苷Rd、原人参三醇PPT、人参皂苷F1、人参皂苷F2和人参皂苷C-K,购于南方标准物质网;乙腈、甲醇均为色谱纯;Taq酶(配套 25mmol/L MgCl2,10 ×Buffer)、dNTP Mixture(ATP、TTP、CTP、GTP)均为上海生工分装,其余试剂均为市售分析纯。
1.1.3 仪器
高效液相色谱仪(戴安ULTIMATE 3000 LPG-3400A四元梯度泵,WPS-3000SL自动进样器,PDA-3000二极管阵列检测器,TCC-3000柱温箱,美国戴安仪器公司),Waters Symmetry C18色谱柱(250×4.6 μm,5 μm,100A),美国 Waters公司;Kodak Image Station 440CF凝胶成像系统,日本Kodak公司;PE9700扩增仪,美国PE公司;Dyy-Ⅲ-33A型垂直电泳仪,北京六一仪器厂。
1.1.4 培养基
(1)分离、纯化、活化培养基(PPDA):洋芋粉20 g/L,蔗糖2 g/L,琼脂18 g/L,蛋白胨5~20 g/L,自然pH,121℃灭菌 30 min。
(2)三七培养基:精确称取1 g过60目筛的三七粉于100 mL三角瓶,加入0.2 mL无机盐(10%KH2PO4,5%MgSO4),20 mL 水,自然pH,121℃灭菌30 min。
1.2 方法
1.2.1 菌株的筛选
将三七根茎粉碎加入到装有无菌水和玻璃珠的三角瓶中,充分摇匀,在不同稀释倍数,采用稀释涂布平板法和平板划线分离法,涂布到含有抗生素(链霉素30 μg/mL)的PPDA平板上,于23℃恒温培养,至长出菌落。挑取不同形态的菌丝分别接种于PPDA平板上纯化,挑取单菌落转接于斜面PPDA备用。
将菌种接种于三七培养基,发酵4d后,抽滤,用10 mL纯水清洗滤渣,合并滤液,离心(4 000 r/min)30 min,取上清液加入等体积水饱和正丁醇超声萃取1 h,静置分层后,收集上层正丁醇相旋蒸(60℃)至干燥,用5 mL 70%甲醇溶解残渣,进行HPLC检测。
1.2.2 菌种的形态鉴定
1.2.2.1 菌落形态观察
将供试菌株接种于PPDA和查氏培养基上,23℃培养,每隔1 d观察1次,记录菌落形态特征和培养性状。
湿室载片培养:将培养基琼脂薄层置于载玻片上,接种后盖上载玻片,置于用无菌水保湿的平皿内培养一定时间,以便菌体在载玻片和盖玻片之间沿载玻片横向生长。
1.2.2.2 产孢结构观察
观察湿室载片上自然生长的菌体状态。
1.2.3 菌株的ITS序列分析
(1)基因组DNA的提取:菌株基因组DNA的提取采用 CTAB 法[6]。
(2)ITS序列PCR扩增及测序:ITS序列引物[7]为 ITS4:5’-GGAAGTAAAAGT-CGTAACAAGG-3’,ITS5:5’-TCCTCCGCTTATTGATATGC-3’。扩增的反应体系(50 μL):灭菌去离子水 33 μL、10 × Buffer(含 2 mmol/L Mg2+)5 μL、2.5 mmol/L dNTP 4 μL、二甲基亚砜(dimethyl sulfoxid,DMSO)2.5 μL、10 μmol/L ITS4 和 ITS5 各2 μL、5U Taq 酶 0.5 μL、模板1 μL。扩增程序为:95℃ 4 min;94℃ 45 s,53℃ 45 s,72 ℃ 50 s,循环30 次,72 ℃ 8 min,12 ℃保存。琼脂糖电泳检测,选取条带清晰并无杂带的扩增样品送华大基因直接测序。
