
人类面孔识别工作记忆的脑电位特征*
2013-01-31 01:46孙天义许远理郭春彦
心理学报 2013年10期
孙天义 许远理 郭春彦
(1华东师范大学心理与认知科学学院,上海 200062)(2信阳师范学院心理系,信阳 464000)(3首都师范大学心理系,北京市“学习与认知”重点实验室,北京 100048)
1 引言
面孔是一种意义丰富的非语言刺激,是人类表达、认知情感的重要工具和途径。面孔识别是人类社会生活的一项重要技能,在缺乏外部线索的情况下,它有助于我们对他人面孔的熟悉度、情绪状态、社会地位、性别、年龄和种族等方面有一个较明确的认识,从而更有利于人们社会交往和适应环境。
面孔识别与物体识别都是人类社会生活中的重要功能,两者之间是否有着相同的脑机制,是否存在独立的面孔加工系统与一般物体加工系统,这些是长期以来存在争议的问题(Moscovitch,Winocur,&Behrmann,1997)。有研究者主张面孔识别具有特异性,例如,面孔失认症病人虽然对面孔不能识别,但对其他物体的识别却基本没有困难(Diamond,Valentine,Mayes,&Sandel,1994;Meadows,1974;Young &Ellis,1989)。现代无创伤脑成像技术PET和fMRI方面的研究发现,人类大脑皮层上存在某些区域专门负责面孔加工,研究者将该区域称为梭状回面孔区(Kanwisher,McDermott &Chun,1997;McCarthy,Puce,Gore,&Allison,1997)。而高时间分辨率的ERP技术对跟踪面孔识别加工进程更为有效。如 Bentin,Allison,Puce,Perez和McCarthy(1996)在对正立面孔、倒置面孔、扭曲面孔、孤立面孔,以及非面孔刺激引发的 ERPs进行比较的实验中发现,只有刺激是面孔时,才会在腹侧颞叶后部和脑后部电极T5及T6处引发一个负波N170(时间在170 ms左右的负波),而汽车、手、家具等均不能产生 N170。还有研究发现,在枕颞两侧,面孔刺激诱发的负波波幅总是比以同等方式呈现的其他刺激(鸟、汽车等) 引起的波幅大得多(Eimer,1998,2000)。研究者认为面孔和其他物体之间的这种N170差异反映了面孔加工具有特异性。并且,研究还发现面孔 N170具有右半球优势(Rossion &Jacques,2008;Tanaka &Pierce,2009;Cauquil,Edmonds &Taylor,2000)。这些研究发现的“右半球优势”缺乏统一性,且所选择的电极较少(如Rossion和Jacques选择的是P10一个电极;James和Lara使用PO7、PO8;Cauquil,Edmonds和Taylor选用左侧枕颞的P7和P9,以及右侧枕颞区的 P8和 P10),缺乏代表性,不足以说明右半球优势的具体情况。此外,研究者发现人类面孔识别不仅具有特异性,而且具有较高的遗传性(Wilmer et al.,2010)。还有,对于面孔识别是整体加工还是局部加工仍存在争议。S.I.Choi,C.H.Choi,Jeong和 Kwak(2012)使用一种像素选择的方法,发现该方法可以提升面孔识别的成绩,这种方法还可以减少面孔识别加工的记忆和计算负荷,认为面孔识别为局部加工过程,而Van Belle,De Graef,Verfaillie,Busigny和Rossion(2010)的专家面孔识别研究却发现,专家的面孔识别不是依靠局部特征,而是视面孔为一整体,支持整体加工。
在人群中搜索特定的面孔(即靶),并把它与其他面孔(分心物)区分开来,涉及到工作记忆(working memory,WM) 。WM是指个体在执行认知任务过程中,用于信息的暂时储存与加工的资源有限的系统(Baddeley,2001,2003,2008,2010;Guo,Adam,Zhang,&Jiang,2008)。由于WM在人类高级认知活动中的突出重要性,Goldman-Rakic(1992)把它评价为“也许是人类心理进化中最重要的成就”。工作记忆作为一种理论构想虽然已有 30多年的研究历史,但对工作记忆如何测量还有待进一步的探索。郭春彦等人(Guo et al.,2008;刘荣,郭春彦,姜扬,2006)采用延迟样本匹配(Delayed matching-to-sample)任务范式,通过 ERP技术探索儿童及成人对客体工作记忆的神经机制,提出工作记忆的ERP测量方法,即使用靶刺激的ERPs减去分心物刺激的 ERPs,两者的差异波反映了被试对靶的保持和对分心物排除的过程。那么,作为视觉系统刺激加工的一个典型代表——面孔识别,其工作记忆用此方法测量结果怎样?两者之间有何异同?