(3)ITS测序数据处理。序列拼接及BLAST搜索:根据测序数据,利用Bioedit检测ABI文件中峰行图的优劣,用Lasergene7.1中的SeqMan进行序列拼接,得到完整的 ITS序列。将上述序列在线 Blast(URL:http://blast.ncbi.nlm.nih.gov/Blast.cgi),发现目标序列与 T.longibrachiatum、T.konilangbra Samuels、Pardo-Schulth/Hypocrea stromatica Bezerra,Costa、Bastos等物种(属于Trichoderma Sect.Longibrachiatum)相似性较高。选择并下载若干T.Sect.Longibrachiatum物种ITS序列,并包含必要的正模标本序列数据,使用Clustalx 2.0[8]将以上ITS序列进行多序列比对,然后用MEGA 5.05中Analysis对ITS进行分区,统计ITS1、ITS2和5.8S区的长度和碱基总数和变异情况。利用ModleTest 3.7[9]寻找最适碱基替换模型,并选择最大似然法Maximum Likelihood Tree(ML)[10],Bootstrap 设为 1 000,以 T.stromaticum Samuels、Pardo-Schulth/Hypocrea stromatica Bezerra,Costa、Bastos为外类群构建系统聚类树。
2 结果与分析
2.1 活性菌株的筛选
通过对三七块茎内生真菌的分离与纯化,共得到16株真菌,分别发酵三七培养基后,经HPLC检测,得到2株能够高产人参皂苷Rd的菌株,分别编号为WYA2012和WYC2012,其中WYA2012转化三七的HPLC色谱图见图1。

图1 标准品(A),原药材(B),WYC2012转化三七(C)的HPLC色谱图
菌株WYA2012和WYC2012的HPLC检测结果如表1,经过真菌转化可以明显看出Rd由原药材的5.33 mg/g,分别增加至 14.17、15.40 mg/g,增加了 2倍,与此同时,小极性皂苷 Rh1、F1、F2、Rg3、PPT 和 CK都各有增加;相反,三七中主要的皂苷R1、Rg1、Re、Rb1显著下降,Rb1由原药材的18.77 mg/g分别降至0.42和0.45 mg/g,说明实验所用真菌能够转化三七中量多的大极性皂苷为小极性皂苷,且高产Rd。
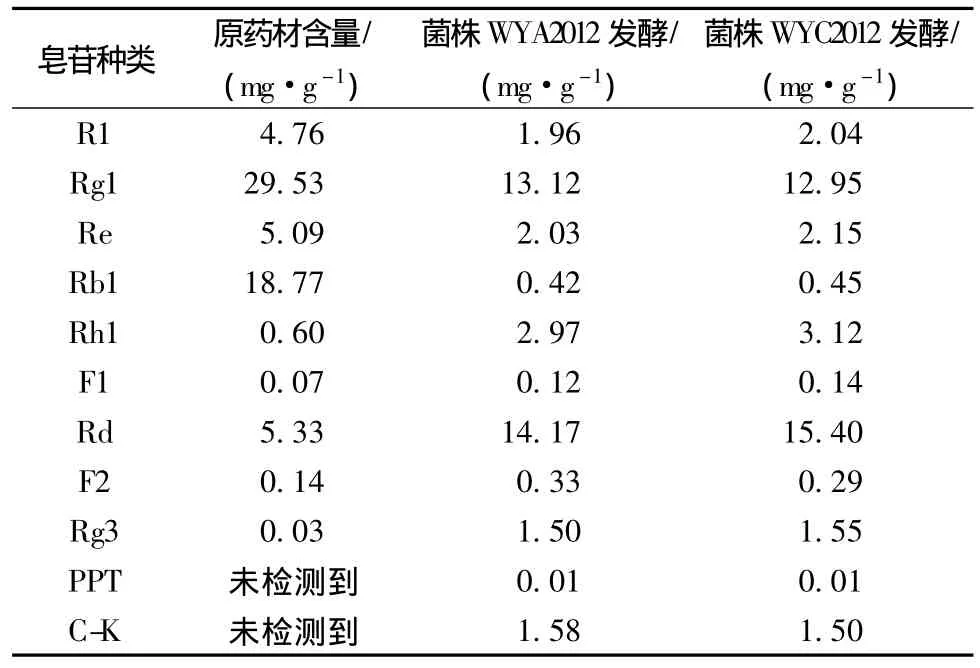
表1 株菌发酵三七的皂苷含量变化
2.2 菌株形态及显微结构鉴定
2.2.1 平板观察
菌株WYA2012形态特征见图2。在PPDA平板上培养1d,菌落生长缓慢,菌落短绒状,中部致密、隆起、呈“脓泡”状结构、雪白;3d后菌落迅速生长,且不规则,绒状隆起的产孢簇平展或疏松成簇,产生青绿色的分生孢子堆,孢子堆密集,呈绒状小团,多集中于菌落中间,菌落边缘出现散生的白色菌丝,菌落在培养基背面呈黄色。查氏平板上的培养性状:性状与PPDA的基本一致,不同的是培养基背面不变色(未产生色素),菌丝生长量明显较PPDA少很多。菌株WYC2012形态特征与菌株WYA2012大体一致,但菌落上产生的分生孢子堆是墨绿色,且孢子发散(见图2)。
2.2.2 显微观察
显微镜下可见2株菌的初级分生孢子梗长而直,以众多小丛束分布于菌落表面,次级分支较短,呈直角伸出或朝主枝弯曲,少见再分枝。瓶梗不规则排列,多数单生,偶尔2或3个轮状排列,基部略变细,圆柱状,顶部明显变细,瓶梗长(4.8)5.6~10.8 μ m,宽(1.9)2.5~4.5 μ m;分生孢子光滑,倒卵形或椭圆形,大小为(2.8)3.0~4.5 μ m×(1.4)1.8~2.6 μm,浅绿色。湿室载片观察见图3。
根据文成敬[11]、章初龙[12]、王斌[13]等人对木霉的分类研究,初步鉴定这2株真菌为木霉Trichoderma。

图2 菌株WYA2012与WYC2012平板形态特征

图3 株菌产孢结构及分生孢子(×100)
2.3 ITS序列分析及聚类树构建
在所研究的22条序列中,有2条序列为T.stromaticum,其余的均属于 T.Sect.Longibrachiatum,经过多序列比对,碱基统计,实验所选取的22条序列的ITS序列总长度为546~551bp,分离得到的2菌株ITS序列总长度为550bp,碱基总含量在57.80%~58.26%,总变异位点数为23个,其中ITS1区总长度217~224bp,GC含量为57.73%~59.36%,其中转换颠换位点6个,插入缺失位点8个;5.8S长度为160bp,GC含量为46.88%,无一变异位点;ITS2长度165~172bp,GC含量为68.02%~69.09%,其中转换颠换位点 2个,插入缺失位点 7个。发现 JC(Jukes-Cantor)+G(Gamma distribution)为最适模型,其BIC(Bayesian Information Criterion)分值最低。利用MEGA5.05软件使用JC+G模型构建ML聚类树,结果见图4。
从图4可以看出,2株木霉菌株与T.longibrachiatum聚为紧密一支,而其他种与其显著分开。另外,从ITS序列本身来看,发现实验的2株材料与本研究所选的来自GenBank中4株长枝木霉仅在全ITS中有一个位点的变异,即第134位点上发生缺失(或插入)变异。从而显著支持实验材料即为长枝木霉。
综合形态和显微结构及ITS序列分析,实验所用高产Rd的菌株为长枝木霉T.longibrachiatum。

图4 木霉菌株(WYC2012、WYA2012)及相关木霉菌物种菌株的ML聚类树
3 讨论
近几年,国内外用于转化Rd的菌株有灰绿犁头霉 Absidia glauca Hagem,专一转化 Rb1 为 Rd[14];镰刀菌Fusarium sacchari(E.J.Butler&Hafiz Khan)W.Gams可将Rb1转化为Rd,再转化为F2,进一步转化为C-K,也可将Rc转化为Fe或Mc,进一步转化为 C-K[15];黄枝孢霉 Cladosporium fulrum Cooke 可将Rb1 专一转化为 Rd[16];腐霉菌 Pythium irregular Buisman可将Rb1转化为Rd或绞股蓝皂苷XVII,进一步转化为 F2[17];拟青霉 Paecilomyces bainier sp.229可将Rb1大量转化为Rd,接着转化为F2或Rg3,F2进一步转化为C-K,而Rg3转化为 Rh2[18]。本研究菌株为长枝木霉T.longibrachiatum,对人参皂苷具有转化能力。
目前,木霉菌是生防益菌中的明星菌。早期对木霉菌的研究主要集中在植物病原菌的防治方面[19],近期的研究重点则是从中发现具有更广泛活性的化合物[20],诸如聚酮类(Polyketides)、氨基酸及其衍生物(amino acid and derivatives)、萜烯类(terpenes)等。此外,还有研究表明,木霉菌对植物有促生作用[21]。长枝木霉作为三七根茎上的真菌,可能与三七存在互利共生的关系,或具有拮抗多种病原真菌的作用,是具有潜力的资源微生物。根据国内外研究,高产Rd的转化机理可能与产生的葡聚糖酶或纤维素酶相关,水解人参皂苷糖基得到Rd[22]。本研究是以三七原药材作为转化底物,经长枝木霉发酵转化后,人参皂苷Rd含量明显增加,为有效利用开发三七生药功效成分提供了新思路。但因为转化机理复杂,所以不能明确转化途径,有待于深入研究。