已有的面孔识别研究,实验任务以简单的分类任务为主(如面孔与房子、汽车等),没有涉及到信息的保持与进一步加工,若以此揭示面孔识别加工的过程,证据显得还不够充足。此外,工作记忆过程会受长时记忆的影响,先前的学习应予以体现,并与未学习的内容进行对照。因此,我们采用延迟样本匹配任务范式(Guo et al.,2008),以面孔作为刺激物,分别考察对熟悉靶面孔、新靶面孔的追踪匹配,以及长时记忆对工作记忆的影响,目的在于:考察面孔识别脑电N170成分在熟悉靶和新靶、靶与分心物、以及左右枕颞区的异同,分析面孔识别 N170成分对应的加工阶段及其脑区优势效应;探索工作记忆中面孔识别的 ERP特征,验证工作记忆的ERP测量方法。
2 方法
2.1 被试
被试为16名在校本科生。男女各8名,年龄在19~23岁之间,平均年龄为 20.31岁。被试均为右利手,身体健康,视力或者矫正视力正常。实验前均与学生签订自愿实验协议,实验结束后给予一定报酬。
2.2 刺激材料
数码相机拍摄的青年人面孔彩色照片作为刺激材料,其中男女各半,表情中性,面部无明显标志(如胡须、眼镜、皮肤痣、化妆等),经软件Adobe Photoshop(7.0)处理后达到大小、明暗和对比度等一致。面孔图片共240张,图片大小为8.3 cm × 5.4 cm(高×宽)。实验前先让被试学习记忆其中的 60张面孔(男女各30张),在工作记忆任务阶段,该60张面孔图片分别作为熟悉靶或熟悉分心物;另 180张未见过的面孔图片中,60张用作新靶图片,余下的120张用作新分心物图片。整个刺激图片呈现在屏幕中央。
2.3 实验程序
实验共有12个组(Block),每组包括10个序列(sequence)。任一个序列都是由 1个靶图片和 9个测验图片组成,序列分两种:一种是“熟悉靶——熟悉靶、熟悉分心物、新分心物”;另一种为“新靶——新靶、熟悉分心物、新分心物”。且每个序列中面孔在性别上是一致的,总的序列中一半是男性面孔,一半是女性面孔。5个男性面孔序列和 5个女性面孔序列随机混合构成一个组,且每一组内的序列是同种序列(即同为“熟悉靶——熟悉靶、熟悉分心物、新分心物”或“新靶——新靶、熟悉分心物、新分心物”)。各组出现顺序采用“ABBAAB···”方法平衡。
实验分为学习阶段和测验阶段(实验程序见图1)。学习阶段,被试在正式实验之前先学习并记忆60张面孔图片,对这些面孔的再认正确率均在95%以上(为了保证高的正确率,让被试在正式实验前记忆这些面孔,达到较高的熟悉度);测验阶段,被试坐在照明柔和适度的实验室被试间内计算机屏幕前,双眼与屏幕中心齐平,视距 80 cm,视角约为 7°,要求被试始终注视屏幕中央的注视点,游戏柄的1,5两键作为左右手食指反应键。实验程序由 presentation(15.0版)编写,使用 17英寸(1024×768)联想 LCD 显示器,刷新频率为 60 Hz,奔Ⅲ计算机。
实验时先呈现一个靶面孔(熟悉靶或新靶)3000 ms,同时伴随着提示音“叮”的声音,之后随机呈现三种测验面孔图片,每种测验面孔图片重复出现2~4次(共 9次),不同类型测验图片呈现次数在整体上进行匹配。每个测验面孔图片呈现时间为700 ms,两个图片呈现间隔1000~1200 ms(ISI)随机变化,此间屏幕中央显示“+”注视点。相继两个序列之间呈现“+”注视点2500 ms。在出现三种测验图片时,要求被试对“靶”做一个按键反应,对“分心物”(熟悉的和新的)做另一个按键反应。左右手按键顺序进行被试间平衡,要求被试又快又准的进行反应。每组结束后被试休息2~3 min。
2.4 ERPs记录
采用Neuroscan ESI-64记录和分析系统,按国际10~20系统扩展的64导电极帽记录EEG。鼻尖作为参考电极(采集数据时以鼻尖做参考,离线分析时再转换为双侧乳突参考),另有 4个电极记录眼电:左眼眶上、下侧2个电极记录垂直眼电,两眼外侧的2个电极记录水平眼电。接地点在Fpz和Fz连线的中点,A/D采样率为500 Hz,电极与头皮接触电阻均保持小于5 kΩ。
2.5 ERPs数据处理与统计分析
完成连续记录原始脑电,然后离线(off-line)叠加处理数据。合并CNT原始数据后,程序剔除反应错误的序列。分析时段取刺激呈现到900 ms,滤波带通为0.05~40Hz,以-100~0 ms作为基线进行矫正,以±75 μV标准排除伴有眨眼、眼动、肌电等伪迹的数据,最后做组平均。本实验只分析测验图片的脑电数据,对反应正确的EEG进行叠加。根据相关研究(Gosling &Eimer,2011;Rossion &Caharel,2011;Studer et al.,2010;Rossion &Jacques,2008;Frühholz,Jellinghaus,&Herrmann,2011;Tanaka &Pierce,2009;Dennis,Malone,&Chen,2009)以及对本实验 ERPs波形的直观观察,面孔识别的特异成分N170分析选取枕颞区P7、P8、PO7、PO8、O1、和O2共6个电极,以160~180 ms的平均振幅作为N170成分的振幅;工作记忆中面孔识别的ERP特征选用5个中线电极FPz、Fz、Cz、Pz和Oz进行分析,分析时段为 250~450 ms、450~650 ms。
数据统计采用被试内重复测量的方差分析。根据本实验变量情况,在以往研究的基础上进行计划比较分析的方法(Keppel,1991),方差分析的p
值采用Greenhouse-Geisser法校正,脑电地形图由64导数据得出。
图1 实验程序
3 结果
3.1 行为数据
在不同靶面孔条件下,3种测验面孔图片的反应时和正确率见表1。采用2×3两因素重复测量方差分析,两因素分别为靶条件(新靶、熟悉靶)、测验图片类型(新分心物、熟悉分心物和靶)。统计结果显示,在平均反应时上,靶条件与测验图片的交互作用不显著,靶条件的主效应、测验图片的主效应均不显著,反应时均没有显著差异。正确率上,靶条件与测验图片的交互作用不显著,靶的主效应不显著,测验图片类型的主效应显著(F
(2,30)=38.28,p
<0.01),熟悉靶条件下,靶与新分心物、熟悉分心物差异显著(t检验p
值均小于0.001);新靶条件下,靶与新分心物、熟悉分心物差异显著(t检验p
值均小于0.001)。两种靶条件下,靶比分心物的正确率更低;新分心物、熟悉分心物间没有显著差异,新靶和熟悉靶的差异不显著。3.2 脑电结果
3.2.1 枕颞区两侧面孔特异性成分N170
新靶条件下,靶、熟悉分心物、新分心物在枕颞区两侧电极 P7、P8、PO7、PO8、O1、O2上的ERP波形,以及熟悉靶条件下,靶、熟悉分心物、新分心物在枕颞区两侧电极P7、P8、PO7、PO8、O1、O2上的ERP波形如图2,对应的各电极N170成分的振幅如表2。在同一电极位置上,单因素重复测量方差分析表明,六种面孔刺激(即新靶条件下靶、熟悉分心物、新分心物面孔,以及熟悉靶条件下靶、熟悉分心物、新分心物面孔)诱发的N170成分均没有显著差异(ps
>0.9);在枕颞区,三因素重复测量方差分析2(左侧:P7、PO7、O1,右侧:P8、PO8、O2) × 3(前:P7、P8,中:PO7、PO8,后:O1、O2) × 6(新靶条件下的靶,熟悉分心物,新分心物,熟悉靶条件下的靶,熟悉分心物,新分心物)显示,三重交互作用不显著,左右侧与前中后位置两重交互作用显著(F
(2,30)=8.86,p
<0.01),进一步简单效应检验发现,在P7、P8两电极上N170振幅差异显著(p
<0.05),P8的振幅大于P7的振幅(如图3所示),即右侧颞叶振幅大于左侧颞叶振幅,但左右位置的主效应不显著,因此,面孔识别 N170的右半球优势具体为颞区的右侧优势。3.2.2 靶与分心物的ERPs
考察头皮电极上波形总的特点,选取有代表性的前额区(FPz)、额区(Fz)、中央区(Cz)、顶区(Pz)和枕区(Oz)作为分析的电极,在新靶条件和熟悉靶条件下,靶、熟悉分心物、新分心物的波形如图4,对应的面孔刺激均诱发了 P300成分,在顶区达到峰值。靶和分心物从250 ms开始分离,延迟到500 ms左右。靶比分心物的波形更正。
熟悉靶条件下,进行3(靶、熟悉分心物、新分心物) × 5(FPz、Fz、Cz、Pz、Oz)两因素重复测量方差分析,结果表明:在250~450 ms,交互作用显著(F
(8,120)=5.22,p
<0.01),进一步简单效应检验发现,靶与两种分心物在 FPz、Fz、Cz、Pz及 Oz差异显著(ps
<0.01),且靶的振幅显著高于两种分心物,熟悉分心物仅在FPz显著正于新分心物(p
<0.05),而其它脑区差异不显著;在450~650 ms,两因素的交互作用显著(F
(8,120)=19.29,p
<0.01),简单效应检验显示,靶的振幅与两种分心物的振幅在FPz、Fz上差异显著,靶的振幅低于分心物(ps
<0.05),在Cz上无显著差异,而Pz、Oz上靶显著高于分心物(ps
<0.05)。熟悉分心物振幅仅在FPz上显著高于新分心物的振幅(p
<0.05)。新靶条件下,3(靶、熟悉分心物、新分心物) × 5(FPz、Fz、Cz、Pz、Oz)两因素重复测量方差分析表明:在 250~450 ms,交互作用显著(F
(8,120)=3.26,p
<0.01),简单效应检验显示,靶与两种分心物在 FPz、Fz、Cz、Pz及 Oz差异显著(ps
<0.05),且靶的振幅显著高于两种分心物,熟悉分心物仅在FPz边缘显著正于新分心物(p
=0.06),其它脑区差异不显著;在450~650 ms,两因素的交互作用显著(F
(8,120)=13.98,p
<0.01),简单效应检验显示,靶的振幅与两种分心物的振幅在FPz、Fz上差异显著,靶的振幅低于分心物(ps
<0.05),在 Cz上无显著差异,而 Pz、Oz 上靶显著高于分心物(ps
<0.05)。熟悉分心物振幅与新分心物的振幅差异不显著。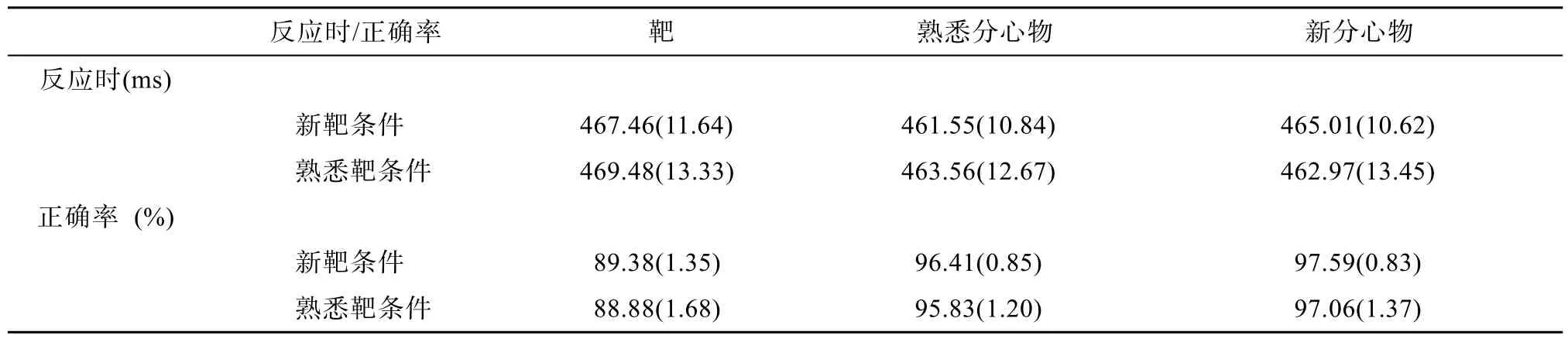
表1 平均反应时与正确率(园括号内数值为标准误)
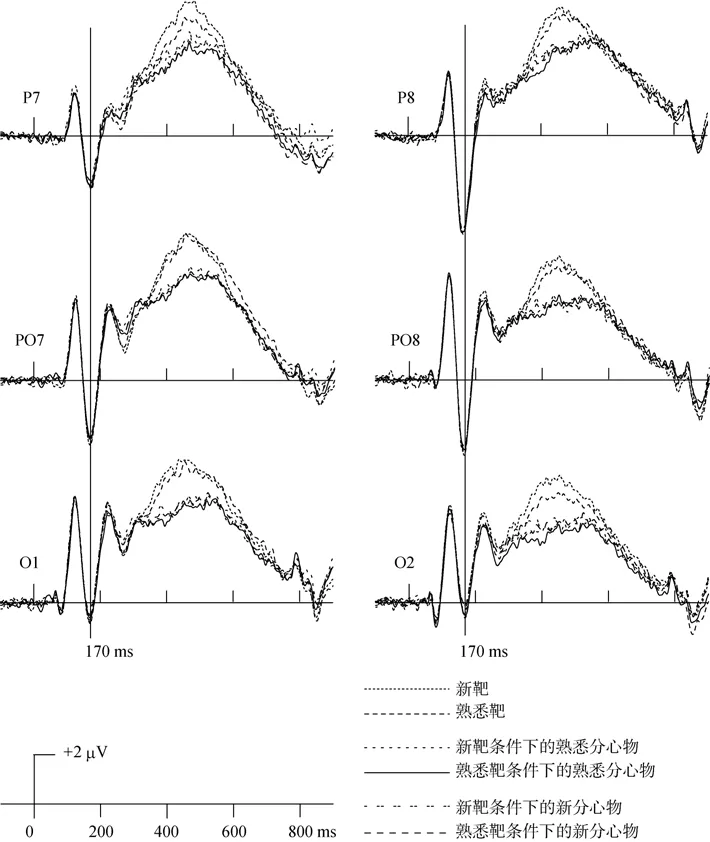
图2 电极P7、P8、PO7、PO8、O1、O2在不同靶条件下靶、熟悉分心物和新分心物的波形

表2 靶、熟悉分心物和新分心物的N170平均振幅(圆括号内数值为标准误,单位为μV)
(1) 新靶与熟悉靶的ERPs
考察不同靶诱发的 ERP波,发现新靶与熟悉靶在整个脑区均诱发了明显的 P300成分,新靶波形比熟悉靶更正(如图5 A)。
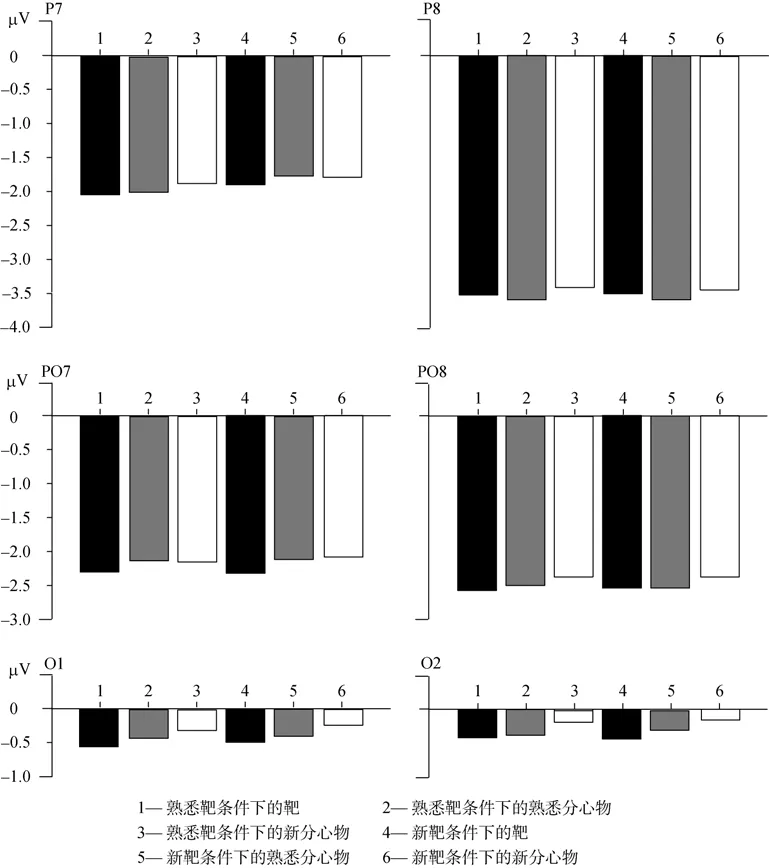
图3 靶、熟悉分心物和新分心物的N170平均电位
2(新靶、熟悉靶) × 5(FPz、Fz、Cz、Pz、Oz)两因素重复测量方差分析表明:在250~450 ms,靶的主效应显著(F
(1,15)=6.78,p
<0.05),新靶比熟悉靶更正,但两因素交互作用不显著;450~650 ms,靶的主效应不显著,两因素的交互作用也不显著。这与两种靶的差异波地形图(图5 B)相吻合。(2) 新分心物与熟悉分心物的ERPs
分别将两种靶条件下的新分心物、熟悉分心物进行合并,结果发现新分心物和熟悉分心物的波形相似,并且走向一致,两种分心物都诱发 P300成分并在脑区后部的振幅最大,熟悉分心物比新分心物更正(图6 A),并且主要出现在前额区。
2(新分心物、熟悉分心物) × 5(FPz、Fz、Cz、Pz、Oz)两因素重复测量方差分析发现:在250~450 ms,两因素的交互作用显著(F
(4,60)=5.57,p
<0.01),简单效应检验显示,分心物的振幅在FPz上差异显著,熟悉分心物的振幅高于新分心物;450~650 ms出现类似的结果,两因素的交互作用显著(F
(4,60)=3.56,p
=0.01),简单效应检验表明熟悉分心物的振幅在FPz上显著高于新分心物。这与两种分心物差异波的地形图(图6 B)相一致。(3) 新旧工作记忆的ERPs
工作记忆的测量采用靶ERPs减去分心物ERPs的方法,此种方法体现了被试对靶的保持、选择和对分心物排除的过程。在新靶条件下,用靶 ERPs减去新分心物 ERPs获得新面孔的工作记忆 ERPs(以下简称新工作记忆);熟悉靶条件下,靶ERPs减去熟悉分心物ERPs获得熟悉面孔的工作记忆ERPs(以下简称旧工作记忆)。从ERPs波形观察(图7 A),新、旧工作记忆均在300 ms至400 ms出现波峰,两者的潜伏期略有不同,新工作记忆比旧工作记忆更早。新、旧工作记忆的脑地形图(图7 B)与此吻合。
2(新工作记忆、旧工作记忆) × 5(FPz、Fz、Cz、Pz、Oz)两因素重复测量方差分析发现:在250~450 ms两因素交互作用未达到显著水平,工作记忆的主效应也均不显著;450~650 ms,两因素交互作用不显著,工作记忆的主效显著(F