一直以来,木霉菌因其形态特征易受培养条件及生长环境的影响,具有明显的生态多样性和物种多样性;加之鉴定过程中稳定形态性状少,变异性大,仅靠形态分类,人为误差较大,准确度较差。ITS序列逐渐被应用于木霉菌的鉴定中[23-24]。木霉菌属被分为4个组,分别是Sect.Longibrachiatum、Sect.Trichoderma、Sect.Pachybasium 和 Sect.Hypocreanum,这 4 个组在ITS聚类树中各自聚类在不同分支上。本研究选择Sect.Longibrachiatum物种为标靶,以T.stromaticum为外类群(该种与Sect.Pachybasium的 T.inhamatum Veerkamp and Gams为姐妹支[25-26])。聚类分析显示,本研究的2株菌株与长枝木霉T.longibrachiatum 菌 株(DAOM167674、DAOM231259、ATCC18648T、ATCC208859)在 Longibrachiatum 组内明显聚为一支,表现出极强的组内同质性。
根据上述2株菌株形态、菌落特征和产孢结构及其ITS分析,鉴定这2株菌株均为长枝木霉T.longibrachiatum。
[1]冯陆冰,潘西芬,孙泽玲.三七的药理作用研究进展[J].中国药师,2008,11(10):584- 589.
[2]Qi L W,Wang C Z,Yuan C S.Isolation and analysis of ginseng:advances and challenges[J].Natural Product Reports,2011(3):467 -495.
[3]Shi S,Lian-Wen Q,Guang-Jian D,et al.Red notoginseng:higher ginsenoside content and stronger anticancer potential than Asian and American ginseng[J].Food Chemistry,2011,125(4):1 299 -1 305.
[4]周超群,周珮.人参皂苷Rd的研究进展[J].中草药,2009,5(40):832-836.
[5]张薇,孙晓东,张萍,等.专一转化人参二醇类皂苷Rb1为Rd的真菌菌株的筛选[J].菌物学报,2011,30(2):305-311.
[6]Rogers S O,Bendich A J.Extraction of DNA from plant tissues.Plant Molecular Biology Manual[M].Dordrecht:Kluwer Academic Publishers,1988,A6:1-10.
[7]White T J,Bruns T,Lee S,et al.Applification and direct sequencing of fungal ribosomal RNA genes for phylogenetics.In:Innis M A,Gelfand D,Sninsky J J,White T J(Eds).PCR Protocols:A Guide to Methods and Applification[M].New York:Academic Press,1990:315-322
[8]Larkin M A,Blackshields G,Brown N P,et al.Clustal W and Clustal X version 2.0[J].Bioinformatics,2007,23(21):2 947-2 948.