(1,15)=4.54,p
<0.05),新工作记忆比旧工作记忆的波形更正。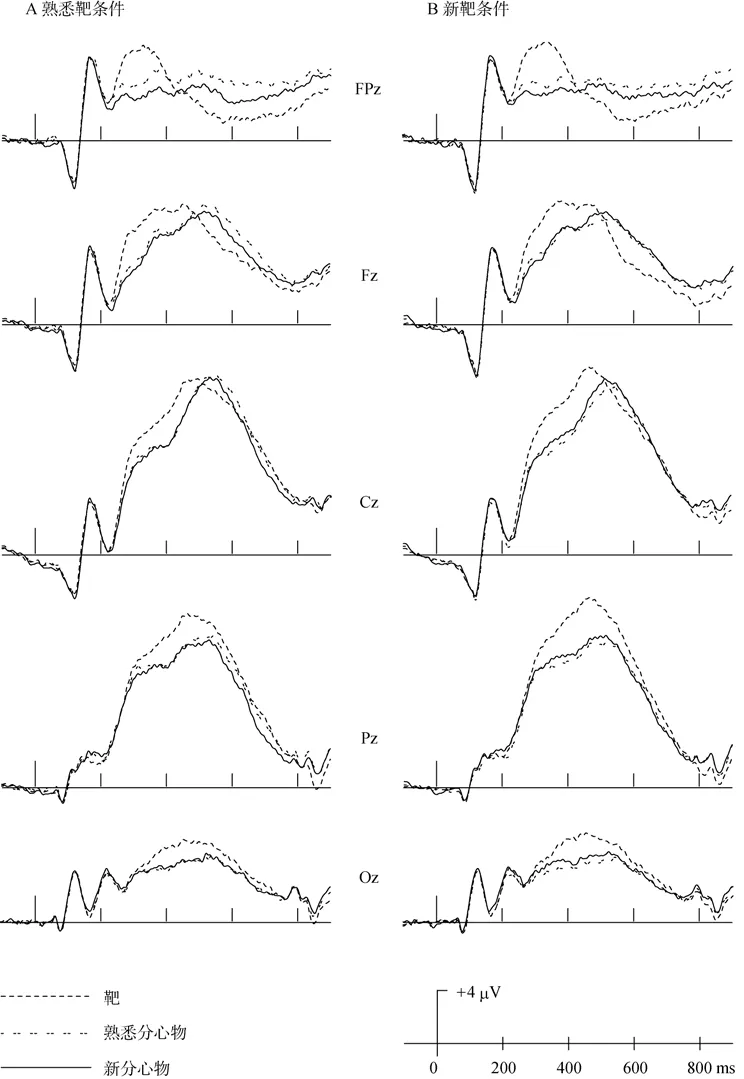
图4 靶、分心物ERPs波形
4 讨论
采用延迟样本匹配任务范式探讨面孔识别的工作记忆。实验时被试先看 1张靶面孔(熟悉面孔或新面孔),接着对呈现的 9张面孔(包括靶、熟悉分心物和新分心物)做靶和分心物的区分判断反应。Desimone(1996)曾提出类似的过程中,被试需要进行三种操作才能正确地完成匹配任务。首先,被试必须注意并知觉靶刺激。其次,在整个序列持续的时间里被试必须保持对靶的记忆。最后,被试必须对测试项刺激进行评估并判断是否与记忆里的靶匹配,这通常是以 P300脑电成分激活为指标的。因此,本实验过程体现了面孔识别的工作记忆。
行为数据结果显示,在正确率上,无论新靶还是熟悉靶都比其对应的分心物显著低。在反应时方面,虽然靶与分心物的平均反应时差异不显著,但出现对靶反应长于分心物的趋势(可能是实验被试只有 16人,数量少而未表现出显著差异)。这暗示着,正确识别一个与记忆中保持的靶相匹配的面孔比排除非匹配面孔需要更多的注意资源。

图5 靶的ERPs波形图和地形图
4.1 面孔识别加工的N170成分

图6 分心物的ERPs波形图和地形图
N170是一个峰值在170 ms左右呈两侧枕颞头皮分布的负性成分,Bentin等人(1996)最早报告了在枕颞部的一个潜伏期约为 170 ms 的负成分(N170)与面孔身份识别(结构分析)有关,N170与面孔熟悉度、性别、年龄和种族等信息加工无关。我们的实验结果表明,N170成分与面孔是追踪的熟悉靶还是新靶无关、与面孔是干扰的熟悉分心物还是新分心物分也无关,并且在枕颞区两侧较大的皮层,熟悉靶面孔、新靶面孔、熟悉分心物面孔及新分心物面孔均诱发一致的 N170波,暗示着在工作记忆任务过程中,面孔识别是早期阶段的整体加工,并且以右半球 N170成分为特征,这与 Ramo和Rossion(2012)的发现一致,进一步证实N170成分反映了Bruce-Young模型中早期的预分类结构编码过程(Bruce &Young,1986;Rossion &Jacques,2008)。尽管也有报告称成人面孔识别N170的潜伏期在150 ms左右(Rousselet,Husk,Bennett &Sekuler,2008;Caharel,d’Arripe,Ramon,Jacques,&Rossion,2009;Marini,Marzi,&Viggiano,2011;Rossion &Caharel,2011),这可能是记录脑电数据时所选取的参考不同造成的(Rousselet等人的研究以Cz电极做参考;Caharel等人、Rossion等人的研究以左侧乳突做参考;Marini等人的研究以右侧乳突做参考)。我们采用鼻尖做参考,得到成人面孔识别 N170成分的潜伏期为 170 ms左右,这与Frühholz,Jellinghaus 和 Herrmann(2011)的研究结果一致。在 N170的半球优势方面,我们的实验结果发现仅在颞区的P8比P7波峰显著高,其他左右两侧脑区(如PO7与PO8、O1与O2)波峰没有差异,数据统计分析结果表明左右侧脑区主效应不显著,因此,面孔识别 N170右半球优势具体为颞区的右侧优势。Sadeh,Podlipsky,Zhdanov和Yovel(2010)结合ERP和fMRI,发现面孔选择反应发生在颞叶和颞上回,与面孔选择 N170高相关,而不是在枕叶皮层。
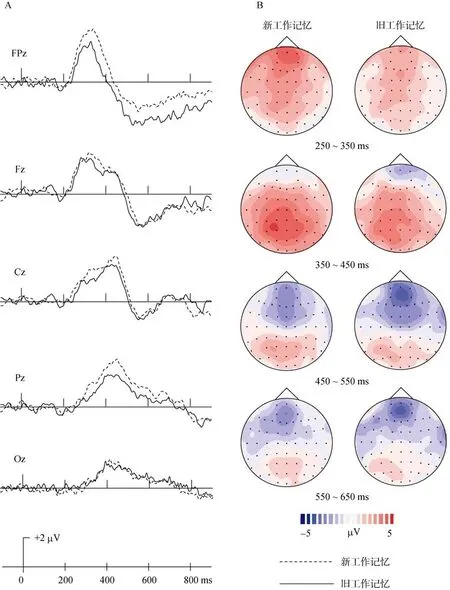
图7 工作记忆的ERPs波形图和地形图
对于脑电成分 N170的本质,它是否是面孔识别特异性的反映,Tanaka和 Curran(2001)曾提出N170 并非特异于面孔,而是特异于专家识别。但面孔识别比物体识别特殊,面孔识别不仅涉及到知觉加工,还涉及到社会性信息加工。Liu,Harris和Kanwisher(2002)发现面孔识别在面孔呈现后 100 ms内,被试已经将面孔刺激与非面孔刺激区分开了,同时亦发现多个脑区参与面孔识别(Liu,Harris,&Kanwisher,2010)。基于此,对面孔识别N170成分的特异性还有待进一步的研究(陶维东,孙宏进,张旭东,郑剑虹,2011)。
4.2 在追踪不同靶条件下面孔识别的工作记忆
面孔识别工作记忆的 ERPs呈现出 250ms ~650ms左右的一个正波成分——P300,我们的结果与许多工作记忆研究类似(Gevins et al.,1996;Kusak,Grune,Hagendorf,&Metz,2000;刘荣等,2006;Guo et al.,2008;Rossion &Jacques,2008;Dennis et al.,2009;Frühholz et al.,2011;Rossion &Caharel,2011)。工作记忆系统的加工过程与P300成分相关联,P300的振幅被认为是任务相关和注意的反映,任务相关和注意的对象 P300振幅更大,而被忽略或非注意项目的 P300振幅减小(Kok,2001;Pritchard,1981);P300的潜伏期反映了对刺激评估持续的时间,注意目标的潜伏期更短(Polich,2007;George &Coch,2011)。在我们面孔识别的工作记忆中,靶面孔是与任务相关联的,是注意的目标,靶的识别需要与记忆保持中的信息匹配,与忽略的分心物面孔相比,靶面孔需要更多的注意资源。因此,靶面孔诱发 P300振幅比分心物面孔更大,且潜伏期更短。在实验中,熟悉靶条件下的 250~450 ms时程,靶的振幅明显高于分心物,在450~650 ms时段,靶在顶区和枕区的振幅高于分心物,而在中央区与分心物的振幅没有差异,在前额区和额区靶的振幅却比分心物显著低;在新靶条件下,两个时段的靶与分心物振幅之间的关系也是如此。这正是由于靶诱发的P300波比分心物更正,且靶的P300波潜伏期短于分心物引起的,体现了工作记忆中对靶的保持需要更多的注意资源。与以往的研究结论(Polich,2004)一致,本实验中靶与分心物的 P300振幅均在顶区最大,且靶振幅更大。Courchesne(1978)采用改编后的 Oddball范式,呈现两个概率不同的刺激,发现对容易再认的小概率刺激(注意不是新异刺激)诱发的 P300成分在中央区/顶区振幅最大。该实验中的小概率刺激类似于本研究中小概率“靶”刺激,因此本实验中“靶”诱发的P300在顶区振幅最大的结果与其结论一致。
新靶与熟悉靶在分析的脑区均诱发明显的P300波,差异波的地形图及数据统计结果都显示两者的差异表现在分析时段早期的 250~450 ms,新靶的振幅显著高于熟悉靶,而在后期没有差异。新靶与熟悉靶的这种差异可能是由于前额皮层接收来自腹侧颞叶和颞中皮层输入信息的不同造成的。在工作记忆中前额皮层对刺激的评估加工十分重要(Desimone,1996;Goldman-Rakic,Cools &Srivastava,1996;Levy &Goldman-Rakic,2000)。腹侧颞叶(例如梭状回面孔和物体区)负责知觉加工,而颞中皮层是再认记忆的重要脑区(Haxby et al.,2001;Kanwisher et al.,1997)。此外,匹配一个项目包括知觉加工和旧/新加工的脑结构(Guo et al.,2008)。相对于熟悉靶而言,暂时记住新靶并在相继任务中进行判断需要更多的心理资源。
新分心物与熟悉分心物在前额区出现“旧/新效应”(old/new effect),已有的研究(Cycowicz,Friedman,Snodgrass,&Rothstein,2000;Itier &Taylor,2004;Guerin &Miller,2009)发现,“旧/新效应”通常出现在额区和(或)顶区,时程为300~800 ms。反映了测试阶段重复出现的旧刺激所诱发的波形变化,旧刺激会诱发更正的波形。本实验分心物的“旧/新效应”虽在潜伏期上有所提前,但脑区分布与上述研究结果一致。此外,靶的类似“旧/新效应”与分心物的“旧/新效应”完全不同,新靶的波形更正,且仅在早期 250~450 ms时段出现在分析的所有脑区,分心物的“旧/新效应”始终体现在大脑的前额区。脑地形图也表明,靶与分心物的“旧/新效应”完全不同。这种差异的出现可能源于实验任务范式的差异,一般地,典型的“旧/新效应”产生于外显再认任务中,而本实验使用了工作记忆任务范式,因此靶的“旧/新效应”属于内隐性质。另外,典型的“旧/新效应”产生于同一个任务序列中,而“靶”旧/新效应则不同,新靶和熟悉靶分别呈现于两个实验序列中,追逐新靶可能需要更多的注意资源,因此表现出新靶波形比熟悉靶波形更正的结果。最后,我们认为个体在追逐目标并同时排除熟悉的与新的非目标时,记忆加工的两类 ERPs“旧/新效应”可能具有不同的神经机制。总之,对熟悉面孔和不熟悉面孔的加工是有显著差异的(Davies-Thompson,Newling,&Andrews,2013)。
在工作记忆的测量上,采用“靶”ERPs减去“分心物”ERPs的方法(Guo et al.,2008;刘荣等人,2006),该操作集中体现了工作记忆的过程:被试根据要求对靶和分心物进行识别时,在头脑中必须始终保持着“靶”刺激,且对靶刺激进行选择反应的同时,不断排除熟悉分心物和新分心物。本实验进一步验证了该评估方法的可行性。工作记忆的波形图和地形图(图7)显示新、旧工作记忆主要表现在300 ms至400 ms阶段,新工作记忆的潜伏期更短,并且在工作记忆的后期(450~650 ms)阶段,与旧工作记忆相比,新工作记忆的正波振幅更大,新客体诱发了更强的神经反应。这表明在大脑中,工作记忆加工受新异或熟悉环境的影响不同。Cycowicz和 Friedman(1998)认为此现象可能与注意资源分配有关,其对工作记忆 P300成分的研究表明,偶尔呈现的事件比经常呈现的事件会引发更大的振幅反应。
综上可知,先前的学习对面孔识别 N170成分未产生影响,但会影响工作记忆过程中大脑对追踪靶和拒绝分心物的反应。此外,面孔识别工作记忆的脑电特征与相同实验范式下的物体识别(Guo et al.,2008)相比,存在明显不同。物体识别工作记忆中靶与分心物脑波在 200 ms左右出现分离,时间比面孔识别早;新靶与熟悉靶之间的差异,物体识别工作记忆表现在额区和中央区,而面孔识别表现更广,还包括中央区、顶区及枕区;熟悉分心物与新分心物之间的“旧/新效应”,物体识别时出现在顶区和枕区,而面孔识别反映在额区;新旧工作记忆方面,物体识别在 200~400 ms表现出新工作记忆的波幅更正,而面孔识别是在 450~650 ms新工作记忆的波幅更正。这些差异都暗示着面孔识别和物体识别可能是相互独立的两种加工过程,两者在脑机制上可能不同。
5 结论
我们的事件相关电位实验研究表明,面孔识别的 N170效应可能反映了知觉的整体加工阶段,且N170右半球优势具体为颞区的右侧优势;先前的面孔学习形成的长时记忆会影响工作记忆期间大脑对面孔的识别反应。
Baddeley,A.D.(2001).Is working memory still working?American Psychologist,56
,851−864.Baddeley,A.D.(2003).Working memory:Looking back and looking forward.Nature Reviews Neuroscience,4
,829−839.Baddeley,A.D.(2008).What′s new in working memory?Psychological Review,13
,2−5.Baddeley,A.D.(2010).Working memory.Current Biology,20
,R136−R140.Bentin,S.,Allison,T.,Puce,A.,Perez,E.,&McCarthy,G.(1996).Electrophysiological studies of face perception in humans.Journal of Cognitive Neuroscience,8
,551−565.Bruce,V.,&Young,A.(1986).Understanding face recognition.British Journal of Psychology,77