[9]Posada D,Buckley T R.Model selection and model averaging in phylogenetics:advantages of the AIC and Bayesian approaches over likelihood ratio tests[J].Systematic Biology,2004,53(5):793-808.
[10]Tamura K,Peterson D,Peterson N,et al.MEGA5:Molecular evolutionary genetics analysis using Maximum Likelihood,evolutionary distance,and Maximum Parsimony methods [J].Molecular Biology and Evolution,2011,28(10):2731-2739.
[11]文成敬,陶家凤,陈文瑞.中国西南地区木霉属分类研究[J].真菌学报,1993,12(2):118-130.
[12]章初龙,徐同.我国河北、浙江、云南及西藏木霉种记述[J].菌物学报,2005,24(2):184-192.
[13]王斌,Ali Khatib Baker,刘金亮,等.长枝木霉TICC鉴定及其生物学特性研究[J].中国农学通报,2011,27(5):338-345.
[14]吕国忠,张薇,孙晓东.人参内生灰绿犁头霉菌及利用其制备人参皂苷 Rd的方法[P].CN,2101010571874,2011-06-01.
[15]Han Y,Sun B,Jiang B,et al.Microbial transformation of ginsenosides Rb1,Rb3 and Rc by Fusarium sacchari[J].Journal of Applied Microbiology.2010,109(3):792-798.
[16]Gao J,Zhao XS,Liu HB,et al.A highly selective ginsenoside Rb1-hydrolyzing β-D-glucosidase from Cladosporium fulvum[J].Process Biochemistry,2010,45:897 -903.
[17]Neculai M A,Ivanov D,Bernads M A.Partial purification and characterization of three ginsenoside-metabolizing β-glucosidases from Pythium irregular[J].Phytochemitry,2009,70(17/18):1 948-1 957.
[18]Yan Q,Zhou W,Shi XL,et al.Biotransformation pathways of ginsenoside Rb1 to compound K by β-glucosidases in fungus Paecilomyces bainier sp.229[J].Process Biochemistry,2010,45(9):1 550-1 556.
[19]Howell C R.Mechanisms employed by Trichoderma species in the biological control of plant diseases:the history and evolution of current concepts[J].Plant Disease,2003,87(1):4-10
[20]李纪顺,陈凯,杨合同,等.木霉抗生性代谢产物研究进展[J].农药,2010,49(10):713-719.
[21]赵蕾,滕安娜.木霉对植物的促生及诱导抗性研究进展[J].植物保护,2010,36(3):43-46.
[22]陈涛,徐燕.纤维素酶产酶菌株的选育与固态发酵[J].食品研究与开发,2011,32(11):119-125.
[23]Yu ZF,Qiao M,Zhang Y,et al.Two new species of Trichoderma from Yunnan,China[J].Antonie van Leeuwenhoek,2007,92(1):101-108.
[24]Hoyos-Carvajal L,Orduz S,Bissett J.Genetic and metabolic biodiversity of Trichoderma from Colombia and adjacent neotropic regions[J].Fungal Genetics and Biology,2009,46(9):615-631.
[25]Kuthls K,Samuels GJ,Meyer W,et al.Revision of Trichoderma sect.Longibrachiatum including related teleomorphs based on analysis of ribosomal DNA internal transcribed spacer sequences[J].Mycologia,1997,89(3):442-460
[26]Lee CF,Hseu TH.Genetic relatedness of Trichoderma sect.Pachybasium species based on molecular approaches[J].Canadian Journal of Microbiology,2002,48(9):831-840.
猜你喜欢
当代水产(2022年1期)2022-04-26
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
食品与机械(2019年1期)2019-03-30
中成药(2018年9期)2018-10-09
中成药(2017年9期)2017-12-19
中成药(2017年6期)2017-06-13
少儿科学周刊·少年版(2015年3期)2015-07-07
少儿科学周刊·少年版(2015年3期)2015-07-07
中国药业(2014年16期)2014-05-14
中国酿造(2014年9期)2014-03-11