,305−327.Caharel,S.,d’Arripe,O.,Ramon,M.,Jacques,C.,&Rossion,B.(2009).Early adaptation to repeated unfamiliar faces across viewpoint changes in the right hemisphere:Evidence from the N170 ERP component.Neuropsychologia,47
,639−643.Cauquil,A.S.,Edmonds,G.E.,&Taylor,M.J.(2000).Is the face-sensitive N170 the only ERP not affected by selective attention?NeuroReport,11
,2167−2171.Choi,S.I.,Choi,C.H.,Jeong,G.M.,&Kwak,N.(2012).Pixel selection based on discriminant features with application to face recognition.Pattern Recognition Letters,33
,1083−1092.Courchesne,E.(1978).Changes in P3 waves with event repetition:Long-term effects on scalp distribution and amplitude.Electroencephalography and Clinical Neurophysiology,45
,754−766.Cycowicz,Y.M.,&Friedman,D.(1998).Effect of sound familiarity on the event-related potentials elicited by novel environmental sounds.Brain and Cognition,36
,30−51.Cycowicz,Y.M.,Friedman,D.,Snodgrass,J.G.,&Rothstein,M.(2000).Adevelopmental trajectory in implicit memory is revealed by picture fragment completion.Memory,8
,19−35.Davies-Thompson,J.,Newling,K.,&Andrews,T.J.(2013).Image-invariant responses in face-selective regions do not explain the perceptual advantage for familiar face recognition.Cerebral Cortex,23
,370−377.Dennis,T.A.,Malone,M.M.,&Chen,C.C.(2009).Emotional face processing and emotion regulation in children:An ERP study.Developmental Neuropsychology,34
,85-102.Desimone,R.(1996).Neural mechanisms for visual memory and their role in attention.Proceedings of the National Academy of Sciences of the United States of America,93
,13494-13499.Diamond,B.J.,Valentine,T.,Mayes,A.R.,&Sandel,M.E.(1994).Evidence of covert recognition in a prosopagnosic patient.Cortex,30
,377−393.Eimer,M.(1998).Does the face-specific N 170 component reflect the activity of a specialized eye detector.NeuroReport,9
,2945−2948.Eimer,M.(2000).The face-specific N 170 component reflects late stages in the structural encoding of faces.NeuroReport,11
,2319−2324.Frühholz,S.,Jellinghaus,A.,&Herrmann,M.(2011).Time course of implicit processing and explicit processing of emotional faces and emotional words.Biological Psychology,87
,265−274.George,E.M.,&Coch,D.(2011).Music training and working memory:An ERP study.Neuropsychologia,49
,1083−1094.Gevins,A.,Smith,M.E.,Le,J.,Leong,H.,Bennett,J.,Martin,N.,et al.(1996).High resolution evoked potential imaging of the cortical dynamics of human working memory.Electroencephalography and Clinical Neurophysiology,98
,327−348.Goldman-Rakic,P.S.(1992).Working memory and the mind.Scientific American,267
,110−117.Goldman-Rakic,P.S.,Cools,A.R.,&Srivastava,K.(1996).The prefrontal landscape:Implications of functional architecture for understanding human mentation and the central executive [and discussion].Philosophical Transactions of the Royal Society Series B:Biological Sciences,351
,1445−1453.Gosling,A.,&Eimer,M.(2011).An event-related brain potential study of explicit face recognition.Neuropsychologia,49
,2736−2745.Guerin,S.A.,&Miller,M.B.(2009).Lateralization of the parietal old/new effect:An event-related fMRI study comparing recognition memory for words and faces.NeuroImage,44
,232−242.Guo,C.Y,Adam,L.L.,Zhang,Q.,&Jiang,Y.(2008).Brain potentials distinguish new and studied objects during working memory.Human Brain Mapping,29
,441−452.Haxby,J.V.,Gobbini,M.I.,Furey,M.L.Ishai,A.,Schouten,J.L.,&Pietrini,P.(2001).Distributed and overlapping representations of faces and objects in ventral temporal cortex.Science,293
,2425−2430.Itier,R.J.,&Taylor,M.J.(2004).Face recognition memory and configural processing:Adevelopmental ERP study using upright,inverted,and contrast-reversed faces.Journal of Cognitive Neuroscience,16
,487−502.Kanwisher,N.,McDermott,J.,&Chun,M.M.(1997).The fusiform face area:Amodule in human extrastriate cortex specialized for face perception.Journal of Neuroscience,17
,4302−4311.Keppel,G.(1991).Design and analysis:Aresearcher’s handbook
(3rd ed.).Englewood cliffs,NJ,US:Prentice Hall,Inc.Kok,A.(2001).On the utility of P3 amplitude as a measure of processing capacity.Psychophysiology,38
,557−577.Kusak,G.,Grune,K.,Hagendorf,H.,&Metz,A.M.(2000).Updating of working memory in a running memory task:An event-related potential study.International Journal of Psychophysiology,39
,51−65.Levy,R.,&Goldman-Rakic,P.S.(2000).Segregation of working memory functions within the dorsolateral prefrontal cortex.Experimental Brain Research,133
,23−32.Liu,J.,Harris,A.,&Kanwisher,N.(2002).Stages of processing in face perception:An MEG study.Nature Neuroscience,5
,910−916.Liu,J.,Harris,A.,&Kanwisher,N.(2010).Perception of face parts and face configurations:An fMRI study.Journal of Cognitive Neuroscience,22
,203−221.Liu,R.,Guo,C.Y.,&Jiang,Y.(2006).An event-related potential study of working memory in children.Chinese Science Bulletin,51
,1467−1475.[刘荣,郭春彦,姜扬.(2006).儿童工作记忆的事件相关电位.科学通报,51
,1182−1189.]Marini,F.,Marzi,T.,&Viggiano,M.P.(2011).“Wanted!”The effects of reward on face recognition:Electrophysiological correlates.Cognitive,Affective,&Behavioral Neuroscience,11
,627−643.McCarthy,G.,Puce,A.,Gore,J.C.,&Allison,T.(1997).Face-specific processing in the human fusiform gyrus.Journal of Cognitive Neuroscience,9
,605−610.Meadows,J.C.(1974).The anatomical basis of prosopagnosia.Journal of Neurology,Neurosurgeon &Psychiatry,37
,489−501.Moscovitch,M.,Winocur,G.,&Behrmann,M.(1997).What′s special about face recognition? Nineteen experiments on a person with visual object agnosia and dyslexia but normal face recognition.Journal of Cognitive Neuroscience,9
,555−604.Polich,J.(2004).Neuropsychology of P3a and P3b:Atheoretical of overview
.California,USA:Kjellberg Inc.Polich,J.(2007).Updating P300:An integrative theory of P3a and P3b.Clinical Neurophysiology,118
,2128−2148.Pritchard,W.S.(1981).Psychophysiology of P300.Psychological Bulletin,89
,506−540.Ramon,M.,&Rossion,B.(2012).Hemisphere-dependent holistic processing of familiar faces.Brain and Cognition,78
,7−13.Rossion,B.,&Caharel,S.(2011).ERP evidence for the speed of face categorization in the human brain:Disentangling the contribution of low-level visual cues from face perception.Vision Research,51
,1297 −1311.Rossion,B.,&Jacques,C.(2008).Does physical interstimulus variance account for early electrophysiological face sensitive responses in the human brain? Ten lessons on the N170.NeuroImage,39
,1959−1979.Rousselet,G.A.,Husk,J.S.,Bennett,P.J.,&Sekuler,A.B.(2008).Time course and robustness of ERP object and face differences.Journal of Vision,8
,1−18.Sadeh,B.,Podlipsky,I.,Zhdanov,A.,&Yovel,G.(2010).Event-related potential and functional MRI measures of face-selectivity are highly correlated:Asimultaneous ERP-fMRI investigation.Human Brain Mapping,31,
1490−1501.Studer,P.,Wangler,S.,Diruf,M.S.,Kratz,O.,Moll,G.H.,&Heinrich,H.(2010).ERP effects of methylphenidate and working memory load in healthy adults during a serial visual working memory task.Neuroscience Letters,482
,172−176.Tanaka,J.W.,&Curran,T.(2001).Aneural basis for expert object recognition.Psychological Science,12
(1),43−47.Tanaka,J.W.,&Pierce,L.J.(2009).The neural plasticity of other-race face recognition.Cognitive,Affective,&Behavioral Neuroscience,9
,122−131.Tao,W.D.,Sun,H.J.,Zhang,X.D.,&Zheng,J.H.(2011).The neural-mechanisms of the formation of inversion effect in non-face object recognition.Advances in Psychological Science,19
,1104−1114.[陶维东,孙宏进,张旭东,郑剑虹.(2011).非面孔物体倒置效应形成过程的认知神经机制.心理科学进展,19
,1104−1114.]Van Belle,G.,De Graef,P.,Verfaillie,K.,Busigny,T.,&Rossion,B.(2010).Whole not hole:Expert face recognition requires holistic perception.Neuropsychologia, 48
,2620−2629.Wilmer,J.B.,Germine,L.,Chabris,C.F.,Chatterjee,G.,Williams,M.,Loken,E.,et al.(2010).Human face recognition ability is specific and highly heritable.Proceedings of the National Academy of Sciences of the United States of America,107
,5238−5241.Young,A.W.,&Ellis,H.D.(1989).Childhood prosopagnosia.Brain and Cognition,9
,16−47.猜你喜欢
航空学报(2022年7期)2022-09-05
汽车零部件(2022年7期)2022-08-03
心理学报(2022年3期)2022-03-08
心理学报(2022年2期)2022-02-15
心理学报(2021年1期)2021-01-29
汽车与驾驶维修(维修版)(2020年2期)2020-01-07
求知导刊(2017年6期)2017-04-15
股市动态分析(2016年24期)2017-01-07
股市动态分析(2016年23期)2016-12-27
股市动态分析(2016年18期)2016-10-11
